Acanthostega (что означает «колючая крыша») — вымерший род стебленогих четвероногих , один из первых позвоночных животных , имеющих узнаваемые конечности . Он появился в позднем девонском периоде ( фаменский ярус) около 365 миллионов лет назад и был анатомически промежуточным между лопастеперыми рыбами и теми, которые были полностью способны выходить на сушу. [1] [2]

Окаменелые останки , как правило, хорошо сохранились. Знаменитая окаменелость , благодаря которой была открыта значимость этого вида, была найдена Дженнифер А. Клэк в Восточной Гренландии в 1987 году, хотя фрагменты черепа были обнаружены в 1933 году Гуннаром Сэве-Сёдербергом и Эриком Ярвиком .

Acanthostega длиной 60 см (24 дюйма) имела восемь пальцев на каждой руке (количество пальцев на ногах неясно), соединенных перепонками, у нее отсутствовали запястья, и в целом она была плохо приспособлена для ходьбы по суше. У нее также были удивительно рыбоподобные плечо и передняя конечность. [3] Передние конечности Acanthostega не могли сгибаться вперед в локте, и поэтому не могли быть приведены в положение для переноса веса , что, по-видимому, больше подходило для гребли или для удержания водных растений. Acanthostega является самым ранним стебленогим четвероногим, у которого наблюдался сдвиг в локомоторном доминировании с плечевого пояса на тазовый. [4]
Существует множество морфологических изменений, которые позволили тазовому поясу Acanthostega стать несущей конструкцией. В более предковых состояниях две стороны пояса не были прикреплены. У Acanthostega есть контакт между двумя сторонами и сращение пояса с крестцовым ребром позвоночника. Эти слияния сделали бы тазовую область более мощной и приспособленной для противодействия силе тяжести, когда она не поддерживалась плавучестью водной среды. [4] У него были внутренние жабры, которые были покрыты, как у рыб. У него также были легкие, но его ребра были слишком короткими, чтобы поддерживать его грудную полость над водой. [2]
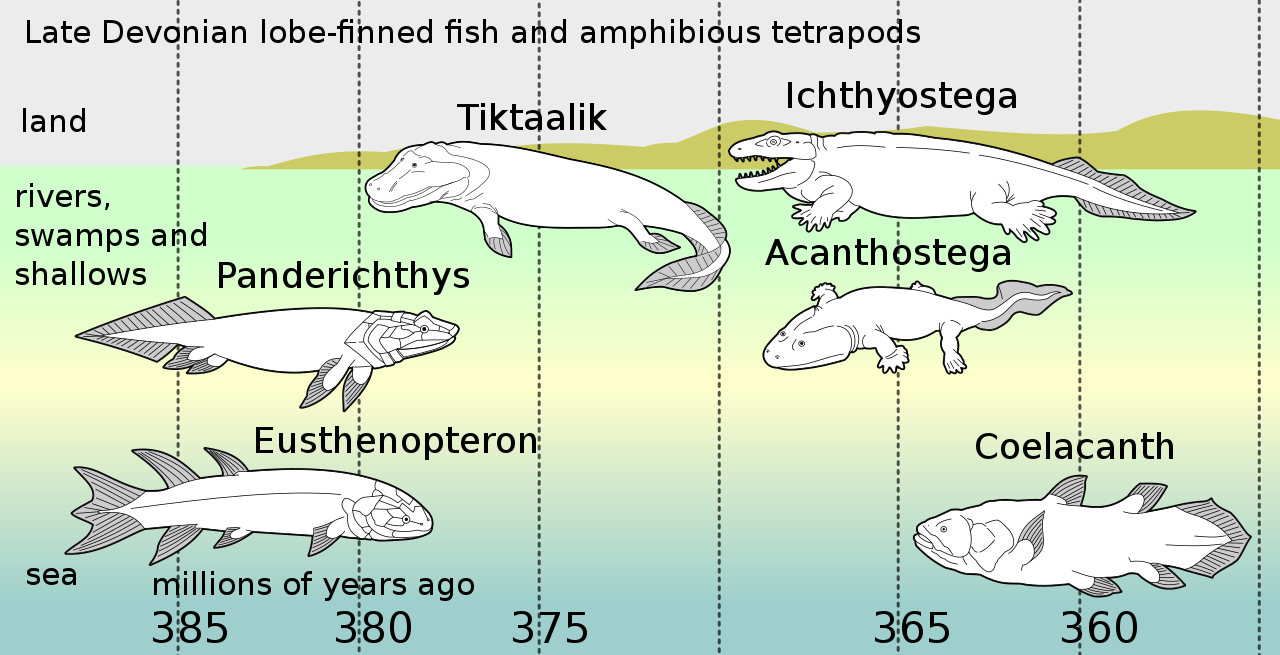
Acanthostega рассматривается как часть широко распространенной эволюционной радиации в позднем девонском периоде, начавшейся с чисто водных плавниковых тетраподоморфов, а их преемники демонстрировали повышенную способность к дыханию воздухом и связанную с этим адаптацию челюстей и жабр, а также более мускулистую шею, позволяющую более свободно двигать головой, чем у рыб, и использование плавников для подъема тела рыбы. [2] Эти особенности демонстрируются более ранним Tiktaalik , который, как и Ichthyostega, демонстрировал признаки большей способности передвигаться по суше, но, как полагают, был в основном водным. [2]
В позднедевонском видообразовании позвоночных потомки пелагических лопастеперых рыб , таких как Eusthenopteron , продемонстрировали последовательность адаптаций: Panderichthys , приспособленный к илистым мелководьям; Tiktaalik с плавниками, похожими на конечности, которые позволяли ему выходить на сушу; стебленогие четвероногие в болотах, заполненных водорослями, такие как Acanthostega , у которой были восьмипалые ноги; и Ichthyostega , с полными конечностями. Их потомки также включали пелагических лопастеперых рыб, таких как виды целакантов .

Было сделано предположение, что Acanthostega , вероятно, жила в неглубоких, заросших водорослями болотах, ее ноги, по-видимому, были адаптированы к этим специфическим экосистемам. Помимо наличия конечностей, она не была никак приспособлена для хождения по суше. Дженнифер А. Клэк интерпретирует это как доказательство того, что Acanthostega была в первую очередь водным животным, произошедшим от рыб, которые никогда не покидали море, и что специализации линии тетрапод были экзаптациями : чертами, которые позже будут полезны для наземной жизни, даже если они возникли для другой цели. В тот период листопадные растения процветали и ежегодно сбрасывали листья в воду, привлекая мелкую добычу на теплые, бедные кислородом мелководья, в которых было трудно плавать более крупным рыбам ; Клэк замечает, как нижняя челюсть Acanthostega показывает изменение по сравнению с рыбами, имеющими два ряда зубов, с большим количеством мелких зубов во внешнем ряду и двумя большими клыками и несколькими более мелкими зубами во внутреннем ряду. Это различие, вероятно, соответствует переходу стволовых тетрапод от питания исключительно в воде к питанию головой над водой или на суше. [2]

Исследования, основанные на анализе морфологии швов на черепе Acanthostega, показывают, что вид мог кусать добычу на краю воды или около нее. Марки и Маршалл сравнили череп с черепами рыб, которые используют всасывающее питание в качестве основного метода захвата добычи, и существ, которые, как известно, использовали прямое укусывание добычи, типичное для наземных животных . Их результаты показывают, что Acanthostega была адаптирована к тому, что они называют наземным стилем питания, что решительно подтверждает гипотезу о том, что наземный способ питания впервые появился у водных животных. Если это верно, это показывает животное, специализированное для охоты и жизни на мелководье на линии между сушей и водой. [5]

Хотя обычно считается, что они более базальны, чем ихтиостега , возможно, что акантостега на самом деле была более производной. Поскольку акантостега напоминает молодь ихтиостеги и показывает гораздо меньше различий от молоди до взрослых особей, чем последние, было высказано предположение, что акантостега может происходить от неотенической линии. Хотя, по-видимому, она провела всю свою жизнь в воде, ее плечевая кость также демонстрирует черты, которые напоминают черты более поздних, полностью наземных стволовых тетрапод (плечевая кость у ихтиостеги в некоторой степени произошла от грудных и тазовых плавниковых костей более ранних рыб и гомологична им). Это может указывать на то, что позвоночные развили наземные черты раньше, чем предполагалось ранее, и многократно независимо друг от друга. [6] Мускульные шрамы на передних конечностях Acanthostega были похожи на шрамы у кроновых тетрапод, что позволяет предположить, что он произошел от предка, у которого было больше наземных приспособлений, чем у него самого. [7]
Гистологическое исследование Acanthostega humeri, проведенное с помощью синхротронного сканирования, показывает, что животное созревало медленно. Некоторые особи достигали половой зрелости (на основе полностью окостеневшей плечевой кости) в возрасте более шести лет, а ископаемые останки взрослых особей встречаются гораздо реже, чем детенышей. Поздняя оссификация плечевой кости подтверждает полностью водный образ жизни Acanthostega . Практически нет никакой корреляции между размером плечевой кости и зрелостью, что позволяет предположить, что существовали значительные различия в размерах среди особей одного возраста. Это может быть связано с конкурентным давлением, различными адаптивными стратегиями или даже половым диморфизмом . Однако небольшой размер выборки не позволяет распознать бимодальное распределение , которое могло бы подтвердить последнюю гипотезу. [8]
{kind=link}