Палочковые клетки — это фоторецепторные клетки в сетчатке глаза, которые могут функционировать при слабом освещении лучше, чем другой тип зрительных фоторецепторов, колбочки . Палочки обычно сосредоточены на внешних краях сетчатки и используются в периферическом зрении . В среднем в сетчатке человека насчитывается около 92 миллионов палочковых клеток (по сравнению с ~6 миллионами колбочек). [1] Палочковые клетки более чувствительны, чем колбочки, и почти полностью отвечают за ночное зрение . Однако палочки играют небольшую роль в цветовом зрении , что является основной причиной того, что цвета гораздо менее заметны при тусклом свете.
Палочки немного длиннее и тоньше, чем колбочки, но имеют ту же самую базовую структуру. Диски, содержащие опсин, лежат на конце клетки, прилегающем к пигментному эпителию сетчатки , который, в свою очередь, прикреплен к внутренней части глаза . Структура стекированных дисков детекторной части клетки обеспечивает очень высокую эффективность. Палочки встречаются гораздо чаще, чем колбочки, с примерно 120 миллионами палочковых клеток по сравнению с 6-7 миллионами колбочек. [2]
Как и колбочки, палочковидные клетки имеют синаптическое окончание, внутренний сегмент и внешний сегмент. Синаптическое окончание образует синапс с другим нейроном, обычно биполярной клеткой или горизонтальной клеткой . Внутренний и внешний сегменты соединены ресничкой , [ 3] которая выстилает дистальный сегмент. [4] Внутренний сегмент содержит органеллы и ядро клетки , в то время как внешний сегмент палочки (сокращенно ROS), который направлен к задней части глаза, содержит поглощающие свет материалы. [3]
Человеческая палочка имеет диаметр около 2 микрон и длину 100 микрон. [5] Палочки не все морфологически одинаковы; у мышей палочки, расположенные близко к внешнему плексиформному синаптическому слою, демонстрируют уменьшенную длину из-за укороченного синаптического окончания. [6]

У позвоночных активация фоторецепторной клетки представляет собой гиперполяризацию (торможение) клетки. Когда они не стимулируются, например, в темноте, палочки и колбочки деполяризуются и спонтанно высвобождают нейротрансмиттер. Этот нейротрансмиттер гиперполяризует биполярную клетку . Биполярные клетки существуют между фоторецепторами и ганглиозными клетками и передают сигналы от фоторецепторов к ганглиозным клеткам . В результате гиперполяризации биполярной клетки она не высвобождает свой трансмиттер в биполярно-ганглиозном синапсе , и синапс не возбуждается.
Активация фотопигментов светом посылает сигнал путем гиперполяризации палочковидной клетки, что приводит к тому, что палочковидная клетка не посылает свой нейромедиатор, что приводит к тому, что биполярная клетка затем высвобождает свой медиатор в биполярно-ганглиозном синапсе и возбуждает синапс.
Деполяризация палочковых клеток (вызывающая высвобождение их нейротрансмиттера) происходит из-за того, что в темноте клетки имеют относительно высокую концентрацию циклического гуанозин 3'-5' монофосфата (цГМФ), который открывает ионные каналы (в основном натриевые каналы, хотя кальций также может проникать через эти каналы). Положительные заряды ионов, которые попадают в клетку по ее электрохимическому градиенту, изменяют мембранный потенциал клетки , вызывают деполяризацию и приводят к высвобождению нейротрансмиттера глутамата . Глутамат может деполяризовать некоторые нейроны и гиперполяризовать другие, позволяя фоторецепторам взаимодействовать антагонистическим образом.
Когда свет попадает на фоторецепторные пигменты внутри фоторецепторной клетки, пигмент меняет форму. Пигмент, называемый родопсином (конопсин находится в колбочках), состоит из большого белка, называемого опсином (расположенного в плазматической мембране), к которому прикреплена ковалентно связанная простетическая группа: органическая молекула, называемая ретиналем (производное витамина А ). Ретиналь существует в форме 11-цис-ретиналя в темноте, а стимуляция светом заставляет его структуру изменяться на полностью транс-ретиналь. Это структурное изменение вызывает повышенное сродство к регуляторному белку, называемому трансдуцином (тип G-белка). При связывании с родопсином альфа-субъединица G-белка заменяет молекулу GDP на молекулу GTP и активируется. Эта замена заставляет альфа-субъединицу G-белка диссоциировать от бета- и гамма-субъединиц G-белка. В результате альфа-субъединица теперь свободна для связывания с фосфодиэстеразой цГМФ (эффекторным белком). [8] Альфа-субъединица взаимодействует с ингибирующими гамма-субъединицами ФДЭ и не дает им блокировать каталитические сайты на альфа- и бета-субъединицах ФДЭ, что приводит к активации фосфодиэстеразы цГМФ, которая гидролизует цГМФ (вторичный мессенджер), расщепляя его на 5'-ГМФ. [9] Снижение цГМФ позволяет закрыть ионные каналы, предотвращая приток положительных ионов, гиперполяризуя клетку и останавливая высвобождение нейротрансмиттера глутамата. [3] Хотя колбочки в основном используют нейротрансмиттерное вещество ацетилхолин , палочки используют его разновидность. Весь процесс, посредством которого свет инициирует сенсорную реакцию, называется зрительной фототрансдукцией.
Активация одной единицы родопсина , светочувствительного пигмента в палочках, может привести к большой реакции в клетке, поскольку сигнал усиливается. После активации родопсин может активировать сотни молекул трансдуцина, каждая из которых, в свою очередь, активирует молекулу фосфодиэстеразы, которая может расщеплять более тысячи молекул цГМФ в секунду. [3] Таким образом, палочки могут иметь большую реакцию на небольшое количество света.
Поскольку ретинальный компонент родопсина происходит из витамина А , дефицит витамина А вызывает дефицит пигмента, необходимого палочковым клеткам. Следовательно, меньше палочковидных клеток способны адекватно реагировать в темных условиях, а поскольку колбочковидные клетки плохо приспособлены для зрения в темноте, может возникнуть ночная слепота .
Палочки используют три тормозных механизма (механизмы отрицательной обратной связи), чтобы обеспечить быстрый возврат в состояние покоя после вспышки света.
Во-первых, существует родопсинкиназа (РК), которая фосфорилирует цитозольный хвост активированного родопсина на множественных серинах, частично ингибируя активацию трансдуцина . Кроме того, ингибирующий белок, аррестин , затем связывается с фосфорилированными родопсинами, чтобы еще больше ингибировать активность родопсина.
В то время как аррестин отключает родопсин, белок RGS (действующий как белок, активирующий ГТФазу (GAP)) переводит трансдуцин (G-белок) в состояние «выключено», увеличивая скорость гидролиза связанного ГТФ в ГДФ.
Когда концентрация цГМФ падает, ранее открытые чувствительные к цГМФ каналы закрываются, что приводит к снижению притока ионов кальция. Связанное с этим снижение концентрации ионов кальция стимулирует чувствительные к ионам кальция белки, которые затем активируют гуанилатциклазу для пополнения цГМФ, быстро восстанавливая его до исходной концентрации. Это открывает чувствительные к цГМФ каналы и вызывает деполяризацию плазматической мембраны. [10]
Когда палочки подвергаются воздействию высокой концентрации фотонов в течение длительного периода времени, они становятся десенсибилизированными (адаптированными) к окружающей среде.
Поскольку родопсин фосфорилируется родопсинкиназой (членом GPCR-киназ (GRK)), он связывается с высоким сродством с аррестином . Связанный аррестин может способствовать процессу десенсибилизации по крайней мере двумя способами. Во-первых, он предотвращает взаимодействие между G-белком и активированным рецептором. Во-вторых, он служит адаптерным белком, помогая рецептору в клатрин-зависимом эндоцитозном аппарате (для индукции рецептор-опосредованного эндоцитоза). [10]
Клетка-палочка достаточно чувствительна, чтобы реагировать на одиночный фотон света [11] и примерно в 100 раз более чувствительна к одному фотону, чем колбочки. Поскольку палочкам для функционирования требуется меньше света, чем колбочкам, они являются основным источником зрительной информации ночью ( скотопическое зрение ). Клетки-колбочки, с другой стороны, требуют от десятков до сотен фотонов для активации. Кроме того, несколько клеток-палочек сходятся на одном интернейроне , собирая и усиливая сигналы. Однако эта конвергенция обходится ценой остроты зрения (или разрешения изображения ), поскольку объединенная информация от нескольких клеток менее различима, чем если бы зрительная система получала информацию от каждой клетки-палочки по отдельности.
Клетки палочек также реагируют на свет медленнее, чем колбочки, и стимулы, которые они получают, добавляются примерно за 100 миллисекунд. Хотя это делает палочки более чувствительными к меньшему количеству света, это также означает, что их способность ощущать временные изменения, такие как быстро меняющиеся изображения, менее точна, чем у колбочек. [3]
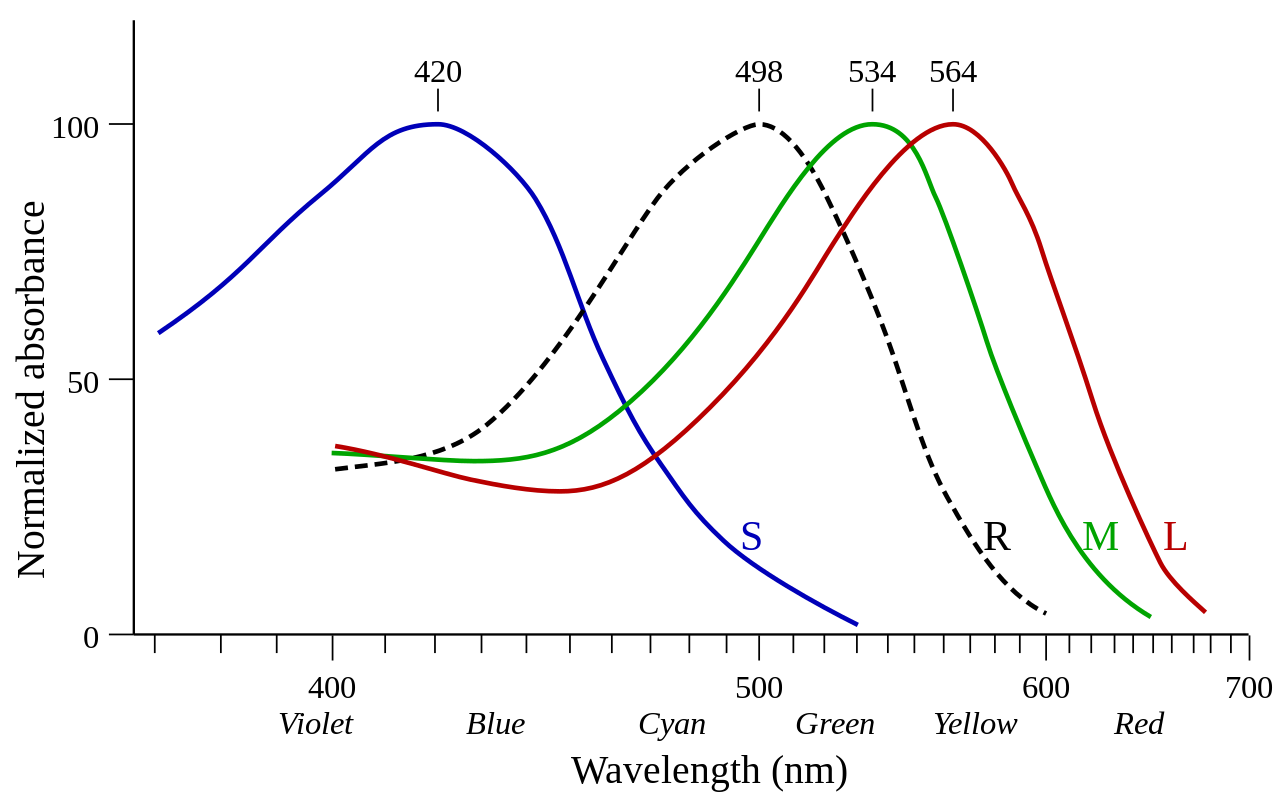
Эксперименты Джорджа Уолда и других показали, что палочки наиболее чувствительны к длинам волн света около 498 нм (зелено-голубой) и нечувствительны к длинам волн более 640 нм (красный). Это отвечает за эффект Пуркинье : по мере того, как интенсивность света уменьшается в сумерках, палочки берут верх, и до того, как цвет полностью исчезнет, пиковая чувствительность зрения смещается в сторону пиковой чувствительности палочек (сине-зеленый). [13]
Список различных типов клеток в организме взрослого человека