
Зрительные опсины позвоночных представляют собой подкласс ресничных опсинов и опосредуют зрение у позвоночных. Они включают опсины в палочках и колбочках человека . Их часто сокращают до опсинов , поскольку они были первыми открытыми опсинами и до сих пор являются наиболее широко изученными опсинами. [1]
Опсин относится строго к апопротеину (без связанного с сетчаткой). Когда опсин связывает ретиналь с образованием голопротеина , его называют ретинилиденовым белком . Однако это различие часто игнорируется, и опсин может свободно относиться к обоим (независимо от того, связан ли сетчатка).
Опсины представляют собой рецепторы, связанные с G-белком (GPCR), и должны связываться с ретиналем — обычно 11- цис -ретиналем — чтобы быть фоточувствительными, поскольку сетчатка действует как хромофор . Когда белок ретинилиден поглощает фотон, сетчатка изомеризуется и высвобождается опсином. Процесс, следующий за изомеризацией и обновлением сетчатки, известен как зрительный цикл . Свободный 11- цис- ретиналь светочувствителен и обладает собственной спектральной чувствительностью 380 нм. [2] Однако, чтобы запустить каскад фототрансдукции , процесс, лежащий в основе зрительного сигнала, сетчатка должна быть связана с опсином, когда он изомеризуется. Белок ретинилиден обладает спектральной чувствительностью, отличной от чувствительности свободного ретиналя и зависит от последовательности опсина.
Хотя опсины могут связывать только сетчатку, существует две формы ретиналя, которые могут действовать как хромофор для зрительных опсинов позвоночных:
Животные, живущие на суше, и морские рыбы формируют свои зрительные пигменты исключительно с помощью ретиналя 1. Однако многие пресноводные рыбы и земноводные также могут образовывать зрительные пигменты с помощью ретиналя 2, в зависимости от активации фермента ретиналь-3,4-десатуразы (GO:0061899). ). Многие из этих видов могут переключаться между этими хромофорами в течение своего жизненного цикла, чтобы адаптироваться к меняющейся среде обитания. [3] [4]
Изомеризация 11- цис -ретиналя в полностью транс -ретиналь под действием света вызывает конформационные изменения в белке, которые активируют путь фототрансдукции .
Существует два класса зрительных опсинов позвоночных, которые различаются по тому, экспрессируются ли они в палочковидных или колбочковых фоторецепторах.
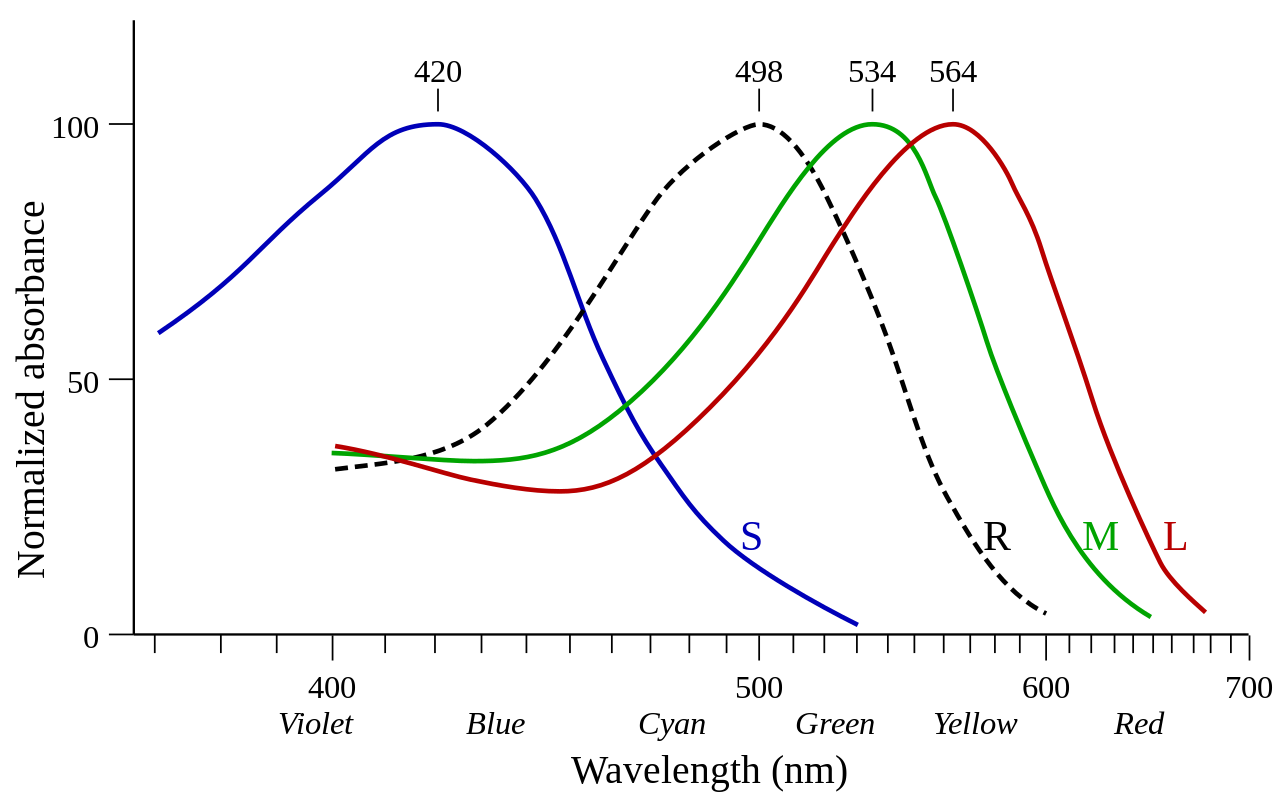
Опсины, экспрессируемые в колбочках, называются конусными опсинами. [1] Опсины колбочек называются фотопсинами , когда они не связаны с сетчаткой, и йодопсинами, когда они связаны с сетчаткой. [1] Опсины колбочек обеспечивают фотопическое зрение (дневной свет). Опсины колбочек далее подразделяются в зависимости от спектральной чувствительности их йодопсина, а именно длины волны, при которой наблюдается наибольшее поглощение света ( λ max ). [7]
Опсины, экспрессируемые в палочковых клетках, называются палочковидными опсинами. Палочки-опсины называются скотопсинами , когда они не связаны с ретиналем, и родопсинами или порфиропсинами , когда они связаны с сетчаткой (1 и 2 соответственно). Опсины палочек обеспечивают скотопическое зрение (тусклый свет). [8] По сравнению с опсинами колбочек спектральная чувствительность родопсина довольно стабильна, не отклоняясь далеко от 500 нм у любого позвоночного.
Современные позвоночные обычно имеют четыре класса опсинов колбочек (LWS, SWS1, SWS2 и Rh2), а также один класс опсинов палочек (родопсин, Rh1), все из которых были унаследованы от ранних предков позвоночных. Согласно кладограмме справа , эти пять классов зрительных опсинов позвоночных возникли в результате серии дупликаций генов, начиная с LWS и заканчивая Rh1. С тех пор каждый класс превратился в множество вариантов. [9] [10] Эволюционные взаимоотношения, выведенные с использованием аминокислотной последовательности опсинов, часто используются для отнесения опсинов колбочек к соответствующему классу. [1] Млекопитающие потеряли классы Rh2 и SWS2 во время ночного «узкого места» . Позже предки приматов развили два опсина LWS (LWS и MWS), в результате чего у людей осталось 4 зрительных опсина в 3 классах.
Джордж Уолд получил Нобелевскую премию по физиологии и медицине 1967 года за свои эксперименты 1950-х годов, которые показали разницу в поглощении этих фотопсинов (см. изображение). [11]
Как видно из рис. 11a, MSP малопригоден для определения спектральной чувствительности колбочек, за исключением случаев, близких к фотопигменту λmax. Большие расхождения между MSP и другими оценками спектральной чувствительности конусов возникают из-за малого отношения сигнал/шум измерений MSP.