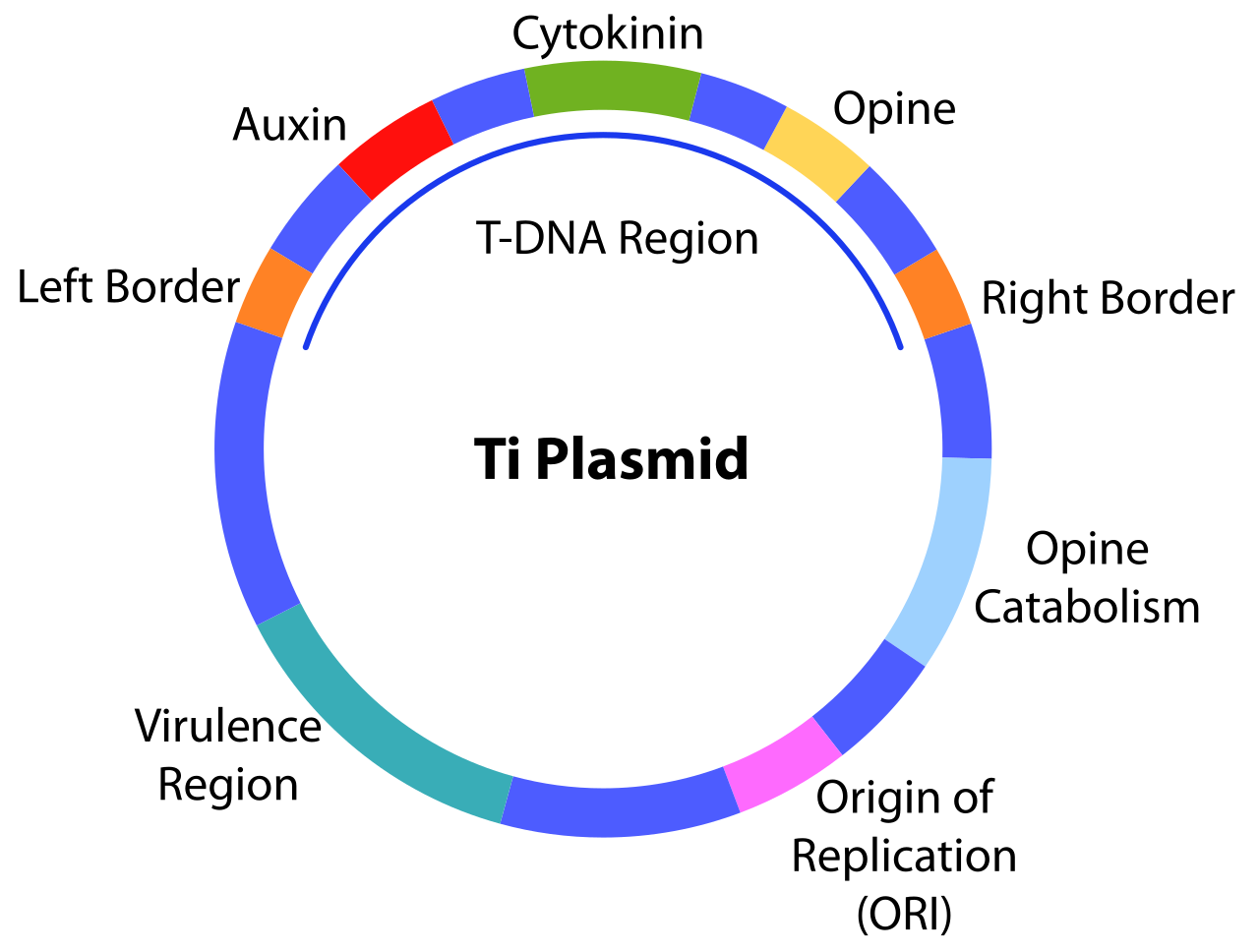
Транспортная ДНК (сокращенно Т-ДНК ) — это перенесенная ДНК плазмиды , вызывающей опухоли (Ti) , некоторых видов бактерий, таких как Agrobacterium tumefaciens и Agrobacterium rhizogenes (фактически плазмида Ri) . Т-ДНК переносится из бактерии в ядерный геном ДНК растения-хозяина . [1] Возможности этой специализированной плазмиды, вызывающей опухоли (Ti), обусловлены двумя основными областями, необходимыми для переноса ДНК в клетку-хозяина. Т-ДНК ограничена повторами из 25 пар оснований на каждом конце. Перенос начинается на правой границе и заканчивается на левой границе и требует генов vir плазмиды Ti.
Бактериальная Т-ДНК имеет длину около 24 000 пар оснований [2] [3] и содержит экспрессируемые в растениях гены , которые кодируют ферменты , синтезирующие опины и фитогормоны . Перенося Т-ДНК в геном растения, бактерия по сути перепрограммирует растительные клетки на рост в опухоль и производство уникального источника пищи для бактерий. Синтез растительных гормонов ауксина и цитокинина ферментами, закодированными в Т-ДНК, позволяет растительной клетке разрастаться, образуя таким образом опухоли корончатого галла, обычно вызываемые инфекцией Agrobacterium tumefaciens . [4] Agrobacterium rhizogenes вызывает похожую инфекцию, известную как болезнь волосатых корней . Опины представляют собой производные аминокислот , используемые бактерией в качестве источника углерода и энергии. Этот естественный процесс горизонтального переноса генов в растениях используется в качестве инструмента для фундаментальных и прикладных исследований в области биологии растений посредством опосредованной Agrobacterium tumefaciens чужеродной генной трансформации и инсерционного мутагенеза. [5] [6] Геномы растений могут быть сконструированы с использованием Agrobacterium для доставки последовательностей, размещенных в бинарных векторах T-ДНК .
Процесс инфицирования Т-ДНК в клетку-хозяина и интеграция в ее ядро включает несколько этапов. Сначала бактерии размножаются в соке раны до инфицирования, а затем прикрепляются к стенкам растительных клеток. Экспрессия генов бактериальной вирулентности приблизительно 10 оперонов активируется восприятием фенольных соединений, таких как ацетосирингон, выделяемых раненой растительной тканью, и следует за контактом между клетками. Затем этот процесс продолжается с макромолекулярной транслокацией из Agrobacterium в цитоплазму клетки-хозяина, передачей Т-ДНК вместе с ассоциированными белками (называемой Т-комплексом ) в ядро клетки-хозяина с последующей разборкой Т-комплекса, стабильной интеграцией Т-ДНК в геном растения-хозяина и окончательной экспрессией перенесенных генов . Интеграция Т-ДНК в геном хозяина включает образование одноцепочечного разрыва в ДНК на правой границе плазмиды Ti. Этот надрез создает область одноцепочечной ДНК от левой границы гена T-ДНК до правой границы, которая была разрезана. Затем одноцепочечные связывающие белки прикрепляются к одноцепочечной ДНК. Синтез ДНК вытесняет одноцепочечную область, а затем второй надрез в области левой границы высвобождает одноцепочечный фрагмент T-ДНК. Далее этот фрагмент может быть включен в геном хозяина. [7]
Известно, что Agrobacterium развивает систему контроля, которая использует факторы растения-хозяина и клеточные процессы для нескольких путей защитного ответа хозяин-растение для вторжения в ядро клетки-хозяина. Для интеграции T-ДНК в целевой геном хозяина Agrobacterium осуществляет множественные взаимодействия с факторами хозяин-растение. [7] Для взаимодействия с белками растения-хозяина многие белки вирулентности Agrobacterium кодируются генами vir. Экспрессия гена vir Agrobacterium происходит через сенсор VirA-VirG, что приводит к образованию мобильной одноцепочечной копии T-ДНК (T-цепи). Обработанная форма VirB2 является основным компонентом T-комплекса, который необходим для трансформации. VirD2 является белком, который закрывает 5'-конец перенесенной T-цепи ковалентным присоединением и транспортируется в цитоплазму клетки-хозяина. [8] [9] VirE2 — это одноцепочечный ДНК-связывающий белок, который, предположительно, покрывает Т-цепь в цитоплазме хозяина путем кооперативного связывания . Затем он направляется в ядро посредством взаимодействия с белками клетки хозяина, такими как импортин a, бактериальный VirE3 и динеин-подобные белки. Несколько других бактериальных эффекторов вирулентности, таких как VirB5, VirB7 (второстепенные компоненты Т-комплекса), VirD5, VirE2, VirE3 и VirF, также могут взаимодействовать с белками клеток растения-хозяина. [10]
Agrobacterium -опосредованный перенос T-ДНК широко используется в качестве инструмента в биотехнологии . На протяжении более двух десятилетий Agrobacterium tumefaciens использовался для введения генов в растения для фундаментальных исследований, а также для коммерческого производства трансгенных культур . [11] В генной инженерии гены, способствующие образованию опухолей и синтезу опинов, удаляются из T-ДНК и заменяются интересующим геном и/или маркером отбора, который требуется для установления того, какие растения были успешно трансформированы. Примерами маркеров отбора являются неомицинфосфотрансфераза, гигромицин Bфосфотрансфераза (которые оба фосфорилируют антибиотики) и фосфинотрицинацетилтрансфераза (которая ацетилирует и дезактивирует фосфинотрицин , мощный ингибитор глутаминсинтетазы ) или гербицидные составы, такие как Basta или Bialophos. [12] Другая система отбора, которая может быть использована, — это использование метаболических маркеров, таких как фосфоманнозоизомераза. [13] Затем Agrobacterium используется в качестве вектора для переноса сконструированной T-ДНК в растительные клетки, где она интегрируется в растительный геном. Этот метод может быть использован для создания трансгенных растений, несущих чужеродный ген. Agrobacterium tumefaciens способен эффективно переносить чужеродную ДНК как в однодольные , так и в двудольные растения, при этом учитывая критически важные факторы, такие как генотип растений, типы и возраст инокулированных тканей, вид векторов, штаммы Agrobacterium , гены маркеров селекции и селективные агенты, а также различные условия культивирования тканей. [4]
Та же процедура переноса T-ДНК может быть использована для разрушения генов посредством инсерционного мутагенеза . [6] Вставленная последовательность T-ДНК не только создает мутацию, но и ее вставка также «маркирует» [14] затронутый ген, что позволяет изолировать его в качестве фланкирующих последовательностей T-ДНК. Репортерный ген может быть связан с правым концом T-ДНК для трансформации вместе с плазмидным репликоном и селективным геном устойчивости к антибиотику (например, гигромицину ), и может проявлять приблизительно 30% средней эффективности, имея успешные инсерции T-ДНК, индуцирующие слияния генов в Arabidopsis thaliana . [15]
Обратная генетика включает в себя тестирование предполагаемой функции гена, который известен, путем его разрушения, а затем поиска эффекта этой индуцированной мутации на фенотип организма. Мутагенез с маркировкой T-ДНК включает в себя скрининг популяций с помощью инсерционных мутаций T-ДНК. Коллекции известных мутаций T-ДНК предоставляют ресурсы для изучения функций отдельных генов, как это было разработано для модельного растения Arabidopsis thaliana . [16] [17] Примеры инсерционных мутаций T-ДНК в Arabidopsis thaliana включают те, которые связаны со многими классами фенотипов, включая летальные для сеянцев, вариации размера, вариации пигмента, дефектные эмбрионы, растения с пониженной фертильностью и морфологически или физиологически аберрантные растения. [18]