В зрительной физиологии адаптация — это способность сетчатки глаза приспосабливаться к различным уровням освещенности . Естественное ночное зрение , или скотопическое зрение , — это способность видеть в условиях низкой освещенности. У людей за ночное зрение отвечают исключительно палочки , а колбочки способны функционировать только при более высоких уровнях освещенности. [1] Качество ночного видения хуже, чем дневное, поскольку оно ограничено в разрешении и невозможно различить цвета; видны только оттенки серого. [1] Чтобы люди могли перейти от дневного к ночному зрению, они должны пройти период адаптации к темноте продолжительностью до двух часов [2] , в течение которого каждый глаз приспосабливается от высокого к низкому «настройке» свечения, значительно увеличивая чувствительность, за счет на многие порядки. [1] Этот период адаптации различен для палочек и колбочек и является результатом регенерации фотопигментов для повышения чувствительности сетчатки. [1] Световая адаптация, напротив, срабатывает очень быстро, в течение нескольких секунд.
Человеческий глаз может функционировать как при очень темном, так и при очень ярком освещении; его сенсорные возможности достигают девяти порядков величины . Это означает, что самый яркий и самый темный световой сигнал, который может воспринимать глаз, различаются примерно в 1 000 000 000 раз. Однако в любой момент времени глаз может ощутить только коэффициент контрастности , равный 1000. Что обеспечивает более широкий охват, так это то, что глаз адаптирует свое определение того, что является черным.
Глазу требуется примерно 20–30 минут, чтобы полностью адаптироваться от яркого солнечного света к полной темноте, и он становится в 10 000–1 000 000 раз более чувствительным, чем при полном дневном свете. При этом процессе также меняется восприятие цвета глазом (это называется эффектом Пуркинье ). Однако глазу требуется около пяти минут, чтобы адаптироваться от темноты к яркому солнечному свету. Это связано с тем, что колбочки приобретают большую чувствительность при первом входе в темноту в течение первых пяти минут, а палочки берут верх через пять или более минут. [3] Колбочки способны восстановить максимальную чувствительность сетчатки за 9–10 минут в темноте, тогда как палочкам для этого требуется 30–45 минут. [4]
Адаптация к темноте у молодых людей происходит гораздо быстрее и глубже, чем у пожилых людей. [5]
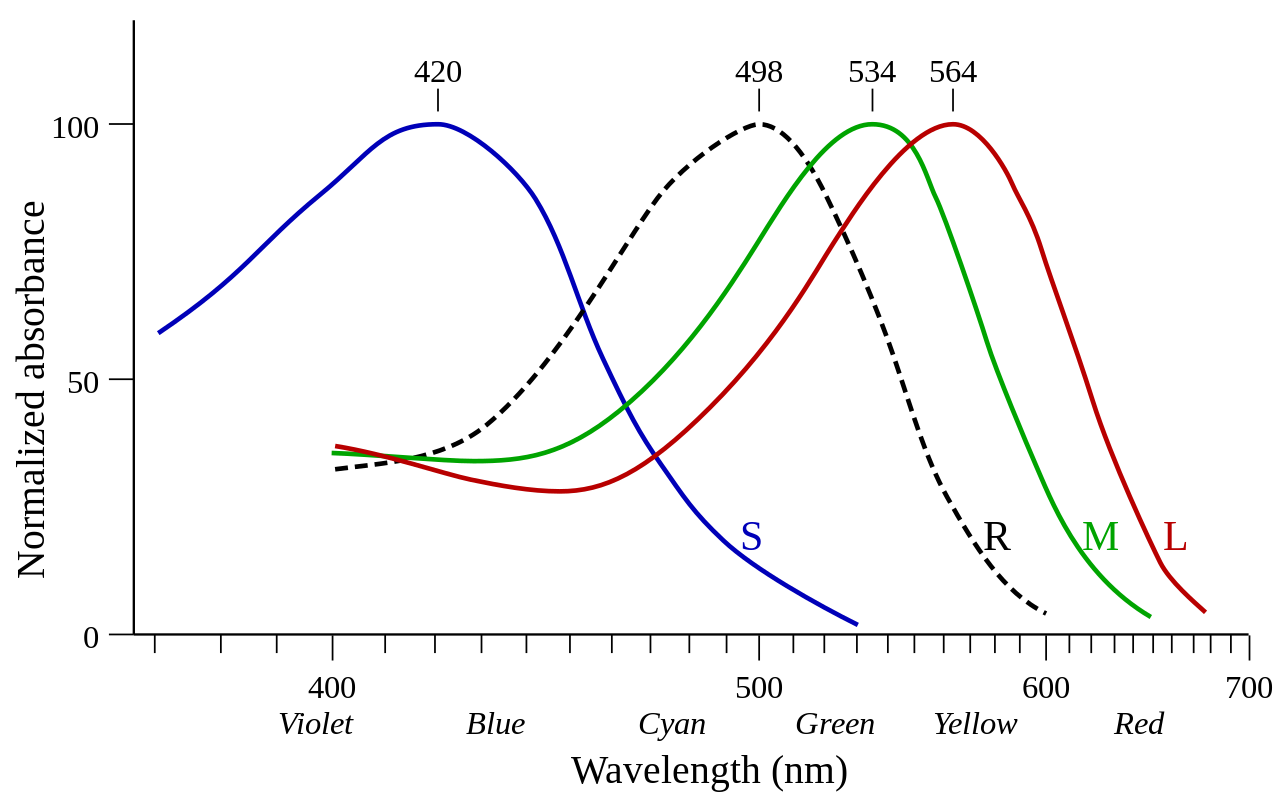
Человеческий глаз содержит три типа фоторецепторов: палочки, колбочки и внутренне светочувствительные ганглиозные клетки сетчатки (ipRGC). Палочки и колбочки отвечают за зрение и связаны со зрительной корой. ipRGC больше связаны с функциями биологических часов и другими частями мозга, но не со зрительной корой. Палочки и колбочки легко отличить по их строению. Фоторецепторы колбочек имеют коническую форму и содержат опсины колбочек в качестве зрительных пигментов. Существует три типа колбочек-фоторецепторов, каждый из которых максимально чувствителен к определенной длине волны света в зависимости от структуры фотопигмента опсина. [6] Различные колбочки максимально чувствительны к коротким длинам волн (синий свет), средним длинам волн (зеленый свет) или длинным волнам (красный свет). Палочки фоторецепторы содержат только один тип фотопигмента, родопсин, который имеет максимальную чувствительность на длине волны примерно 500 нанометров, что соответствует сине-зеленому свету. [6] Распределение фоторецепторных клеток по поверхности сетчатки имеет важные последствия для зрения. [7] Конусные фоторецепторы сосредоточены в углублении в центре сетчатки, известном как центральная ямка , и их количество уменьшается по направлению к периферии сетчатки. [7] И наоборот, палочковидные фоторецепторы присутствуют с высокой плотностью на большей части сетчатки с резким снижением в ямке. В условиях высокой люминесценции в восприятии преобладают колбочки, несмотря на то, что палочек их значительно меньше (примерно от 4,5 до 91 миллиона). [7]


Второстепенным механизмом адаптации является зрачковый световой рефлекс , который очень быстро регулирует количество света, достигающего сетчатки, примерно в десять раз. Поскольку он вносит лишь незначительную долю в общую адаптацию к свету, здесь он далее не рассматривается.
В ответ на изменение уровня окружающего освещения палочки и колбочки глаз функционируют как изолированно, так и совместно, регулируя зрительную систему. Изменения чувствительности палочек и колбочек глаза вносят основной вклад в темновую адаптацию.
Выше определенного уровня яркости (около 0,03 кд/м 2 ) в обеспечении зрения участвует механизм колбочек; фотопическое зрение . Ниже этого уровня вступает в действие стержневой механизм, обеспечивающий скотопическое (ночное) зрение . Диапазон, в котором два механизма работают вместе, называется мезопическим диапазоном , поскольку между двумя механизмами нет резкого перехода. Эта адаптация составляет основу теории двойственности . [8]

Многие животные, например кошки , обладают ночным зрением высокого разрешения, что позволяет им различать объекты с высокими частотами в условиях низкой освещенности. Tapetum lucidum — это отражающая структура, которая отвечает за превосходное ночное зрение, поскольку она отражает свет обратно через сетчатку, подвергая фоторецепторные клетки повышенному количеству света. [9] Большинство животных, обладающих тапетумом lucidum, скорее всего, ведут ночной образ жизни, потому что при отражении света обратно через сетчатку первоначальные изображения становятся размытыми. [9] Люди, как и их родственники-приматы, не обладают тапетумом lucidum и поэтому были предрасположены к дневному образу жизни. [10]
Несмотря на то, что разрешение человеческого дневного зрения намного превосходит разрешение ночного видения, ночное зрение человека дает много преимуществ. Как и многие хищные животные, люди могут использовать свое ночное зрение, чтобы охотиться на других животных и устраивать засады без их ведома. Более того, в случае чрезвычайной ситуации, возникающей ночью, люди могут увеличить свои шансы на выживание, если они смогут воспринимать окружающую обстановку и добраться до безопасного места. Оба этих преимущества можно использовать, чтобы объяснить, почему люди не полностью утратили способность видеть в темноте от своих ночных предков. [11]

Родопсин , биологический пигмент фоторецепторов сетчатки, немедленно фотообесцвечивается в ответ на свет. [12] Визуальная фототрансдукция начинается с изомеризации пигментного хромофора из 11-цис в полностью транс- ретиналь . [13] Затем этот пигмент диссоциирует на свободный опсин и полностью транс-ретиналь. Адаптация к темноте как палочек, так и колбочек требует регенерации зрительного пигмента из опсина и 11-цис-ретиналя. [13] Таким образом, время, необходимое для темновой адаптации и регенерации пигмента, во многом определяется локальной концентрацией 11-цис-ретиналя и скоростью, с которой он доставляется к опсину в отбеленных палочках. [14] Уменьшение притока ионов кальция после закрытия канала вызывает фосфорилирование метародопсина II и ускоряет инактивацию цис-ретиналя в транс-ретиналь. [13] Фосфорилирование активированного родопсина опосредуется рековином . [13] Регенерация фотопигментов происходит во время темновой адаптации, хотя и с заметно разной скоростью. [15] Палочки более чувствительны к свету, поэтому им требуется больше времени, чтобы полностью адаптироваться к изменению освещения. Палочки, фотопигменты которых регенерируют медленнее, не достигают максимальной чувствительности около двух часов. [3] [16] Колбочкам требуется примерно 9–10 минут, чтобы адаптироваться к темноте. [3] Чувствительность к свету модулируется изменениями внутриклеточных ионов кальция и циклического гуанозинмонофосфата . [17]
Чувствительность палочечного пути значительно улучшается в течение 5–10 минут в темноте. Цветовое тестирование использовалось для определения момента, когда стержневой механизм вступит в силу; когда стержневой механизм берет на себя управление, цветные пятна кажутся бесцветными, поскольку только колбочки кодируют цвет. [18]
На то, как быстро стержневой механизм становится доминирующим, влияют три фактора:
В условиях скотопии в фоторецепторах высока внутриклеточная концентрация цГМФ . цГМФ связывается с цГМФ-закрытыми Na + -каналами и открывает их , обеспечивая приток натрия и кальция. [22] Приток натрия способствует деполяризации, в то время как приток кальция увеличивает локальные концентрации кальция вблизи рецептора. Кальций связывается с модулирующим белком, которым, как предполагается, является GUCA1B , [23] устраняя стимулирующий эффект этого белка на гуанилилциклазу . [22] Это снижает выработку цГМФ гуанилатциклазой и снижает концентрацию цГМФ во время продолжительной темноты. Повышенная концентрация кальция также увеличивает активность фосфодиэстеразы [22] , которая гидролизует цГМФ и еще больше снижает его концентрацию. Это уменьшает открытие цГМФ-закрытых Na + -каналов, вызывающих гиперполяризацию клетки, что снова делает ее чувствительной к небольшому увеличению яркости. Без темновой адаптации фоторецептор оставался бы деполяризованным в скотопических условиях и, следовательно, не реагировал бы на небольшие изменения яркости.
Торможение нейронов также влияет на активацию в синапсах. Вместе с обесцвечиванием палочковидного или колбочкового пигмента тормозится слияние сигналов на ганглиозных клетках , уменьшая конвергенцию.
Альфа-адаптация, то есть быстрые колебания чувствительности, обеспечивается нервным контролем. Слияние сигналов за счет диффузных ганглиозных клеток, а также горизонтальных и амакриновых клеток обеспечивает кумулятивный эффект. Таким образом, эта область стимуляции обратно пропорциональна интенсивности света: сильный стимул в 100 палочек эквивалентен слабому стимулу в 1000 палочек.
При достаточно ярком освещении конвергенция низкая, но при адаптации к темноте конвергенция сигналов палочек усиливается. Это связано не со структурными изменениями, а с возможным отключением торможения, останавливающего схождение сообщений при ярком свете. Если открыт только один глаз, закрытый глаз при повторном открытии должен адаптироваться отдельно, чтобы соответствовать уже адаптированному глазу. [3]
Офтальмологи иногда измеряют темновую адаптацию пациентов с помощью прибора, известного как темновой адаптометр.
Существует несколько коммерческих темновых адаптометров (AdaptDx, MetroVision MonCvONE, Roland Consult DARK-Adaptometer). Помимо этих устройств со свободным обзором, недавно был внедрен рабочий процесс с отслеживанием глазного дна путем модификации устройства микропериметрии , которое позволяет тестировать пациентов с нестабильной фиксацией. [24]
Многочисленные клинические исследования показали, что функция адаптации к темноте резко ухудшается на самых ранних стадиях возрастной макулярной дегенерации (ВМД), пигментного ретинита (РП) и других заболеваний сетчатки, причем ухудшение ухудшается по мере прогрессирования заболевания. [25] [26] ВМД — это хроническое прогрессирующее заболевание, при котором часть сетчатки, называемая макулой, со временем медленно разрушается. Это основная причина потери зрения среди людей в возрасте 50 лет и старше. [27] Он характеризуется разрушением комплекса РПЭ/мембраны Бруха в сетчатке, что приводит к накоплению отложений холестерина в макуле. Со временем эти отложения становятся клинически видимыми друзами, которые влияют на здоровье фоторецепторов, вызывая воспаление и предрасположенность к хориоидальной неоваскуляризации (ХНВ). Во время течения ВМД функция РПЭ/Бруха продолжает ухудшаться, затрудняя транспорт питательных веществ и кислорода к фоторецепторам палочек и колбочек. В качестве побочного эффекта этого процесса у фоторецепторов наблюдается нарушение темновой адаптации, поскольку им требуются эти питательные вещества для пополнения фотопигментов и выведения опсина, чтобы восстановить скотопическую чувствительность после воздействия света.
Измерение функции адаптации пациента к темноте — это, по сути, биоанализ здоровья мембраны Бруха. Таким образом, исследования показали, что, измеряя темновую адаптацию, врачи могут обнаружить субклиническую ВМД как минимум на три года раньше, чем она становится клинически очевидной. [28]
.jpg/1280px-%D0%9F%D0%B5%D1%80%D1%81%D0%B5%D1%97%D0%B4%D0%B8-2021_-_%D0%B5%D0%BA%D1%81%D0%BA%D1%83%D1%80%D1%81%D1%96%D1%8F_%D0%93%D0%BE%D0%BB%D0%BE%D0%B2%D0%BD%D0%BE%D1%97_%D0%B0%D1%81%D1%82%D1%80%D0%BE%D0%BD%D0%BE%D0%BC%D1%96%D1%87%D0%BD%D0%BE%D1%97_%D0%BE%D0%B1%D1%81%D0%B5%D1%80%D0%B2%D0%B0%D1%82%D0%BE%D1%80%D1%96%D1%97_%D0%9D%D0%90%D0%9D_%D0%A3%D0%BA%D1%80%D0%B0%D1%97%D0%BD%D0%B8_(5).jpg)
Было предложено или продемонстрировано несколько различных методов с разным уровнем доказательности для увеличения скорости адаптации зрения в темноте.
В результате того, что палочки имеют максимальную чувствительность на длине волны 500 нанометров, они не могут воспринимать все цвета зрительного спектра. Поскольку палочки нечувствительны к длинным волнам, использование красного света и очков с красными линзами стало обычной практикой для ускорения адаптации к темноте. [29] Для того, чтобы адаптация к темноте значительно ускорилась, в идеале человеку следует начать эту практику за 30 минут до перехода в режим низкой люминесценции. [30] Эта практика позволит человеку сохранить фотопическое (дневное) зрение, одновременно готовясь к скотопическому зрению. Нечувствительность к красному свету предотвратит дальнейшее обесцвечивание палочек и позволит фотопигменту родопсину восстановиться до его активной конформации. [29] Как только человек попадает в темноту, большинство его палочек уже адаптируются к темноте и смогут передавать зрительные сигналы в мозг без периода аккомодации. [30]
Концепция красных линз для адаптации к темноте основана на экспериментах Антуана Беклера и его ранних работах в области радиологии. В 1916 году ученый Вильгельм Тренделенбург изобрел первую пару красных адаптационных очков для рентгенологов, чтобы приспособить их глаза к просмотру экранов во время рентгеноскопических процедур.
Хотя многие аспекты зрительной системы человека остаются неясными, теория эволюции палочек и колбочек фотопигментов согласована большинством ученых. Считается, что самыми ранними зрительными пигментами были пигменты колбочек, а белки палочек-опсинов появились позже. [31] После эволюции млекопитающих от их предков-рептилий примерно 275 миллионов лет назад наступила ночная фаза, во время которой сложное цветовое зрение было утрачено. [31] Поскольку эти промлекопитающие вели ночной образ жизни, они увеличили свою чувствительность в условиях низкой люминесценции и уменьшили свою фотопическую систему с тетрахроматической до дихроматической. [31] Переход к ночному образу жизни потребует большего количества палочек-фоторецепторов для поглощения синего света, излучаемого Луной в ночное время. [32] Можно экстраполировать, что высокое соотношение палочек и колбочек, присутствующее в глазах современного человека, сохранилось даже после перехода от ночного режима к дневному.
Считается, что появление трихроматии у приматов произошло примерно 55 миллионов лет назад, когда температура поверхности планеты начала повышаться. [31] Приматы вели дневной, а не ночной образ жизни, и поэтому им требовалась более точная фотопическая зрительная система. Третий фотопигмент конуса был необходим для покрытия всего зрительного спектра, позволяя приматам лучше различать фрукты и обнаруживать те, которые имеют наибольшую пищевую ценность. [31]

Витамин А необходим для правильного функционирования человеческого глаза. Фотопигмент родопсин, обнаруженный в палочковых клетках человека, состоит из ретиналя, формы витамина А, связанного с белком опсина. [35] При поглощении света родопсин разлагался на ретиналь и опсин посредством обесцвечивания. [35] Тогда у ретиналя могла быть одна из двух судеб: он мог рекомбинировать с опсином, образуя родопсин, или превращаться в свободный ретинол. [35] Американский учёный Джордж Уолд был первым, кто осознал, что зрительная система расходует витамин А и его восполнение зависит от диеты. [35]
Витамин А выполняет множество функций в организме человека, помимо здорового зрения. Это жизненно важно для поддержания здоровой иммунной системы, а также для содействия нормальному росту и развитию. [36] Среднестатистический взрослый мужчина и женщина должны потреблять 900 и 700 микрограммов витамина А в день соответственно. [36] Потребление более 3000 микрограммов в день может привести к острому или хроническому гипервитаминозу А. [37]
Витамин А присутствует как в животных, так и в растительных источниках в виде ретиноидов и каротиноидов соответственно. [36] Ретиноиды могут быть использованы организмом сразу после всасывания в сердечно-сосудистую систему; однако каротиноиды растительного происхождения должны быть преобразованы в ретинол перед использованием организмом. [36] Самыми высокими источниками витамина А животного происхождения являются печень, молочные продукты и рыба. [36] Фрукты и овощи, содержащие большое количество каротиноидов, имеют темно-зеленый, желтый, оранжевый и красный цвет. [36]
Белки опсины на основе витамина А использовались для восприятия света в организмах на протяжении большей части эволюционной истории, начиная примерно с 3 миллиардов лет назад. [38] Эта особенность была передана от одноклеточных к многоклеточным организмам, включая Homo sapiens. [38] Этот витамин, скорее всего, был выбран в ходе эволюции для восприятия света, поскольку ретиналь вызывает сдвиг поглощения фоторецепторов в диапазон видимого света. [38] Этот сдвиг в поглощении особенно важен для жизни на Земле, поскольку он обычно соответствует пиковой интенсивности солнечного излучения на ее поверхности. [38] Вторая причина, по которой сетчатка стала жизненно важной для человеческого зрения, заключается в том, что она претерпевает большие конформационные изменения под воздействием света. [38] Считается, что это конформационное изменение облегчает белку фоторецептора различать его молчащее и активированное состояние, тем самым лучше контролируя зрительную фототрансдукцию. [38]
Были проведены различные исследования, проверяющие эффективность добавок витамина А на адаптацию к темноте. В исследовании Cideciyan et al. продолжительность темновой адаптации измерялась у пациента с системным дефицитом витамина А (VAD) до и после приема витамина А. [39] Функция адаптации к темноте измерялась до приема добавок, через 1 день после лечения и через 75 дней после лечения. Было замечено, что всего лишь через один день приема витамина А кинетика восстановления темновой адаптации значительно ускорилась после обесцвечивания фоторецепторов. [39] Темновая адаптация еще больше ускорилась после 75 дней лечения. [39]
Последующее исследование Kemp et al. изучали адаптацию к темноте у пациентов с первичным билиарным циррозом печени и болезнью Крона , у которых был дефицит витамина А. [40] В течение 8 дней перорального приема витамина А у обоих пациентов зрительные функции восстановились до нормальных значений. [40] Кроме того, после приема добавок кинетика адаптации значительно улучшилась у обоих субъектов. [40]
Более поздние независимые исследования дистрофии глазного дна Сорби подтвердили влияние витамина А на адаптацию к темноте. [41] Аналогичным образом было показано, что витамин А ускоряет (в меньшей степени) адаптацию к темноте при ВМД. [42] [43]
Антоцианы составляют большую часть из 4000 известных фитохимических флавоноидов. [44] Эта группа, состоящая примерно из 600 биологически активных антиоксидантов, оказывает самое сильное физиологическое воздействие среди всех растительных соединений. [45] Эти химические вещества также являются наиболее заметными из флавоноидных фитохимических веществ, поскольку они придают многим видам растений ярко-синюю, красную или фиолетовую пигментацию. [45] Антоцианы также служат для защиты фотосинтетических тканей от прямых солнечных лучей. [46] Кроме того, антиоксидантные , противовоспалительные и вазопротективные свойства антоцианов позволяют им оказывать разнообразное воздействие на здоровье. [45] У людей антоцианы эффективны при различных заболеваниях, включая неврологические нарушения, атеросклероз, диабет, а также нарушения зрения. [46] Антоцианы часто взаимодействуют с другими фитохимическими веществами, усиливая биологические эффекты; поэтому вклад отдельных биомолекул по-прежнему трудно расшифровать. [44] Благодаря тому, что антоцианы придают цветам яркую окраску, растения, содержащие эти фитохимические вещества, естественным образом успешно привлекают опылителей, таких как птицы и пчелы. [46] Фрукты и овощи, производимые такими растениями, также ярко пигментированы, что привлекает животных, чтобы съесть их и распространить семена. [46] Благодаря этому естественному механизму антоциансодержащие растения широко распространены в большинстве регионов мира. Высокое обилие и распространение растений, содержащих антоцианы, делают его естественным источником пищи для многих животных. Благодаря ископаемым свидетельствам известно, что эти соединения в больших количествах употреблялись в пищу примитивными гомининами. [45]
Известно, что во время Первой и Второй мировых войн авиаторы британских ВВС употребляли большое количество черничного варенья. Авиаторы употребляли эту богатую антоцианами пищу из-за ее многочисленных визуальных преимуществ, включая ускоренную адаптацию к темноте, что было бы полезно для ночных бомбардировок. [47]

Ярко окрашенные фрукты и овощи богаты антоцианами. Интуитивно это имеет смысл, поскольку антоцианы обеспечивают пигментацию растений. Ежевика — самый богатый антоцианами продукт, содержащий 89–211 миллиграммов на 100 граммов. [46] Другие продукты, богатые этим фитохимическим веществом, включают красный лук, чернику, чернику, красную капусту и баклажаны. [46] Употребление в пищу любого из этих источников пищи приводит к образованию множества фитохимических веществ в дополнение к антоцианам, поскольку они естественным образом существуют вместе. [44] Среднестатистическому взрослому человеку ежедневное потребление антоцианов составляет примерно 200 миллиграммов; однако это значение может достигать нескольких граммов в день, если человек потребляет добавки с флавоноидами. [44]
Антоцианы ускоряют адаптацию человека к темноте, усиливая регенерацию палочкового фотопигмента, родопсина. [48] Антоцианы достигают этого, связываясь непосредственно с опсином при разложении родопсина на его отдельные составляющие под действием света. [48] После связывания с опсином антоциан меняет свою структуру, тем самым ускоряя доступ к связывающему карману сетчатки. Соблюдая диету, богатую антоцианами, человек может вырабатывать родопсин за более короткие периоды времени из-за повышенного сродства опсина к сетчатке. [48] Благодаря этому механизму человек может ускорить адаптацию к темноте и достичь ночного видения за более короткий период времени.
В двойном слепом плацебо-контролируемом исследовании, проведенном Nakaishi et al. Ряду участников был предоставлен порошкообразный концентрат антоцианов, полученный из черной смородины. [49] [ ненадежный медицинский источник? ] Участники получили одну из трех доз антоцианов, чтобы определить, возник ли результат в зависимости от дозы. Период темновой адаптации измерялся до и через два часа после приема добавок у всех участников. Результаты этого эксперимента показывают, что антоцианы значительно ускоряют темновую адаптацию всего лишь при одном уровне дозы по сравнению с плацебо. [49] [ ненадежный медицинский источник? ] Наблюдение за данными в целом Nakaishi et al. пришли к выводу, что антоцианы эффективно сокращают период адаптации к темноте дозозависимым образом. [49] [ ненадежный медицинский источник? ]
Несмотря на то, что многие ученые считают, что антоцианы способствуют ускорению адаптации человека к темноте, исследование, проведенное Kalt et al. в 2014 году было показано, что антоцианы черники не оказывают никакого эффекта. В этом исследовании были проведены два двойных слепых плацебо-контролируемых исследования для изучения адаптации к темноте после употребления продуктов из черники. [50] Ни в одном исследовании потребление антоцианов черники не повлияло на продолжительность темновой адаптации. [50] На основании этих результатов Kalt et al. пришли к выводу, что антоцианы черники не оказывают существенного влияния на темновой компонент адаптации человеческого зрения. [50]
При адаптации к свету глаз должен быстро адаптироваться к фоновой освещенности, чтобы иметь возможность различать объекты на этом фоне. Процесс светоадаптации происходит в течение пяти минут.
Фотохимическая реакция – это:

Используя эксперименты с порогом приращения, адаптацию к свету можно измерить клинически. [51] В эксперименте с увеличением порога тестовый стимул предъявляется на фоне определенной яркости , стимул увеличивается до тех пор, пока не будет достигнут порог обнаружения на фоне. С помощью этого метода получается монофазная или двухфазная кривая зависимости порога от интенсивности TVI как для колбочек, так и для палочек .
Если взять пороговую кривую для одной системы (т. е. только конусов или только стержней ) изолированно, то можно увидеть, что она имеет четыре участка: [52]

Недостаточность адаптации чаще всего проявляется как недостаточная адаптация к темноте, называемая куриной слепотой или никталопией . [35] Противоположная проблема, известная как гемералопия , то есть неспособность ясно видеть при ярком свете, встречается гораздо реже.
Ямка слепа к тусклому свету (из-за того, что она состоит только из колбочек), а палочки более чувствительны, поэтому тусклую звезду в безлунную ночь нужно смотреть сбоку , поэтому она стимулирует палочки. Это не связано с шириной зрачка, поскольку искусственный зрачок фиксированной ширины дает те же результаты. [3]
Куриная слепота может быть вызвана рядом факторов, наиболее частым из которых является дефицит витамина А. При обнаружении никталопии на достаточно раннем этапе можно обратить вспять и восстановить зрительную функцию; однако; длительный дефицит витамина А может привести к необратимой потере зрения, если его не лечить. [55]
Куриная слепота особенно распространена в развивающихся странах из-за недостаточного питания и, следовательно, недостатка витамина А в рационе. [55] В развитых странах куриная слепота исторически была редкостью из-за достаточного наличия пищи; однако ожидается, что заболеваемость будет увеличиваться по мере того, как ожирение становится более распространенным. Рост показателей ожирения соответствует увеличению количества бариатрических операций, вызывающих нарушение всасывания витамина А в организме человека. [55]