Лиганды соединяются со специфическими белками-рецепторами в зависимости от формы активного центра белка.
Рецептор высвобождает посланника, как только лиганд присоединяется к рецептору.
В биохимии и фармакологии рецепторы — это химические структуры, состоящие из белка , которые принимают и передают сигналы, которые могут быть интегрированы в биологические системы. [1] Эти сигналы обычно представляют собой химические посланники [nb 1] , которые связываются с рецептором и вызывают физиологические реакции , такие как изменение электрической активности клетки . Например, ГАМК , тормозной нейромедиатор , ингибирует электрическую активность нейронов, связываясь с рецепторами ГАМК А. [2] Существует три основных способа классификации действия рецептора: передача сигнала, усиление или интеграция. [3] Ретрансляция отправляет сигнал дальше, амплификация увеличивает эффект одного лиганда , а интеграция позволяет включить сигнал в другой биохимический путь. [3]
Белки-рецепторы можно классифицировать по их местоположению. Рецепторы клеточной поверхности , также известные как трансмембранные рецепторы, включают лиганд-управляемые ионные каналы , рецепторы, связанные с G-белком , и связанные с ферментами рецепторы гормонов . [1] Внутриклеточные рецепторы находятся внутри клетки и включают цитоплазматические рецепторы и ядерные рецепторы . [1] Молекула, которая связывается с рецептором, называется лигандом и может представлять собой белок, пептид (короткий белок) или другую небольшую молекулу , например нейротрансмиттер , гормон , фармацевтический препарат, токсин, ион кальция или части внешней среды. вируса или микроба. Эндогенно произведенное вещество, которое связывается с определенным рецептором, называется его эндогенным лигандом. Например, эндогенным лигандом никотинового рецептора ацетилхолина является ацетилхолин , но он также может активироваться никотином [4] [5] и блокироваться кураре . [6] Рецепторы определенного типа связаны с определенными клеточными биохимическими путями, которые соответствуют сигналу. Хотя в большинстве клеток обнаружено множество рецепторов, каждый рецептор связывается только с лигандами определенной структуры. Это можно сравнить с тем, как замки принимают только ключи определенной формы . Когда лиганд связывается с соответствующим рецептором, он активирует или ингибирует связанный с рецептором биохимический путь, который также может быть узкоспециализированным.
Трансмембранный рецептор: E = внеклеточное пространство; I = внутриклеточное пространство; P = плазматическая мембрана
Структуры рецепторов очень разнообразны и включают, среди прочего, следующие основные категории:
Тип 1: лиганд-управляемые ионные каналы (ионотропные рецепторы). Эти рецепторы обычно являются мишенями быстрых нейротрансмиттеров, таких как ацетилхолин (никотиновый) и ГАМК ; активация этих рецепторов приводит к изменениям в движении ионов через мембрану. Они имеют гетеромерную структуру, в которой каждая субъединица состоит из внеклеточного лиганд-связывающего домена и трансмембранного домена, который включает четыре трансмембранные альфа-спирали . Лигандсвязывающие полости расположены на границе раздела субъединиц.
Тип 2: Рецепторы, связанные с G-белком (метаботропные рецепторы). Это самое большое семейство рецепторов, включающее рецепторы нескольких гормонов и медленных передатчиков, например дофамина, метаботропного глутамата. Они состоят из семи трансмембранных альфа-спиралей. Петли, соединяющие альфа-спирали, образуют внеклеточные и внутриклеточные домены. Сайт связывания более крупных пептидных лигандов обычно расположен во внеклеточном домене, тогда как сайт связывания более мелких непептидных лигандов часто расположен между семью альфа-спиралями и одной внеклеточной петлей. [7] Вышеупомянутые рецепторы связаны с различными внутриклеточными эффекторными системами через G-белки . [8] G-белки представляют собой гетеротримеры, состоящие из трех субъединиц: α (альфа), β (бета) и γ (гамма). В неактивном состоянии три субъединицы связываются вместе, а α-субъединица связывает ВВП. [9] Активация G-белка вызывает конформационные изменения, которые приводят к замене GDP на GTP. Связывание GTP с α-субъединицей вызывает диссоциацию β- и γ-субъединиц. [10] Кроме того, три субъединицы, α, β и γ, имеют дополнительные четыре основных класса на основе их первичной последовательности. К ним относятся G s , G i , G q и G 12 . [11]
Тип 3: Киназно-связанные и родственные рецепторы (см. « Рецептор тирозинкиназы » и « Фермент-связанный рецептор »). Они состоят из внеклеточного домена, содержащего сайт связывания лиганда, и внутриклеточного домена, часто с ферментативной функцией, соединенного между собой одна трансмембранная альфа-спираль. Примером может служить инсулиновый рецептор .
Тип 4: Ядерные рецепторы . Хотя их называют ядерными рецепторами, на самом деле они расположены в цитоплазме и мигрируют в ядро после связывания со своими лигандами. Они состоят из С-концевой лиганд-связывающей области, основного ДНК-связывающего домена (DBD) и N-концевого домена, который содержит область AF1 (функция активации 1). Центральная область имеет два цинковых пальца, которые отвечают за распознавание последовательностей ДНК, специфичных для этого рецептора. N-конец взаимодействует с другими клеточными факторами транскрипции лиганд-независимым образом; и, в зависимости от этих взаимодействий, он может изменять связывание/активность рецептора. Примерами таких рецепторов являются рецепторы стероидных и тироидных гормонов. [12]
Мембранные рецепторы могут быть выделены из клеточных мембран с помощью сложных процедур экстракции с использованием растворителей , детергентов и/или аффинной очистки .
Связывание лигандов представляет собой равновесный процесс. Лиганды связываются с рецепторами и диссоциируют от них в соответствии с законом действия масс в следующем уравнении для лиганда L и рецептора R. Скобки вокруг химических видов обозначают их концентрации.
Одним из показателей того, насколько хорошо молекула соответствует рецептору , является ее аффинность связывания, которая обратно пропорциональна константе диссоциации Kd . Хорошее соответствие соответствует высокому сродству и низкому K d . Окончательный биологический ответ (например, каскад вторичных мессенджеров , сокращение мышц) достигается только после активации значительного числа рецепторов.
Сродство является мерой склонности лиганда связываться со своим рецептором. Эффективность — это мера способности связанного лиганда активировать свой рецептор.
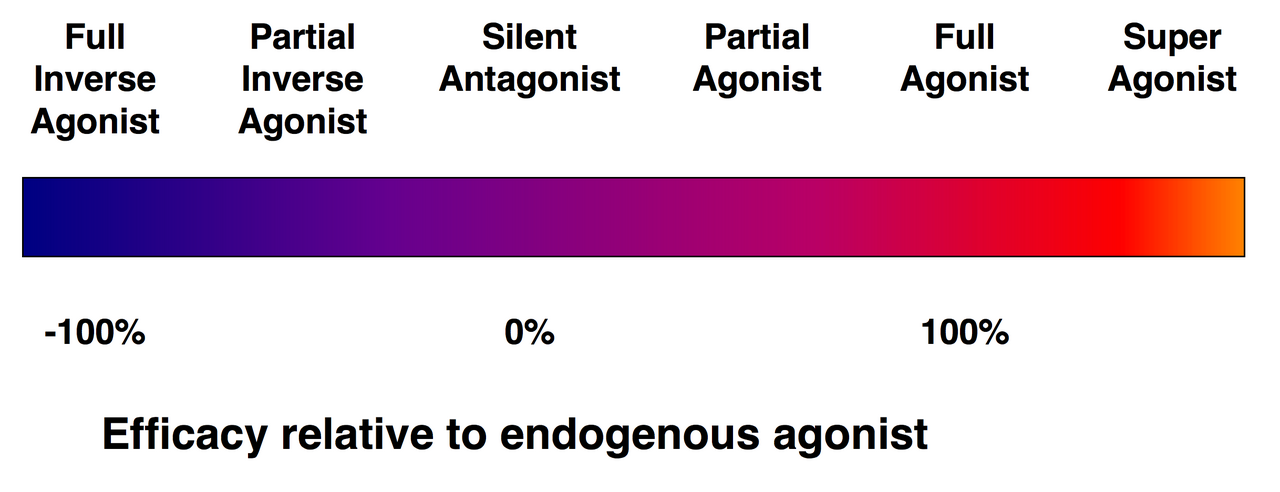
Агонисты против антагонистов
Спектр эффективности рецепторных лигандов.
Не каждый лиганд, который связывается с рецептором, также активирует этот рецептор. Существуют следующие классы лигандов:
(Полные) агонисты способны активировать рецептор и вызывать сильный биологический ответ. Природный эндогенный лиганд с наибольшей эффективностью для данного рецептора по определению является полным агонистом (100% эффективность).
Частичные агонисты не активируют рецепторы с максимальной эффективностью, даже при максимальном связывании, вызывая частичные ответы по сравнению с полными агонистами (эффективность от 0 до 100%).
Антагонисты связываются с рецепторами, но не активируют их. Это приводит к блокаде рецепторов, ингибируя связывание агонистов и обратных агонистов. Антагонисты рецептора могут быть конкурентными (или обратимыми) и конкурировать с агонистом за рецептор, или же они могут быть необратимыми антагонистами, которые образуют ковалентные связи (или нековалентные связи с чрезвычайно высоким сродством) с рецептором и полностью блокируют его. Ингибитор протонной помпы омепразол является примером необратимого антагониста. Эффекты необратимого антагонизма можно обратить вспять только путем синтеза новых рецепторов.
Обратные агонисты снижают активность рецепторов за счет ингибирования их конститутивной активности (отрицательная эффективность).
Аллостерические модуляторы : они связываются не с сайтом связывания агониста рецептора, а со специфическими сайтами аллостерического связывания, посредством которых они изменяют эффект агониста. Например, бензодиазепины (БЗД) связываются с участком БЗД на рецепторе ГАМК А и усиливают эффект эндогенной ГАМК.
Обратите внимание, что идея агонизма и антагонизма рецепторов относится только к взаимодействию между рецепторами и лигандами, а не к их биологическим эффектам.
Учредительная деятельность
Говорят, что рецептор, который способен вызывать биологический ответ в отсутствие связанного лиганда, проявляет «конститутивную активность». [13] Конститутивная активность рецептора может быть заблокирована обратным агонистом . Препараты против ожирения римонабант и таранабант являются обратными агонистами каннабиноидного рецептора CB1 , и хотя они вызывают значительную потерю веса, оба они были отменены из-за высокой частоты депрессии и тревоги, которые, как полагают, связаны с ингибированием конститутивной активности каннабиноидный рецептор.
Мутации рецепторов, приводящие к повышению конститутивной активности, лежат в основе некоторых наследственных заболеваний, таких как преждевременное половое созревание (из-за мутаций в рецепторах лютеинизирующего гормона) и гипертиреоз (из-за мутаций в рецепторах тиреотропного гормона).
Теории взаимодействия лекарств и рецепторов
Занятие
Ранние формы фармакологической теории рецепторов утверждали, что эффект лекарства прямо пропорционален количеству занятых рецепторов. [14] Более того, эффект препарата прекращается по мере диссоциации комплекса лекарство-рецептор.
Ариенс и Стивенсон ввели термины «аффинность» и «эффективность» для описания действия лигандов, связанных с рецепторами. [15] [16]
Сродство : способность лекарства соединяться с рецептором с образованием комплекса лекарство-рецептор.
Эффективность : способность препарата инициировать ответ после образования комплекса лекарство-рецептор.
Ставка
В отличие от принятой теории оккупации , теория скорости предполагает, что активация рецепторов прямо пропорциональна общему числу встреч лекарства с его рецепторами в единицу времени. Фармакологическая активность прямо пропорциональна скорости диссоциации и ассоциации, а не количеству занятых рецепторов: [17]
Агонист: препарат с быстрой ассоциацией и быстрой диссоциацией.
Частичный агонист: препарат с промежуточной ассоциацией и промежуточной диссоциацией.
Антагонист: препарат с быстрой ассоциацией и медленной диссоциацией.
Индуцированная посадка
Когда лекарство приближается к рецептору, рецептор меняет конформацию своего сайта связывания, образуя комплекс лекарство-рецептор.
Запасные рецепторы
В некоторых рецепторных системах (например, ацетилхолин в нервно-мышечном соединении гладких мышц) агонисты способны вызывать максимальный ответ при очень низких уровнях занятости рецептора (<1%). Таким образом, эта система имеет запасные рецепторы или резерв рецепторов. Такое расположение обеспечивает экономику производства и высвобождения нейромедиаторов. [12]
Регуляция рецепторов
Клетки могут увеличивать ( активировать ) или уменьшать ( понижать ) количество рецепторов к данному гормону или нейротрансмиттеру , чтобы изменить их чувствительность к различным молекулам. Это локально действующий механизм обратной связи .
Изменение конформации рецептора, при котором связывание агониста не активирует рецептор. Это видно по рецепторам ионных каналов.
Секвестрация рецепторов (интернализация), [18], например, в случае рецепторов гормонов.
Примеры и лиганды
Лиганды рецепторов столь же разнообразны, как и их рецепторы. GPCR (7TM) представляют собой особенно обширную семью, насчитывающую не менее 810 членов. Существуют также LGIC , по крайней мере, для дюжины эндогенных лигандов и многих других рецепторов, возможных благодаря разному составу субъединиц. Некоторые распространенные примеры лигандов и рецепторов включают: [19]
Ионные каналы и рецепторы, связанные с G-белком
Некоторые примеры ионотропных (LGIC) и метаботропных (в частности, GPCR) рецепторов показаны в таблице ниже. Главными нейротрансмиттерами являются глутамат и ГАМК; другие нейротрансмиттеры являются нейромодулирующими . Этот список ни в коем случае не является исчерпывающим.
Рецепторы, связанные с ферментами
Рецепторы, связанные с ферментами, включают рецепторные тирозинкиназы (RTK), серин/треонин-специфическую протеинкиназу, как в костном морфогенетическом белке, и гуанилатциклазу, как в рецепторе предсердного натрийуретического фактора. Из RTK было идентифицировано 20 классов, членами которых являются 58 различных RTK. Некоторые примеры показаны ниже:
Внутриклеточные рецепторы
Рецепторы можно классифицировать на основе их механизма или положения в клетке. Ниже показаны 4 примера внутриклеточных LGIC:
Роль в здоровье и болезни
При генетических нарушениях
Многие генетические заболевания связаны с наследственными дефектами генов-рецепторов. Часто трудно определить, не функционирует ли рецептор или гормон вырабатывается в пониженном уровне; это приводит к возникновению «псевдо-гипо-» группы эндокринных расстройств , при которых, по-видимому, наблюдается снижение гормонального уровня, хотя на самом деле именно рецептор недостаточно реагирует на гормон.
^ abc Гайтон, Артур К.; Холл, Джон Э. (2016). Учебник медицинской физиологии Гайтона и Холла. Филадельфия, Пенсильвания: Эльзевир Сондерс. стр. 930–937. ISBN 9781455770052. ОСЛК 1027900365.
^ Михич, С. Джон; Харрис, Р. Адрон (1997). «ГАМК и рецептор ГАМК». Алкоголь, здоровье и мир исследований . 21 (2): 127–131. ISSN 0090-838X. ПМК 6826832 . ПМИД 15704348.
^ ab Альбертс Б., Брей Д., Хопкин К., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2014). Основная клеточная биология (Четвертое изд.). Нью-Йорк, штат Нью-Йорк, США: Garland Science. п. 534. ИСБН978-0-8153-4454-4.
^ Готти, Сесилия; Маркс, Майкл. Дж.; Миллар, Нил С.; Воннакотт, Сьюзен (16 сентября 2019 г.). «Никотиновые рецепторы ацетилхолина (версия 2019.4)». Руководство IUPHAR/BPS по фармакологии CITE . 2019 (4). дои : 10.2218/gtopdb/F76/2019.4 . Проверено 17 ноября 2020 г.
^ Маленка Р.К., Нестлер Э.Дж., Хайман С.Е. (2009). «Глава 9: Автономная нервная система». В Сидоре А., Брауне Р.Ю. (ред.). Молекулярная нейрофармакология: фонд клинической неврологии (2-е изд.). Нью-Йорк: McGraw-Hill Medical. п. 234. ИСБН9780071481274. Никотин... является природным алкалоидом табачного растения. Лобелин – природный алкалоид индийского табака. Оба препарата являются агонистами никотиновых холинергических рецепторов...
^ "Информация о лекарствах Curare, Professional" . Наркотики.com . Проверено 8 декабря 2020 г.
^ Конгрив М., Маршалл Ф. (март 2010 г.). «Влияние структур GPCR на фармакологию и разработку лекарств на основе структуры». Британский журнал фармакологии . 159 (5): 986–96. дои : 10.1111/j.1476-5381.2009.00476.x. ПМЦ 2839258 . ПМИД 19912230.
^ Цинь К., Донг С., Ву Г, Ламберт Н.А. (август 2011 г.). «Предварительная сборка G(q)-связанных рецепторов и G(q)-гетеротримеров» в неактивном состоянии». Химическая биология природы . 7 (10): 740–7. дои : 10.1038/nchembio.642. ПМК 3177959 . ПМИД 21873996.
^ Зубай, Джеффри (1998). Биохимия 4-е изд . Дубьюк, Айова: Паб William C Brown. п. 684. ИСБН0697219003.
^ Хамм, Хайди Э.; Олдхэм, Уильям М. (2008). «Активация гетеротримерного G-белка рецепторами, связанными с G-белком». Nature Reviews Молекулярно-клеточная биология . Издательская группа «Природа». 9 (1): 60–71. дои : 10.1038/nrm2299. PMID 18043707. S2CID 24267759.
^ ab Rang HP, Дейл М.М., Риттер Дж.М., Флауэр Р.Дж., Хендерсон Дж. (2012). Фармакология Ранг и Дейла (7-е изд.). Эльзевир Черчилль Ливингстон. ISBN978-0-7020-3471-8.
^ Миллиган Дж. (декабрь 2003 г.). «Конститутивная активность и обратные агонисты рецепторов, связанных с G-белком: современная перспектива». Молекулярная фармакология . 64 (6): 1271–6. дои :10.1124/моль.64.6.1271. PMID 14645655. S2CID 2454589.
^ Ранг, HP (январь 2006 г.). «Концепция рецептора: большая идея фармакологии». Британский журнал фармакологии . 147 (Приложение 1): С9-16. дои : 10.1038/sj.bjp.0706457. ПМК 1760743 . ПМИД 16402126.
^ Ariens EJ (сентябрь 1954 г.). «Сродство и внутренняя активность в теории конкурентного торможения. I. Проблемы и теория». Archives Internationales de Pharmacodynamie et de Therapie . 99 (1): 32–49. ПМИД 13229418.
^ Стивенсон Р.П. (декабрь 1956 г.). «Модификация теории рецепторов». Британский журнал фармакологии и химиотерапии . 11 (4): 379–93. doi :10.1111/j.1476-5381.1956.tb00006.x. ПМК 1510558 . ПМИД 13383117.
^ Сильверман РБ (2004). «3.2.C Теории взаимодействия лекарств и рецепторов». Органическая химия разработки и действия лекарств (2-е изд.). Амстердам: Elsevier Academic Press. ISBN0-12-643732-7.
^ Буле Г., Кретьен Л., Ричард Д.Е., Гиймет Дж. (ноябрь 1994 г.). «Кратковременная десенсибилизация рецептора ангиотензина II клубочковых клеток надпочечников крупного рогатого скота соответствует переходу от состояния высокого сродства к состоянию низкого». Эндокринология . 135 (5): 2130–6. дои : 10.1210/en.135.5.2130. ПМИД 7956936.
^ abcdefghijkl Boulpaep EL, Boron WF (2005). Медицинская физиология: клеточный и молекулярный подход . Сент-Луис, Миссури: Эльзевир Сондерс. п. 90. ИСБН1-4160-2328-3.
^ Уолтенбо С., Доан Т., Мелволд Р., Визелли С. (2008). Иммунология . Филадельфия: Wolters Kluwer Health/Lippincott Williams & Wilkins. п. 20. ISBN978-0-7817-9543-2.
Внешние ссылки
База данных IUPHAR GPCR и сборник ионных каналов, заархивированные 23 марта 2019 г. на Wayback Machine.
Рецептом плазматической мембраны человека. Архивировано 15 сентября 2019 г. в Wayback Machine.
.svg/1280px-Receptor_(Biochemistry).svg.png)
![{\displaystyle {[{\ce {L}}]+[{\ce {R}}]{\ce {<=>[{K_{d}}]}}[{\text{LR}}]} }](https://wikimedia.org/api/rest_v1/media/math/render/svg/8bc59879baab1d42236bb74c50da94e2ceef5afd)