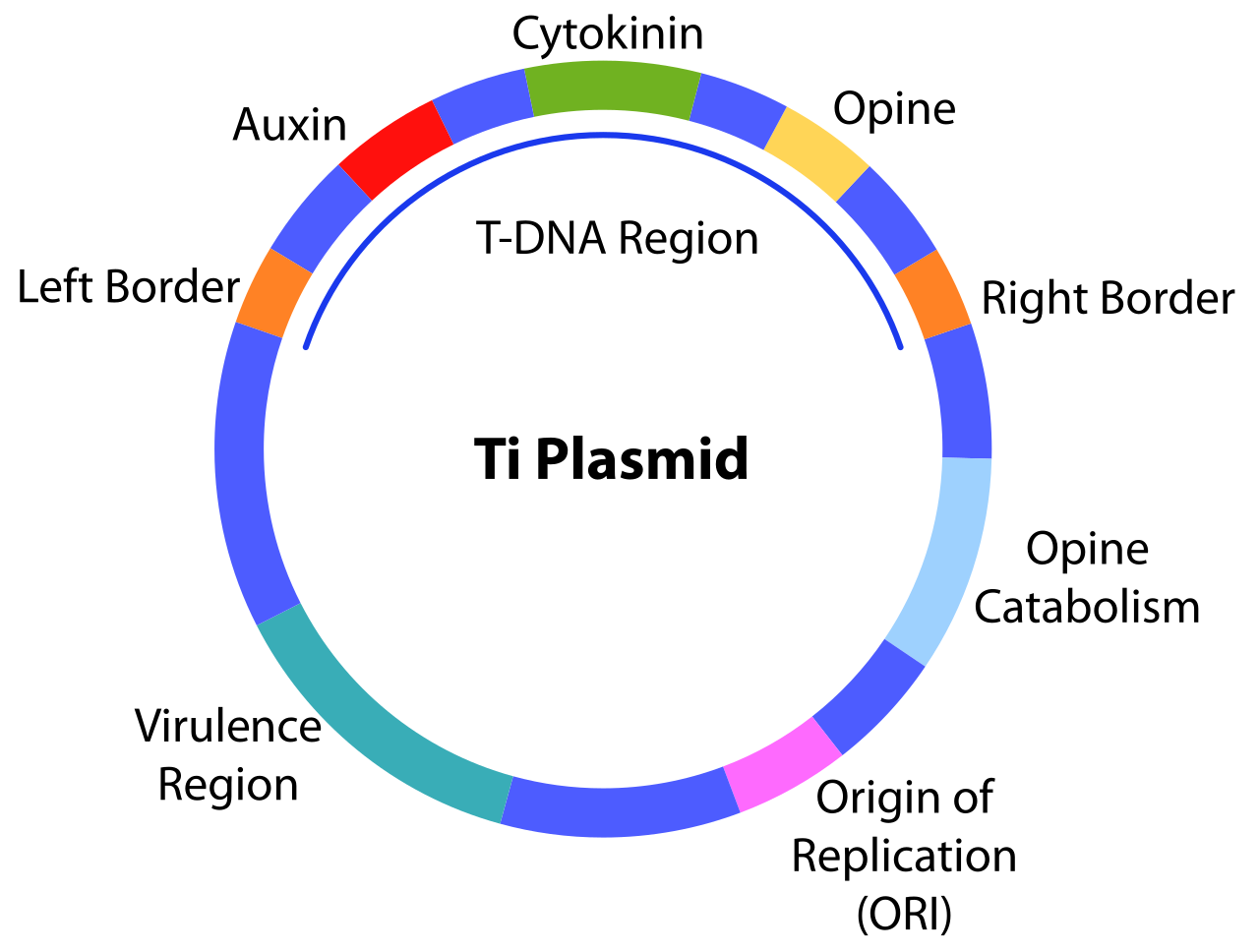
Плазмида , индуцирующая опухоль (Ti), — это плазмида, обнаруженная в патогенных видах Agrobacterium , включая A. tumefaciens , A. rhizogenes , A. rubi и A. vitis .
Эволюционно плазмида Ti является частью семейства плазмид, переносимых многими видами Alphaproteobacteria . Члены этого семейства плазмид определяются наличием консервативной области ДНК, известной как генная кассета repABC , которая опосредует репликацию плазмиды, разделение плазмиды на дочерние клетки во время деления клетки , а также поддержание плазмиды при низком количестве копий в клетке. [1] Сами плазмиды Ti сортируются по различным категориям в зависимости от типа молекулы, или опина , они позволяют бактериям расщепляться в качестве источника энергии. [2]
Наличие этой плазмиды Ti необходимо для того, чтобы бактерии вызывали заболевание корончатого галла у растений. [1] Этому способствуют определенные важные области в плазмиде Ti, включая область vir , которая кодирует гены вирулентности, и область переноса ДНК (T-ДНК), которая является частью плазмиды Ti, которая переносится посредством конъюгации в клетки растения-хозяина после того, как бактерии ощущают место повреждения. Эти области обладают характеристиками, которые позволяют доставлять T-ДНК в клетки растения-хозяина, и могут модифицировать клетку растения-хозяина, вызывая синтез молекул, таких как фитогормоны (например, ауксины , цитокинины ) и опины, а также образование опухолей корончатого галла. [1]
Поскольку область T-ДНК плазмиды Ti может переноситься из бактерий в растительные клетки, это открыло новые возможности для переноса ДНК между царствами и послужило толчком к проведению большого количества исследований плазмиды Ti и ее возможного использования в биоинженерии.
Плазмида Ti является членом семейства плазмид RepABC, обнаруженного в Alphaproteobacteria. [3] Эти плазмиды часто имеют относительно большой размер, от 100 кб до 2 Мб. Их также часто называют репликонами , поскольку их репликация начинается в одном месте. Члены этого семейства имеют характерную кассету генов repABC . [4] Другим заметным членом этого семейства является плазмида, индуцирующая образование корней (Ri), переносимая A. rhizogenes , которая вызывает другое заболевание растений, известное как болезнь волосатых корней. [1]
Ключевой особенностью плазмид Ti является их способность управлять производством опинов, которые являются производными различных аминокислот или фосфатов сахаров , в клетках растений-хозяев. Эти опины затем могут быть использованы в качестве питательного вещества для инфицирующих бактерий, которые катаболизируют соответствующие опины с помощью генов, закодированных в плазмиде Ti.
Соответственно, плазмиды Ti были классифицированы на основе типа опина, который они катаболизируют, а именно: нопалиновый , октопиновый или маннитильный типы, которые являются производными аминокислот, или агроцинопиновый тип, которые являются производными фосфата сахара. [1]
Выявление A. tumefaciens как причины опухолей желчных пузырьков у растений проложило путь к пониманию молекулярной основы заболевания корончатым галлом. [5]
Первое указание на генетический эффект на клетки растения-хозяина появилось в 1942-1943 годах, когда было обнаружено, что в растительных клетках вторичных опухолей не было никаких бактериальных клеток. Однако эти опухолевые клетки обладали способностью продуцировать опины, метаболизируемые инфицирующим бактериальным штаммом. [6] Важно то, что продуцирование соответствующих опинов происходило независимо от вида растения и иногда только в тканях корончатого галла, что указывает на то, что бактерии перенесли некоторый генетический материал в клетки растения-хозяина, чтобы обеспечить синтез опинов. [5]
Однако, как и в какой степени происходил перенос ДНК, оставалось открытым вопросом. Добавление только ДНК A. tumefaciens не вызывало опухолей у растений, [7] в то время как было обнаружено, что очень мало ДНК A. tumefaciens интегрировалось в геном клетки растения-хозяина. [8] Добавление дезоксирибонуклеаз (ДНКаз) для деградации ДНК также не смогло предотвратить образование и рост опухолей растений. [9] Это предполагает, что лишь малая часть ДНК A. tumefaciens , если вообще какая-либо, переносится в клетку растения-хозяина, чтобы вызвать заболевание, и, если ДНК действительно переносится от бактерий к растению, это должно происходить защищенным образом.
Впоследствии было обнаружено, что онкогенные штаммы бактерий способны превращать непатогенные бактерии в патогенные посредством процесса конъюгации, при котором гены, ответственные за вирулентность, переносились в непатогенные клетки. [10] Роль плазмиды в этой патогенной способности была дополнительно подтверждена, когда крупные плазмиды были обнаружены только в патогенных бактериях, но не в авирулентных бактериях. [11] В конце концов, было установлено обнаружение частей бактериальных плазмид в клетках растений-хозяев, что подтвердило, что это был генетический материал, ответственный за генетический эффект инфекции. [12]
С идентификацией плазмиды Ti было проведено много исследований для определения характеристик плазмиды Ti и того, как генетический материал передается от Agrobacterium к растению-хозяину. Некоторые заметные ранние вехи в исследованиях плазмид Ti включают картирование плазмиды Ti в 1978 году и изучение сходства последовательностей между различными плазмидами Ti в 1981 году. [13] [14]
В период с 1980 по 2000 год также проводилась характеристика области T-ДНК и области «vir». Исследования области T-ДНК определили процесс их переноса и идентифицировали гены, позволяющие синтезировать растительные гормоны и опины. [15] Отдельно ранние работы были направлены на определение функций генов, закодированных в области «vir» — они были в целом разделены на те, которые допускали взаимодействие бактерии с хозяином, и те, которые обеспечивали доставку T-ДНК. [2]

Репликация, разделение и поддержание плазмиды Ti зависят от генной кассеты repABC , которая в основном состоит из трех генов: repA , repB и repC . Каждый из repA и repB кодирует белки, участвующие в разделении плазмиды, в то время как repC кодирует инициатор репликации. [1] Эти гены экспрессируются с 4 различных промоторов, расположенных выше repA . repE кодирует небольшую антисмысловую РНК и расположен между repB и repC . [4] Кроме того, в кассете repABC присутствуют сайт разделения ( parS ) и точка начала репликации ( oriV ) . [1]
Репликация плазмиды Ti управляется белком-инициатором RepC ( P05684 ), который обладает двумя белковыми доменами : N-концевым доменом (NTD), который связывается с ДНК, и C-концевым доменом (CTD). Мутационный анализ показал, что без функционального белка RepC плазмида Ti неспособна реплицироваться. [4] Между тем, последовательность oriV имеет длину около 150 нуклеотидов и находится в гене repC . [3] Лабораторные эксперименты показали, что белок RepC связывается с этой областью, что предполагает его роль в качестве источника репликации. [16] Таким образом, хотя полный процесс репликации плазмиды Ti не был полностью описан, начальный этап репликации, вероятно, будет зависеть от экспрессии RepC и его связывания с oriV . Следует отметить, что белок RepC действует только в цис-положении , то есть он управляет репликацией только той плазмиды, в которой он закодирован, а не какой-либо другой плазмиды, также присутствующей в бактериальной клетке. [16]
Система разделения плазмиды Ti похожа на систему ParA/ParB, используемую в других плазмидах и бактериальных хромосомах, и, как полагают, действует таким же образом. [17] Мутации в белках RepA или RepB привели к снижению стабильности плазмиды, что указывает на их роль и важность в разделении плазмиды. [4] Способность RepA образовывать нити позволяет ему создавать физический мост, по которому ДНК может быть вытянута к противоположным полюсам делящейся клетки. Между тем, белок RepB может специфически связываться с последовательностью parS , образуя комплекс с ДНК, который может быть распознан RepA. [1] [4] Эта система особенно важна для правильного разделения плазмиды Ti, поскольку плазмида присутствует только в нескольких копиях в бактериальной клетке.
Плазмида Ti поддерживается на низком уровне копийности в бактериальной клетке. Это частично достигается путем влияния на экспрессию инициатора репликации RepC. [1] При связывании с АДФ , RepA активируется для работы с RepB, действуя как отрицательный регулятор кассеты repABC . [3] Таким образом, уровень RepC поддерживается на низком уровне в клетке, предотвращая слишком много раундов репликации во время каждого цикла деления клетки. Кроме того, существует небольшая РНК, известная как RepE, закодированная между repB и repC , которая снижает экспрессию repC . [18] RepE комплементарен RepC и будет связываться с мРНК repC, образуя двухцепочечную молекулу. Это может затем блокировать трансляционное производство белка RepC. [18]
Отдельно, экспрессия кассеты repABC и, следовательно, количество копий плазмиды Ti также зависит от системы кворумного сенсора в Agrobacterium . [4] Системы кворумного сенсора реагируют на плотность популяции бактерий, распознавая молекулу, известную как аутоиндуктор, которая вырабатывается бактериальными клетками на низких уровнях и будет накапливаться до порогового уровня при высокой плотности бактерий. [18] В этом случае аутоиндуктором является молекула N-3-оксооктаноил-L-гомосерин лактона (3-OC 8 -AHL), которая распознается регулятором, известным как TraR. [4] При активации TraR будет связываться с областями, известными как tra- боксы, в промоторных областях генной кассеты repABC , чтобы управлять экспрессией. Таким образом, высокий уровень плотности популяции увеличивает количество плазмид, присутствующих в каждой бактериальной клетке, что, вероятно, поддерживает патогенез в растении-хозяине. [4]

Экспрессия vir -региона обычно подавляется в нормальных условиях и активируется только тогда, когда бактерии воспринимают сигналы, полученные от растений из мест ран. Эта активация необходима для производства белков Vir и переноса ДНК и белков в клетки растения-хозяина. [1]
VirA и VirG образуют двухкомпонентную регуляторную систему в Agrobacterium . [19] Это тип сенсорной и сигнальной системы, обычно встречающийся у бактерий; в этом случае они действуют, чтобы распознавать сигналы, полученные от растений, чтобы управлять экспрессией области vir . Во время распознания VirA, сенсорная киназа гистидина, фосфорилируется перед передачей этой фосфатной группы регулятору ответа VirG. [20] Активированный регулятор ответа VirG затем может связываться с областью ДНК, известной как vir -бокс, расположенной выше каждого промотора vir , чтобы активировать экспрессию области vir . [1] [19] Одной из возможных нисходящих функций распознания, опосредованного VirA и VirG, является направленное движение, или хемотаксис , бактерий в направлении сигналов, полученных от растений; это позволяет Agrobacterium двигаться к месту раны в растениях. [21] Кроме того, при индукции региона vir перенос Т-ДНК может опосредоваться белками Vir. [22]
Оперон virB является крупнейшим опероном в регионе vir , кодирующим 11 белков VirB, участвующих в процессе переноса Т-ДНК и бактериальных белков в клетки растения-хозяина (см. аппарат переноса ниже). [23] [24]
Оперон virC кодирует два белка: VirC1 и VirC2. Эти белки влияют на патогенез Agrobacterium по отношению к различным растениям-хозяевам, и мутации могут снизить, но не устранить вирулентность бактерий. [25] Оба оперона virC и virD могут быть подавлены хромосомно-кодируемым белком, известным как Ros. [26] [27] Ros связывается с областью ДНК, которая перекрывается с сайтом связывания регулятора VirG, и поэтому конкурирует с VirG за контроль их уровней экспрессии. [26] [27] Функционально VirC1 и VirC2 способствуют сборке комплекса релаксосом во время конъюгативного переноса Т-ДНК от бактерий к клетке растения-хозяина. [28] Это энергозависимый процесс, опосредованный их активностью NTPase, и происходит, когда они связываются с областью ДНК, известной как overdrive . [28] В результате они действуют, увеличивая количество производимых цепей Т-ДНК. После производства цепи ДНК, которая должна быть перенесена (цепь переноса, Т-цепь), белки VirC также могут помочь направить цепь переноса в аппарат переноса. [28]
Оперон virD кодирует 4 белка: VirD1-D4. [29] VirD1 и VirD2 участвуют в обработке T-ДНК во время конъюгации для получения T-цепи; это одноцепочечная молекула ДНК, которая транспортируется в клетку растения-хозяина (см. аппарат переноса ниже). [30] Во время обработки VirD1 будет действовать как топоизомераза для раскручивания цепей ДНК. [30] VirD2, релаксаза , затем надрежет одну из цепей ДНК и останется связанной с ДНК, пока она переносится в клетку-реципиент. [31] [32] Внутри клетки-реципиента VirD2 также будет работать вместе с VirE2, чтобы направить перенесенную ДНК в ядро клетки-реципиента. Есть предположения, что VirD2 может фосфорилироваться и дефосфорилироваться различными белками, что влияет на его способность доставлять ДНК. [33] Напротив, о VirD3 известно немного, и мутационные анализы не предоставили никаких подтверждений его роли в вирулентности Agrobacterium . [34] Наконец, VirD4 является важнейшей частью процесса конъюгации, выступая в качестве фактора связи, который распознает и переносит Т-цепь в транспортный канал. [35]
Оперон virE кодирует 2 белка: VirE1 и VirE2. [36] VirE2 — это эффекторный белок, транслоцируемый вместе с T-цепью в клетки растения-хозяина. Там он связывается с T-цепью, чтобы направить его доставку в ядро клетки растения-хозяина. [37] [38] Часть этой активности включает наличие последовательностей ядерной локализации внутри белка, которые маркируют белок и связанную с ним ДНК для входа в ядро. Он также защищает T-цепь от атаки нуклеазы . [39] Существуют некоторые предположения относительно роли VirE2 как белкового канала, позволяющего ДНК перемещаться через цитоплазматическую мембрану растения . [40] С другой стороны, VirE1 может участвовать в содействии переносу белка VirE2 в клетку растения-хозяина. [41] Он связывается с доменом связывания одноцепочечной ДНК VirE2, тем самым предотвращая преждевременное связывание белка VirE2 с Т-цепью внутри бактериальной клетки. [42]
virF — это фактор специфичности хозяина, обнаруженный в некоторых, но не во всех типах плазмид Ti; например, плазмиды Ti октопинового типа обладают virF, а плазмиды нопалинового типа — нет. [43] [44] Способность A. tumefaciens вызывать корончатые галлы у определенных видов растений, но не у других, объясняется наличием или отсутствием этого гена virF . [43] [44]
Оперон virH кодирует 2 белка: VirH1 и VirH2. [45] Биоинформатическое исследование аминокислотных последовательностей белка VirH показало сходство между ними и суперсемейством белков, известных как ферменты цитохрома P450 . [46] Затем было обнаружено, что VirH2 метаболизирует определенные фенольные соединения, обнаруженные VirA. [ 45]
Длина T-ДНК Agrobacterium составляет приблизительно 15-20 кб и будет интегрирована в геном растения-хозяина при его переносе посредством процесса, известного как рекомбинация . Этот процесс использует уже существующие пробелы в геноме клетки растения-хозяина, чтобы позволить T-ДНК спариваться с короткими последовательностями в геноме, запуская процесс лигирования ДНК , где T-ДНК постоянно соединена с геномом растения. [37] Область T-ДНК фланкирована с обоих концов последовательностями из 24 пб.
В геноме клетки растения-хозяина T-ДНК Agrobacterium экспрессируется для производства двух основных групп белков. [1] Одна группа отвечает за производство гормонов роста растений. По мере производства этих гормонов будет происходить увеличение скорости деления клеток и, следовательно, образование опухолей корончатого галла. [47] Вторая группа белков отвечает за управление синтезом опинов в клетках растения-хозяина. Конкретные производимые опины зависят от типа плазмиды Ti, но не от растения-хозяина. Эти опины не могут быть использованы растением-хозяином, а вместо этого будут экспортированы из клетки растения, где они могут быть поглощены клетками Agrobacterium . Бактерии обладают генами в других областях плазмиды Ti, что позволяет катаболизировать опины. [1]
Аппараты переноса, закодированные в плазмиде Ti, должны достигать двух целей: обеспечивать конъюгативный перенос плазмиды Ti между бактериями и обеспечивать доставку T-ДНК и определенных эффекторных белков в клетки растения-хозяина. Это достигается с помощью системы Tra/Trb и системы VirB/VirD4 соответственно, которые являются членами системы секреции типа IV (T4SS). [47]
Для того, чтобы плазмида Ti и T-ДНК были переданы посредством конъюгации, они должны быть сначала обработаны различными белками, такими как фермент релаксаза (TraA/VirD2) и белки переноса и репликации ДНК (Dtr). Вместе эти белки будут распознавать и связываться с областью, известной как начало переноса ( oriT ) в плазмиде Ti, чтобы сформировать комплекс релаксосомы. Для T-ДНК будет создан надрез на пограничной последовательности T-ДНК, и надрезанная T-цепь будет транспортирована к клеточной мембране, где присутствует остальная часть механизма переноса. [31]
В системе VirB/VirD4 релаксаза VirD2 поддерживается дополнительными факторами VirD1, VirC1 и VirC2 при обработке субстрата ДНК. [48] Кроме того, релаксаза VirD2 и белки VirC будут способствовать доставке цепи ДНК к рецептору VirD4 на клеточной мембране. [28] Этот рецептор является важным компонентом T4SS и, как полагают, активирует и опосредует перенос ДНК в канал транслокации между двумя клетками. [49] В таблице ниже обобщены белки, кодируемые в опероне virB , который составляет канал транслокации системы VirB/VirD4. [1]
Способность Agrobacterium доставлять ДНК в растительные клетки открыла новые двери для генной инженерии растений , что позволило производить генетически модифицированные растения (трансгенные растения). [57] Белки, участвующие в опосредовании переноса T-ДНК, сначала распознают пограничные последовательности области T-ДНК. Поэтому ученые могут использовать пограничные последовательности T-ДНК для фланкирования любой желаемой интересующей последовательности — такой продукт затем может быть вставлен в плазмиду и введен в клетки Agrobacterium . [58] Там пограничные последовательности будут распознаны переносящим аппаратом A. tumefaciens и доставлены стандартным образом в целевую растительную клетку. [1] Более того, оставляя только пограничные последовательности T-ДНК, полученный продукт будет редактировать геном растения, не вызывая никаких опухолей у растений. [59] Этот метод использовался для модификации нескольких сельскохозяйственных культур, включая рис, [60] ячмень [61] и пшеницу. [62] Дальнейшие исследования с тех пор расширили целевые объекты A. tumefaciens , включив в них грибы и линии клеток человека. [63] [64]