Лиганды связываются со специфическими рецепторными белками в зависимости от формы активного центра белка.
Рецептор высвобождает мессенджер после того, как лиганд соединился с рецептором.
В биохимии и фармакологии рецепторы представляют собой химические структуры, состоящие из белка , которые получают и передают сигналы, которые могут быть интегрированы в биологические системы. [1] Эти сигналы обычно являются химическими мессенджерами [nb 1], которые связываются с рецептором и вызывают физиологические реакции, такие как изменение электрической активности клетки . Например, ГАМК , ингибирующий нейротрансмиттер , ингибирует электрическую активность нейронов, связываясь с рецепторами ГАМК А. [2] Существует три основных способа классификации действия рецептора: передача сигнала, усиление или интеграция. [3] Ретрансляция посылает сигнал дальше, усиление увеличивает эффект одного лиганда , а интеграция позволяет включить сигнал в другой биохимический путь. [ 3]
Рецепторные белки можно классифицировать по их местоположению. Рецепторы клеточной поверхности , также известные как трансмембранные рецепторы, включают лиганд-управляемые ионные каналы , рецепторы, сопряженные с G-белком , и рецепторы гормонов , связанные с ферментами . [1] Внутриклеточные рецепторы — это те, которые находятся внутри клетки, и включают цитоплазматические рецепторы и ядерные рецепторы . [1] Молекула, которая связывается с рецептором, называется лигандом и может быть белком, пептидом (коротким белком) или другой небольшой молекулой , такой как нейротрансмиттер , гормон , фармацевтический препарат, токсин, ион кальция или части внешней части вируса или микроба. Эндогенно продуцируемое вещество, которое связывается с определенным рецептором, называется его эндогенным лигандом. Например, эндогенным лигандом для никотинового ацетилхолинового рецептора является ацетилхолин , но он также может быть активирован никотином [4] [5] и заблокирован кураре . [6] Рецепторы определенного типа связаны с определенными клеточными биохимическими путями, которые соответствуют сигналу. Хотя в большинстве клеток обнаружено множество рецепторов, каждый рецептор будет связываться только с лигандами определенной структуры. Это можно сравнить с тем, как замки будут принимать только ключи определенной формы . Когда лиганд связывается с соответствующим рецептором, он активирует или ингибирует связанный с рецептором биохимический путь, который также может быть высокоспециализированным.
Структуры рецепторов весьма разнообразны и включают, среди прочего, следующие основные категории:
Тип 1: Лиганд-зависимые ионные каналы (ионотропные рецепторы) – Эти рецепторы обычно являются мишенями быстрых нейротрансмиттеров, таких как ацетилхолин (никотиновый) и ГАМК ; активация этих рецепторов приводит к изменениям в движении ионов через мембрану. Они имеют гетеромерную структуру, в которой каждая субъединица состоит из внеклеточного лиганд-связывающего домена и трансмембранного домена, который включает четыре трансмембранные альфа-спирали . Лиганд-связывающие полости расположены на границе между субъединицами.
Тип 2: Рецепторы, сопряженные с G-белком (метаботропные рецепторы) — это самое большое семейство рецепторов, включающее рецепторы нескольких гормонов и медленных трансмиттеров, например, дофамина, метаботропного глутамата. Они состоят из семи трансмембранных альфа-спиралей. Петли, соединяющие альфа-спирали, образуют внеклеточные и внутриклеточные домены. Сайт связывания для более крупных пептидных лигандов обычно находится во внеклеточном домене, тогда как сайт связывания для более мелких непептидных лигандов часто находится между семью альфа-спиралями и одной внеклеточной петлей. [7] Вышеупомянутые рецепторы связаны с различными внутриклеточными эффекторными системами через G-белки . [8] G-белки представляют собой гетеротримеры, состоящие из 3 субъединиц: α (альфа), β (бета) и γ (гамма). В неактивном состоянии три субъединицы связаны вместе, и α-субъединица связывает GDP. [9] Активация G-белка вызывает конформационное изменение, которое приводит к обмену GDP на GTP. Связывание GTP с α-субъединицей вызывает диссоциацию β- и γ-субъединиц. [10] Кроме того, три субъединицы, α, β и γ, имеют четыре дополнительных основных класса, основанных на их первичной последовательности. К ним относятся G s , Gi , G q и G 12 . [11]
Тип 3: Связанные с киназой и родственные рецепторы (см. « Рецепторная тирозинкиназа » и « Фермент-связанный рецептор ») — они состоят из внеклеточного домена, содержащего сайт связывания лиганда, и внутриклеточного домена, часто с ферментативной функцией, соединенных одной трансмембранной альфа-спиралью. Примером является инсулиновый рецептор .
Тип 4: Ядерные рецепторы – Хотя их называют ядерными рецепторами, на самом деле они расположены в цитоплазме и мигрируют в ядро после связывания со своими лигандами. Они состоят из С-концевой лиганд-связывающей области, основного ДНК-связывающего домена (DBD) и N-концевого домена, который содержит область AF1 (активационная функция 1). Основная область имеет два цинковых пальца, которые отвечают за распознавание последовательностей ДНК, специфичных для этого рецептора. N-конец взаимодействует с другими клеточными факторами транскрипции лиганд-независимым образом; и в зависимости от этих взаимодействий он может изменять связывание/активность рецептора. Примерами таких рецепторов являются рецепторы стероидов и тиреоидных гормонов. [12]
Мембранные рецепторы могут быть выделены из клеточных мембран с помощью сложных процедур экстракции с использованием растворителей , детергентов и/или аффинной очистки .
Связывание лиганда — это равновесный процесс. Лиганды связываются с рецепторами и диссоциируют от них в соответствии с законом действующих масс в следующем уравнении для лиганда L и рецептора R. Скобки вокруг химических видов обозначают их концентрации.
Одной из мер того, насколько хорошо молекула подходит рецептору, является ее связывающее сродство, которое обратно пропорционально константе диссоциации K d . Хорошее соответствие соответствует высокому сродству и низкому K d . Окончательный биологический ответ (например, каскад вторичных мессенджеров , сокращение мышц) достигается только после активации значительного числа рецепторов.
Сродство — это мера тенденции лиганда связываться со своим рецептором. Эффективность — это мера способности связанного лиганда активировать свой рецептор.
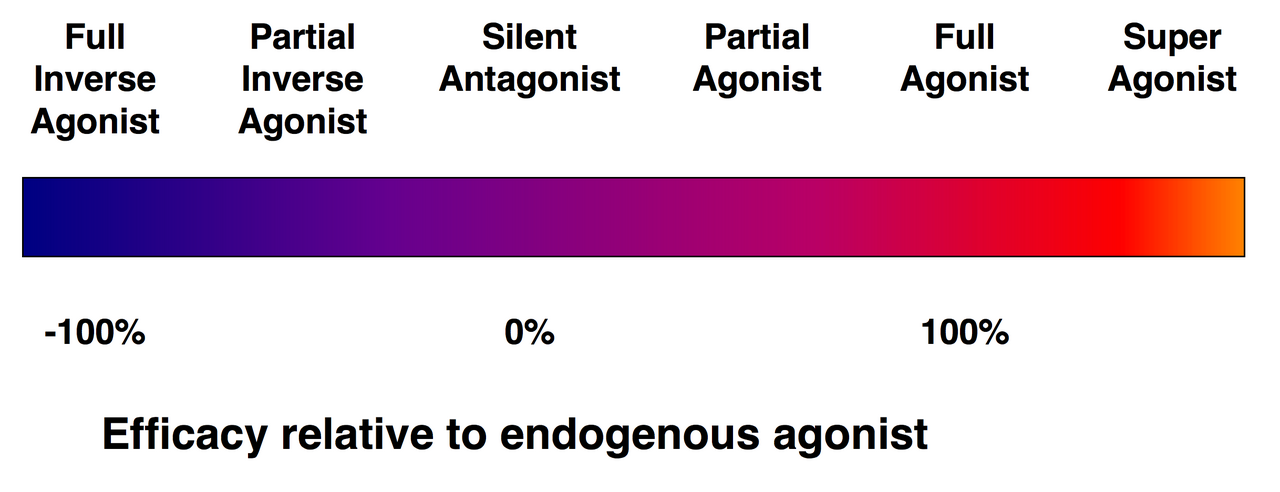
Агонисты против антагонистов
Спектр эффективности лигандов рецепторов.
Не каждый лиганд, который связывается с рецептором, также активирует этот рецептор. Существуют следующие классы лигандов:
(Полные) агонисты способны активировать рецептор и вызывать сильный биологический ответ. Естественный эндогенный лиганд с наибольшей эффективностью для данного рецептора по определению является полным агонистом (100% эффективность).
Частичные агонисты не активируют рецепторы с максимальной эффективностью даже при максимальном связывании, вызывая частичные ответы по сравнению с таковыми у полных агонистов (эффективность от 0 до 100%).
Антагонисты связываются с рецепторами, но не активируют их. Это приводит к блокаде рецепторов, ингибируя связывание агонистов и обратных агонистов. Антагонисты рецепторов могут быть конкурентными (или обратимыми) и конкурировать с агонистом за рецептор, или они могут быть необратимыми антагонистами, которые образуют ковалентные связи (или чрезвычайно высокоаффинные нековалентные связи) с рецептором и полностью блокируют его. Ингибитор протонной помпы омепразол является примером необратимого антагониста. Эффекты необратимого антагонизма могут быть обращены вспять только путем синтеза новых рецепторов.
Аллостерические модуляторы : они не связываются с сайтом связывания агониста рецептора, а вместо этого со специфическими аллостерическими сайтами связывания, через которые они модифицируют эффект агониста. Например, бензодиазепины (БЗД) связываются с сайтом БЗД на рецепторе ГАМК А и усиливают эффект эндогенной ГАМК.
Обратите внимание, что идея агонизма и антагонизма рецепторов относится только к взаимодействию между рецепторами и лигандами, а не к их биологическим эффектам.
Конститутивная деятельность
Рецептор, способный вызывать биологическую реакцию в отсутствие связанного лиганда, как говорят, демонстрирует «конститутивную активность». [13] Конститутивная активность рецептора может быть заблокирована обратным агонистом . Препараты против ожирения римонабант и таранабант являются обратными агонистами каннабиноидного рецептора CB1 , и хотя они вызывали значительную потерю веса, оба были сняты с производства из-за высокой частоты депрессии и тревожности, которые, как полагают, связаны с ингибированием конститутивной активности каннабиноидного рецептора.
Рецептор ГАМК А имеет конститутивную активность и проводит некоторый базальный ток в отсутствие агониста. Это позволяет бета -карболину действовать как обратный агонист и снижать ток ниже базального уровня.
Мутации в рецепторах, приводящие к повышению конститутивной активности, лежат в основе некоторых наследственных заболеваний, таких как преждевременное половое созревание (из-за мутаций в рецепторах лютеинизирующего гормона) и гипертиреоз (из-за мутаций в рецепторах тиреотропного гормона).
Теории взаимодействия лекарств и рецепторов
Занятие
Ранние формы рецепторной теории фармакологии утверждали, что эффект препарата прямо пропорционален количеству занятых рецепторов. [14] Более того, эффект препарата прекращается по мере диссоциации комплекса препарат-рецептор.
Ариенс и Стивенсон ввели термины «сродство» и «эффективность» для описания действия лигандов, связанных с рецепторами. [15] [16]
Сродство : способность лекарственного средства соединяться с рецептором, образуя комплекс лекарственное средство-рецептор.
Эффективность : способность препарата вызывать реакцию после образования комплекса препарат-рецептор.
Ставка
В отличие от принятой теории оккупации , теория скорости предполагает, что активация рецепторов прямо пропорциональна общему числу встреч препарата с его рецепторами в единицу времени. Фармакологическая активность прямо пропорциональна скоростям диссоциации и ассоциации, а не числу занятых рецепторов: [17]
Агонист: препарат с быстрой ассоциацией и быстрой диссоциацией.
Частичный агонист: препарат с промежуточной ассоциацией и промежуточной диссоциацией.
Антагонист: Препарат с быстрой ассоциацией и медленной диссоциацией.
Индуцированная подгонка
Когда лекарство приближается к рецептору, рецептор изменяет конформацию своего участка связывания, образуя комплекс лекарство—рецептор.
Запасные Рецепторы
В некоторых рецепторных системах (например, ацетилхолин в нервно-мышечном соединении в гладких мышцах) агонисты способны вызывать максимальный ответ при очень низких уровнях занятости рецепторов (<1%). Таким образом, эта система имеет запасные рецепторы или резерв рецепторов. Такое расположение обеспечивает экономию производства и высвобождения нейротрансмиттера. [12]
Регуляция рецепторов
Клетки могут увеличивать ( upregulate ) или уменьшать ( downregulate ) количество рецепторов к данному гормону или нейротрансмиттеру , чтобы изменить их чувствительность к различным молекулам. Это механизм обратной связи местного действия .
Изменение конформации рецептора таким образом, что связывание агониста не активирует рецептор. Это наблюдается в случае рецепторов ионных каналов.
Секвестрация рецепторов (интернализация) [18] , например, в случае гормональных рецепторов.
Примеры и лиганды
Лиганды для рецепторов столь же разнообразны, как и их рецепторы. GPCR (7TM) — это особенно обширное семейство, насчитывающее не менее 810 членов. Существуют также LGIC для не менее дюжины эндогенных лигандов, и еще много рецепторов, возможных через различные составы субъединиц. Некоторые общие примеры лигандов и рецепторов включают: [19]
Ионные каналы и рецепторы, сопряженные с G-белком
Некоторые примеры ионотропных (LGIC) и метаботропных (в частности, GPCR) рецепторов показаны в таблице ниже. Главными нейротрансмиттерами являются глутамат и ГАМК; другие нейротрансмиттеры являются нейромодуляторными . Этот список ни в коем случае не является исчерпывающим.
Рецепторы, связанные с ферментами
Рецепторы, связанные с ферментами, включают рецепторные тирозинкиназы (RTK), серин/треонин-специфическую протеинкиназу, как в костном морфогенетическом белке, и гуанилатциклазу, как в рецепторе предсердного натрийуретического фактора. Из RTK было идентифицировано 20 классов, включающих 58 различных RTK. Некоторые примеры приведены ниже:
Внутриклеточные рецепторы
Рецепторы можно классифицировать на основе их механизма или положения в клетке. Ниже приведены 4 примера внутриклеточных LGIC:
Роль в здоровье и болезнях
При генетических нарушениях
Многие генетические нарушения связаны с наследственными дефектами в генах рецепторов. Часто бывает трудно определить, является ли рецептор нефункциональным или гормон вырабатывается на пониженном уровне; это приводит к появлению группы эндокринных нарушений «псевдо-гипо-» , когда кажется, что уровень гормона снижен, хотя на самом деле это рецептор недостаточно реагирует на гормон.
^ abc Guyton, Arthur C.; Hall, John E. (2016). Учебник медицинской физиологии Guyton and Hall. Филадельфия, Пенсильвания: Elsevier Saunders. С. 930–937. ISBN 9781455770052. OCLC 1027900365.
^ Михик, С. Джон; Харрис, Р. Адрон (1997). «ГАМК и рецептор ГАМКА». Alcohol Health and Research World . 21 (2): 127–131. ISSN 0090-838X. PMC 6826832. PMID 15704348 .
^ ab Alberts B, Bray D, Hopkin K, Johnson A, Lewis J, Raff M, Roberts K, Walter P (2014). Essential Cell Biology (четвертое издание). Нью-Йорк, штат Нью-Йорк, США: Science. стр. 534. ISBN978-0-8153-4454-4.
^ Gotti, Cecilia; Marks, Michael. J.; Millar, Neil S.; Wonnacott, Susan (16 сентября 2019 г.). «Никотиновые ацетилхолиновые рецепторы (версия 2019.4)». Руководство по фармакологии IUPHAR/BPS CITE . 2019 (4). doi : 10.2218/gtopdb/F76/2019.4 . Получено 17 ноября 2020 г.
^ Malenka RC, Nestler EJ, Hyman SE (2009). "Глава 9: Автономная нервная система". В Sydor A, Brown RY (ред.). Молекулярная нейрофармакология: основа клинической нейронауки (2-е изд.). Нью-Йорк: McGraw-Hill Medical. стр. 234. ISBN9780071481274. Никотин ... является натуральным алкалоидом табачного растения. Лобелин является натуральным алкалоидом индийского табака. Оба препарата являются агонистами [никотиновых] холинергических рецепторов ...
^ "Curare Drug Information, Professional". Drugs.com . Архивировано из оригинала 16 ноября 2018 года . Получено 8 декабря 2020 года .
^ Congreve M, Marshall F (март 2010). «Влияние структур GPCR на фармакологию и дизайн лекарств на основе структуры». British Journal of Pharmacology . 159 (5): 986–96. doi :10.1111/j.1476-5381.2009.00476.x. PMC 2839258. PMID 19912230 .
^ Qin K, Dong C, Wu G, Lambert NA (август 2011 г.). «Предварительная сборка в неактивном состоянии рецепторов, связанных с G(q), и гетеротримеров G(q)». Nature Chemical Biology . 7 (10): 740–7. doi :10.1038/nchembio.642. PMC 3177959 . PMID 21873996.
^ Зубай, Джеффри (1998). Биохимия 4-е изд . Dubuque, IA: William C Brown Pub. стр. 684. ISBN0697219003.
^ Хамм, Хайди Э.; Олдхэм, Уильям М. (2008). «Гетеротримерная активация G-белка рецепторами, связанными с G-белком». Nature Reviews Molecular Cell Biology . 9 (1). Nature Publishing Group: 60–71. doi : 10.1038/nrm2299. PMID 18043707. S2CID 24267759.
^ ab Rang HP, Dale MM, Ritter JM, Flower RJ, Henderson G (2012). Фармакология Rang & Dale (7-е изд.). Elsevier Churchill Livingstone. ISBN978-0-7020-3471-8.
^ Миллиган Г. (декабрь 2003 г.). «Конститутивная активность и обратные агонисты рецепторов, сопряженных с G-белком: современная перспектива». Молекулярная фармакология . 64 (6): 1271–6. doi :10.1124/mol.64.6.1271. PMID 14645655. S2CID 2454589.
^ Rang, HP (январь 2006). «Концепция рецептора: большая идея фармакологии». British Journal of Pharmacology . 147 (Suppl 1): S9-16. doi :10.1038/sj.bjp.0706457. PMC 1760743. PMID 16402126 .
^ Ariens EJ (сентябрь 1954 г.). «Сродство и внутренняя активность в теории конкурентного ингибирования. I. Проблемы и теория». Archives Internationales de Pharmacodynamie et de Therapie . 99 (1): 32–49. PMID 13229418.
^ Stephenson RP (декабрь 1956 г.). «Модификация теории рецепторов». British Journal of Pharmacology and Chemotherapy . 11 (4): 379–93. doi :10.1111/j.1476-5381.1956.tb00006.x. PMC 1510558. PMID 13383117 .
^ Silverman RB (2004). "3.2.C Теории взаимодействия лекарств и рецепторов". Органическая химия разработки лекарств и действия лекарств (2-е изд.). Амстердам: Elsevier Academic Press. ISBN0-12-643732-7.
^ Boulay G, Chrétien L, Richard DE, Guillemette G (ноябрь 1994 г.). «Кратковременная десенсибилизация рецептора ангиотензина II клеток клубочков надпочечников быка соответствует переходу от состояния высокого к состоянию низкого сродства». Эндокринология . 135 (5): 2130–6. doi :10.1210/en.135.5.2130. PMID 7956936.
^ abcdefghijkl Boulpaep EL, Boron WF (2005). Медицинская физиология: клеточный и молекулярный подход . Сент-Луис, Миссури: Эльзевир Сондерс. п. 90. ИСБН1-4160-2328-3.
^ Waltenbaugh C, Doan T, Melvold R, Viselli S (2008). Иммунология . Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins. стр. 20. ISBN978-0-7817-9543-2.
Внешние ссылки
База данных GPCR IUPHAR и сборник ионных каналов. Архивировано 23.03.2019 на Wayback Machine
Рецептом плазматической мембраны человека Архивировано 15.09.2019 на Wayback Machine
.svg/1280px-Receptor_(Biochemistry).svg.png)
![{\displaystyle {[{\ce {L}}]+[{\ce {R}}]{\ce {<=>[{K_{d}}]}}[{\text{LR}}]} }](https://wikimedia.org/api/rest_v1/media/math/render/svg/8bc59879baab1d42236bb74c50da94e2ceef5afd)