В коллоидной химии одним из свойств липидного бислоя является относительная подвижность (текучесть) отдельных липидных молекул и то, как эта подвижность изменяется с температурой. Эта реакция известна как фазовое поведение бислоя. В широком смысле, при данной температуре липидный бислой может существовать как в жидкой, так и в твердой фазе. Твердую фазу обычно называют « гелевой » фазой. Все липиды имеют характерную температуру, при которой они претерпевают переход ( плавление ) из гелевой в жидкую фазу. В обеих фазах липидные молекулы ограничены двумерной плоскостью мембраны, но в бислоях жидкой фазы молекулы свободно диффундируют в пределах этой плоскости. Таким образом, в жидком бислое данный липид будет быстро обмениваться местоположениями со своим соседом миллионы раз в секунду и будет, посредством процесса случайного блуждания , мигрировать на большие расстояния. [1]
В отличие от этой большой подвижности в плоскости, молекулам липидов очень трудно переходить с одной стороны липидного бислоя на другую. В бислое на основе фосфатидилхолина этот процесс обычно происходит в течение нескольких недель. [2] Это несоответствие можно понять с точки зрения базовой структуры бислоя. Чтобы липид перешел с одного листа на другой, его гидратированная головная группа должна пересечь гидрофобное ядро бислоя, что является энергетически невыгодным процессом. В отличие от бислоев жидкой фазы, липиды в бислое гелевой фазы зафиксированы на месте и не проявляют ни переворота, ни боковой подвижности. Из-за этой ограниченной подвижности гелевые бислои лишены важного свойства жидких бислоев: способности повторно запечатывать небольшие отверстия. Бислои жидкой фазы могут спонтанно залечивать небольшие пустоты, во многом так же, как пленка масла на воде может затекать, чтобы заполнить зазор. Эта функциональность является одной из причин того, что клеточные мембраны обычно состоят из бислоев жидкой фазы. Ограничения движения липидов в липидных бислоях также накладываются присутствием белков в биологических мембранах, особенно в кольцевой липидной оболочке, «прикрепленной» к поверхности интегральных мембранных белков .
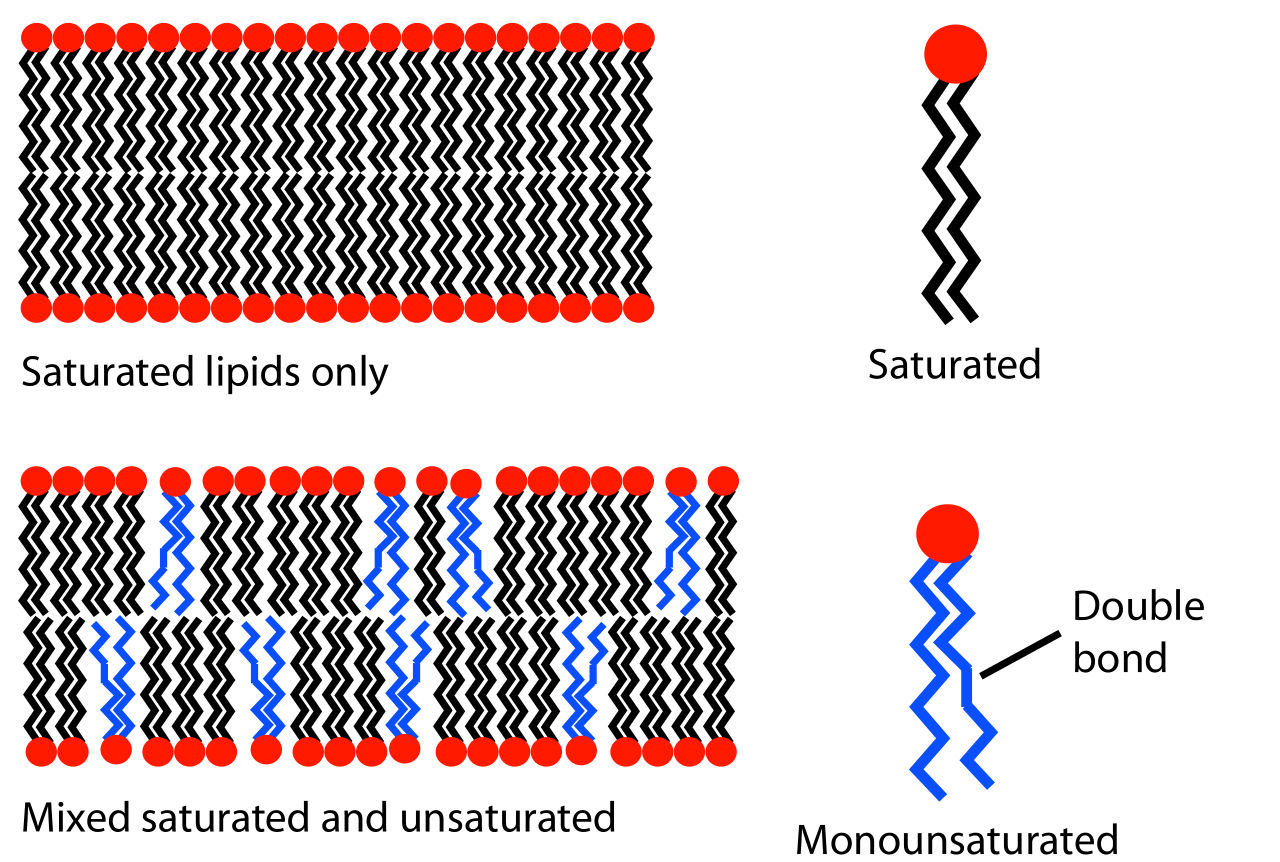
Фазовое поведение липидных бислоев в значительной степени определяется силой притягивающих ван-дер-ваальсовых взаимодействий между соседними липидными молекулами. Степень этого взаимодействия, в свою очередь, регулируется длиной липидных хвостов и тем, насколько хорошо они могут упаковываться вместе. Липиды с более длинными хвостами имеют большую площадь для взаимодействия, что увеличивает силу этого взаимодействия и, следовательно, снижает подвижность липидов. Таким образом, при заданной температуре липид с коротким хвостом будет более текучим, чем идентичный липид с длинным хвостом. [3] Другой способ выразить это — сказать, что температура фазового перехода гель-жидкость увеличивается с увеличением числа атомов углерода в цепях липидных алканов . Насыщенные фосфатидилхолиновые липиды с хвостами длиннее 14 атомов углерода являются твердыми при комнатной температуре, в то время как те, у которых их меньше 14, являются жидкими. Это явление аналогично тому, что парафиновый воск , который состоит из длинных алканов, является твердым при комнатной температуре, в то время как октан ( бензин ), короткий алкан, является жидким.
Помимо длины цепи, на температуру перехода может также влиять степень ненасыщенности липидных хвостов. Ненасыщенная двойная связь может вызывать перегиб в алкановой цепи, нарушая регулярную периодическую структуру. Это нарушение создает дополнительное свободное пространство внутри бислоя, что обеспечивает дополнительную гибкость в соседних цепях. Именно это нарушение упаковки приводит к более низким температурам перехода с увеличением двойных связей. [3] Это особенно мощный эффект; уменьшение общей длины цепи на один углерод обычно изменяет температуру перехода липида на десять градусов Цельсия или меньше, но добавление одной двойной связи может снизить температуру перехода на пятьдесят градусов или больше (см. таблицу) . Пример этого эффекта можно заметить в повседневной жизни, поскольку сливочное масло , которое имеет большой процент насыщенных жиров , является твердым при комнатной температуре, в то время как растительное масло , которое в основном ненасыщенное, является жидким.
Бислои не обязательно должны состоять из одного типа липидов, и, по сути, большинство природных мембран представляют собой сложную смесь различных липидных молекул. Такие смеси часто проявляют свойства, промежуточные по отношению к своим компонентам, но также способны к явлению, не наблюдаемому в однокомпонентных системах: разделению фаз . Если некоторые из компонентов являются жидкими при заданной температуре, а другие находятся в фазе геля, две фазы могут сосуществовать в пространственно разделенных популяциях. Это разделение фаз играет решающую роль в биохимических явлениях, поскольку компоненты мембраны, такие как белки, могут разделяться на одну или другую фазу [5] и, таким образом, локально концентрироваться или активироваться.
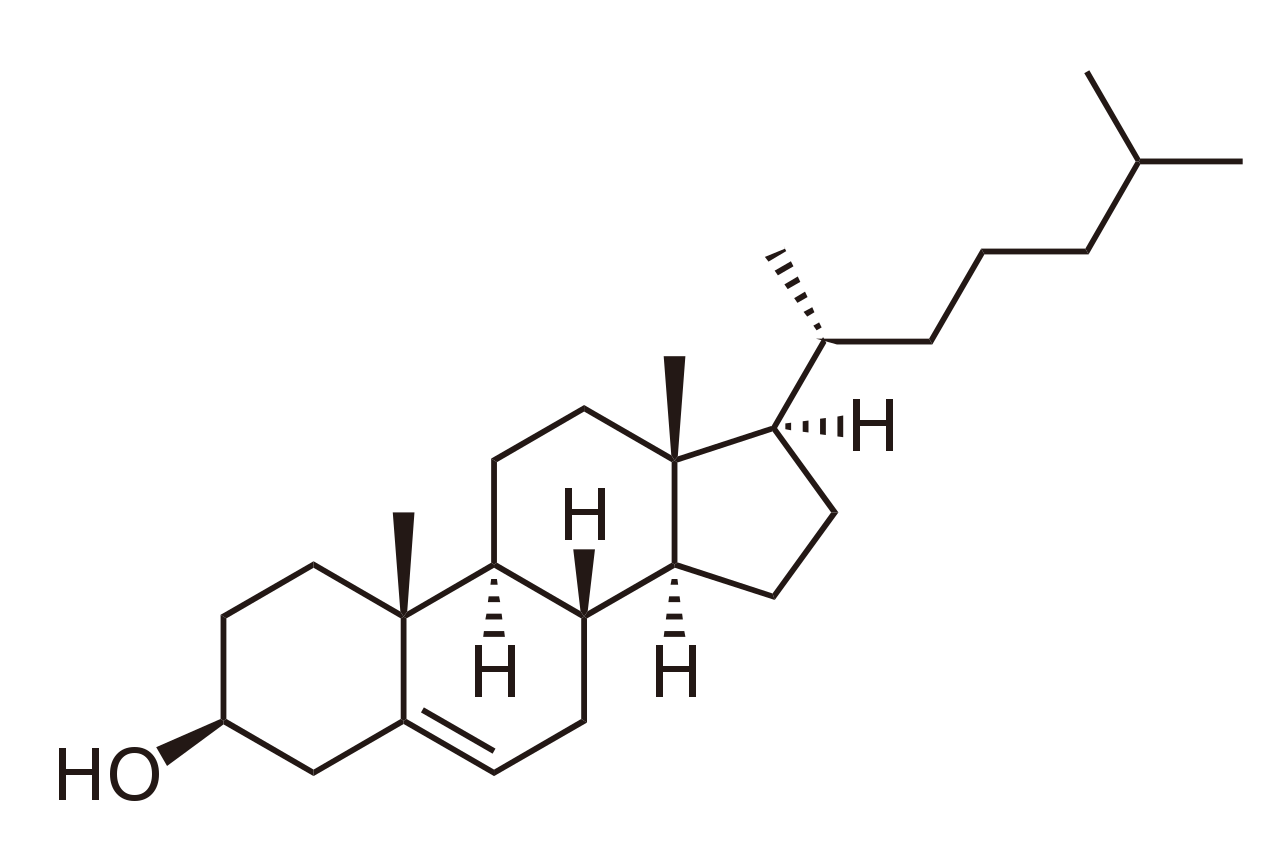
Присутствие холестерина оказывает глубокое, но сложное влияние на свойства липидного бислоя из-за его уникальных физических характеристик. Хотя это липид, холестерин мало похож на фосфолипид . Гидрофильный домен холестерина довольно мал и состоит из одной гидроксильной группы. Рядом с этой гидроксильной группой находится жесткая плоская структура, состоящая из нескольких слитых колец. На противоположном конце кольцевой структуры находится короткий одноцепочечный хвост. На протяжении десятилетий известно, что добавление холестерина к бислою жидкой фазы снижает его проницаемость для воды. [6] [7] Недавно было показано, что режим этого взаимодействия обусловлен интеркалированием холестерина между липидными молекулами, заполнением свободного пространства и уменьшением гибкости окружающих липидных цепей. [8] Это взаимодействие также увеличивает механическую жесткость липидных бислоев жидкой мембраны [9] и снижает их коэффициент боковой диффузии. [10] Напротив, добавление холестерина к бислоям гелевой фазы нарушает локальный порядок упаковки, увеличивая коэффициент диффузии [10] и уменьшая модуль упругости. Взаимодействие холестерина с многокомпонентными системами еще более сложно, поскольку они могут приводить к запутанным фазовым диаграммам . [11] Одной из систем липид-холестерин, которая недавно была тщательно изучена, является липидный плот. Липидные плоты представляют собой обогащенные холестерином гелевые домены, которые потенциально вовлечены в определенные клеточные сигнальные процессы, [12] но этот вопрос остается спорным, и некоторые исследователи сомневаются даже в их существовании in vivo. [13]
Смешанные липидные липосомы могут претерпевать изменения в различные фазовые дисперсионные структуры, называемые липидными полиморфизмами , например, сферические мицеллы , липидные двухслойные ламели и гексагональные фазовые цилиндры, в зависимости от физических и химических изменений в их микроокружении. [14] Температуру фазового перехода липосом и биологических мембран можно измерить с помощью калориметрии , магнитно-резонансной спектроскопии и других методов. [15]