
.gif/1280px-Illustration_of_the_Caspar-Klug_model_for_viruses_(or_%22Goldberg_Polyhedra%22_or_%22Geodesic_domes%22_or_%22Fullerenes%22).gif)
Капсид – это белковая оболочка вируса , содержащая его генетический материал . Он состоит из нескольких олигомерных (повторяющихся) структурных субъединиц, состоящих из белка, называемых протомерами . Наблюдаемые трехмерные морфологические субъединицы, которые могут соответствовать или не соответствовать отдельным белкам, называются капсомерами . Белки, составляющие капсид, называются капсидными белками или белками вирусной оболочки ( VCP ). Капсид и внутренний геном называются нуклеокапсидом .
Капсиды широко классифицируются в зависимости от их структуры. Большинство вирусов имеют капсиды со спиральной или икосаэдрической [2] [3] структурой. Некоторые вирусы, такие как бактериофаги , имеют более сложную структуру из-за ограничений эластичности и электростатики. [4] Икосаэдрическая форма, имеющая 20 равносторонних треугольных граней, приближается к сфере , а спиральная форма напоминает форму пружины , занимая пространство цилиндра, но не являясь самим цилиндром. [5] Лица капсида могут состоять из одного или нескольких белков. Например, капсид вируса ящура имеет грани, состоящие из трех белков, называемых VP1–3. [6]
Некоторые вирусы имеют оболочку , то есть капсид покрыт липидной мембраной, известной как вирусная оболочка . Оболочка приобретается капсидом из внутриклеточной мембраны хозяина вируса; примеры включают внутреннюю ядерную мембрану, мембрану Гольджи и внешнюю мембрану клетки . [7]
Как только вирус заразил клетку и начал размножаться, новые субъединицы капсида синтезируются с использованием клеточного механизма биосинтеза белка . У некоторых вирусов, в том числе со спиральными капсидами и особенно с РНК-геномами, капсидные белки собираются вместе с их геномами. У других вирусов, особенно у более сложных вирусов с геномами двухцепочечной ДНК, капсидные белки собираются в пустые прокапсиды- предшественники , которые включают специализированную портальную структуру в одной вершине. Через этот портал вирусная ДНК транслоцируется в капсид. [8]
Структурный анализ архитектуры главного капсидного белка (MCP) использовался для классификации вирусов по линиям. Например, бактериофаг PRD1, вирус водорослей Paramecium bursaria Chlorella вирус-1 (PBCV-1), мимивирус и аденовирус млекопитающих отнесены к одной и той же линии, тогда как бактериофаги с двухцепочечной ДНК ( Caudovirales ) и герпесвирус принадлежат к одной и той же линии. вторая родословная. [9] [10] [11] [12]

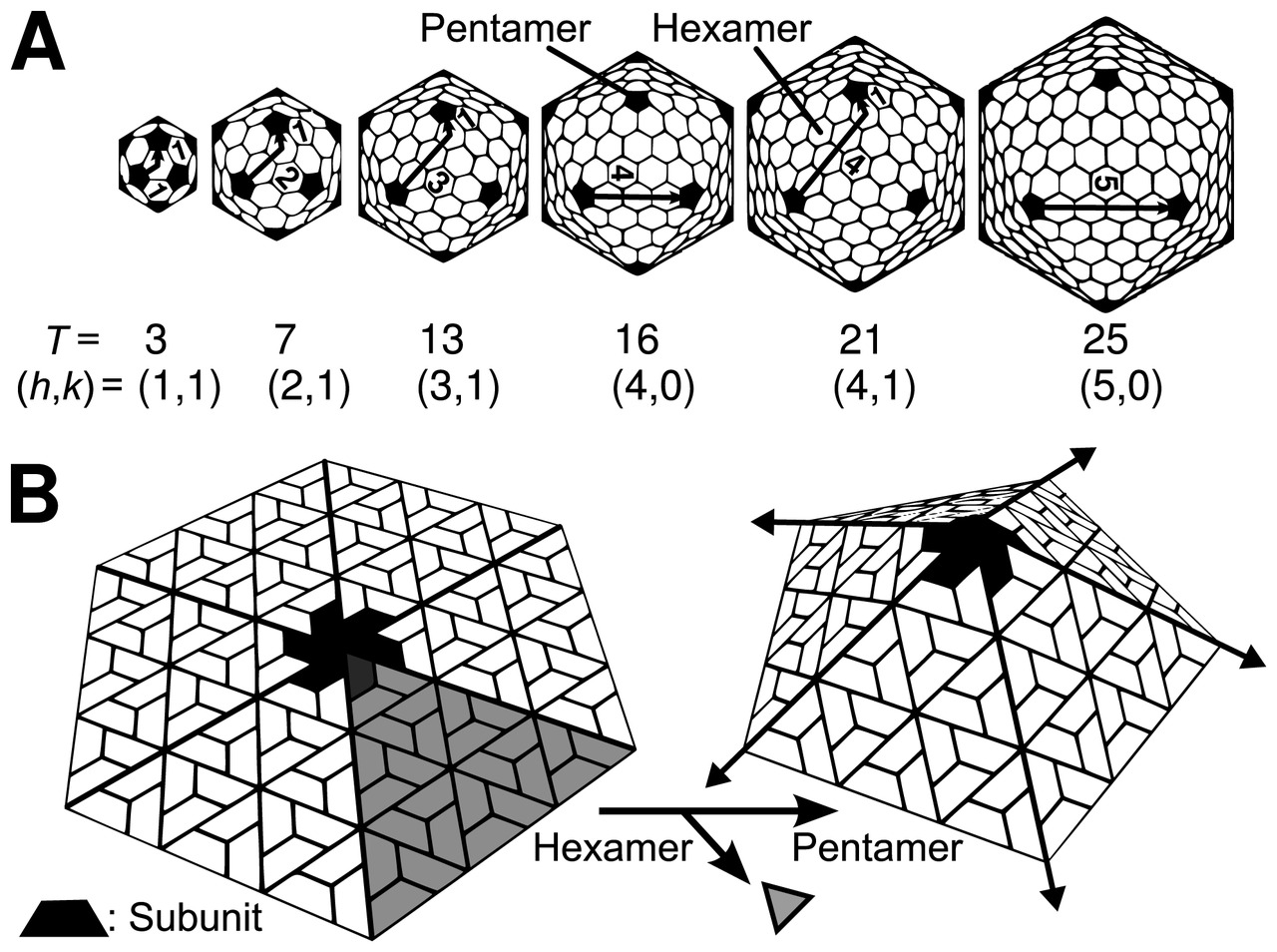
Икосаэдрическая структура чрезвычайно распространена среди вирусов. Икосаэдр состоит из 20 треугольных граней , ограниченных 12 пятикратными вершинами, и состоит из 60 асимметричных звеньев. Таким образом, икосаэдрический вирус состоит из белковых субъединиц 60N. Число и расположение капсомеров в икосаэдрическом капсиде можно классифицировать с помощью «принципа квазиэквивалентности», предложенного Дональдом Каспаром и Аароном Клугом . [13] Как и многогранники Гольдберга , икосаэдрическую структуру можно рассматривать как построенную из пентамеров и гексамеров. Структуры могут быть проиндексированы двумя целыми числами h и k , с и ; структуру можно представить себе так, что она делает h шагов от края пентамера, поворачивает на 60 градусов против часовой стрелки, а затем делает k шагов, чтобы добраться до следующего пентамера. Число триангуляции T капсида определяется как:
В этой схеме икосаэдрические капсиды содержат 12 пентамеров плюс 10 ( Т - 1) гексамеров. [14] [15] Т - число характеризует размер и сложность капсидов. [16] Геометрические примеры для многих значений h , k и T можно найти в списке геодезических многогранников и многогранников Гольдберга .
Существует множество исключений из этого правила: например, полиомавирусы и папилломавирусы имеют пентамеры вместо гексамеров в шестивалентных положениях в квази-решетке T = 7. Члены линии вируса с двухцепочечной РНК, включая реовирус , ротавирус и бактериофаг φ6, имеют капсиды, построенные из 120 копий капсидного белка, что соответствует капсиду T = 2 или, возможно, капсиду T = 1 с димером в асимметричной единице. Аналогичным образом, многие небольшие вирусы имеют псевдокапсид T = 3 (или P = 3), который организован в соответствии с решеткой T = 3, но с отдельными полипептидами, занимающими три квазиэквивалентных положения [17].
Т-числа могут быть представлены по-разному, например Т = 1 можно представить только в виде икосаэдра или додекаэдра и, в зависимости от типа квазисимметрии, Т = 3 можно представить в виде усеченного додекаэдра , икосододекаэдра , или усеченный икосаэдр и их соответствующие двойники: триакисикосаэдр , ромбический триаконтаэдр или пентакисдодекаэдр . [18] [ нужны разъяснения ]

Вытянутый икосаэдр — обычная форма головок бактериофагов. Такая конструкция представляет собой цилиндр с крышками на обоих концах. Цилиндр состоит из 10 вытянутых треугольных граней. Число Q (или Tmid ) , которое может быть любым положительным целым числом, [19] указывает количество треугольников, состоящих из асимметричных субъединиц, которые составляют 10 треугольников цилиндра. Колпачки классифицируются по номеру T (или T end ). [20]
Бактерия E. coli является хозяином бактериофага Т4 , имеющего вытянутую головку. Кодируемый бактериофагом белок gp31 функционально гомологичен белку- шаперону GroES E. coli и способен замещать его при сборке вирионов бактериофага Т4 во время инфекции. [21] Как и GroES, gp31 образует стабильный комплекс с шаперонином GroEL , который абсолютно необходим для сворачивания и сборки in vivo основного капсидного белка gp23 бактериофага Т4. [21]

Многие палочковидные и нитевидные вирусы растений имеют капсиды со спиральной симметрией . [22] Спиральную структуру можно описать как набор n 1-D молекулярных спиралей, связанных n -кратной осевой симметрией. [23] Спиральные преобразования делятся на две категории: одномерные и двумерные спиральные системы. [23] Создание целой спиральной структуры опирается на набор трансляционных и вращательных матриц, которые закодированы в банке данных белков. [23] Спиральная симметрия задается формулой P = μ x ρ , где μ — количество структурных единиц на виток спирали, ρ — осевой подъем на единицу, а P — шаг спирали. Структура называется открытой из-за того, что любой объем можно замкнуть, изменяя длину спирали. [24] Наиболее изученным спиральным вирусом является вирус табачной мозаики. [22] Вирус представляет собой одну молекулу (+) цепи РНК. Каждый белок оболочки внутри спирали связывает три нуклеотида генома РНК. Вирусы гриппа А отличаются тем, что содержат несколько рибонуклеопротеинов, вирусный белок NP организует РНК в спиральную структуру. Размер также отличается; вирус табачной мозаики имеет 16,33 белковых субъединиц на виток спирали [22] , тогда как вирус гриппа А имеет хвостовую петлю из 28 аминокислот. [25]
Функции капсида заключаются в следующем:
Вирус должен собрать стабильную защитную белковую оболочку, чтобы защитить геном от смертельных химических и физических агентов. К ним относятся экстремальные значения pH или температуры, а также протеолитические и нуклеолитические ферменты . У вирусов без оболочки капсид сам по себе может участвовать во взаимодействии с рецепторами клетки-хозяина, что приводит к проникновению через мембрану клетки-хозяина и интернализации капсида. Доставка генома происходит путем последующего снятия оболочки или разборки капсида и высвобождения генома в цитоплазму или путем выброса генома через специализированную портальную структуру непосредственно в ядро клетки-хозяина.
Было высказано предположение, что многие вирусные капсидные белки неоднократно эволюционировали из функционально разнообразных клеточных белков. [26] Рекрутирование клеточных белков, по-видимому, происходило на разных стадиях эволюции, так что некоторые клеточные белки были захвачены и рефункционализированы до расхождения клеточных организмов в три современные области жизни, тогда как другие были захвачены относительно недавно. В результате одни капсидные белки широко распространены у вирусов, заражающих отдаленно родственные организмы (например, капсидные белки с желеобразной складкой ), тогда как другие ограничены определенной группой вирусов (например, капсидные белки альфавирусов). [26] [27]
Вычислительная модель (2015) показала, что капсиды могли возникнуть раньше вирусов и что они служили средством горизонтального переноса между сообществами репликаторов, поскольку эти сообщества не могли выжить, если количество генных паразитов увеличивалось, а за формирование отвечали определенные гены. этих структур и тех, которые способствовали выживанию самовоспроизводящихся сообществ. [28] Смещение этих предковых генов между клеточными организмами может способствовать появлению новых вирусов в ходе эволюции. [27]


