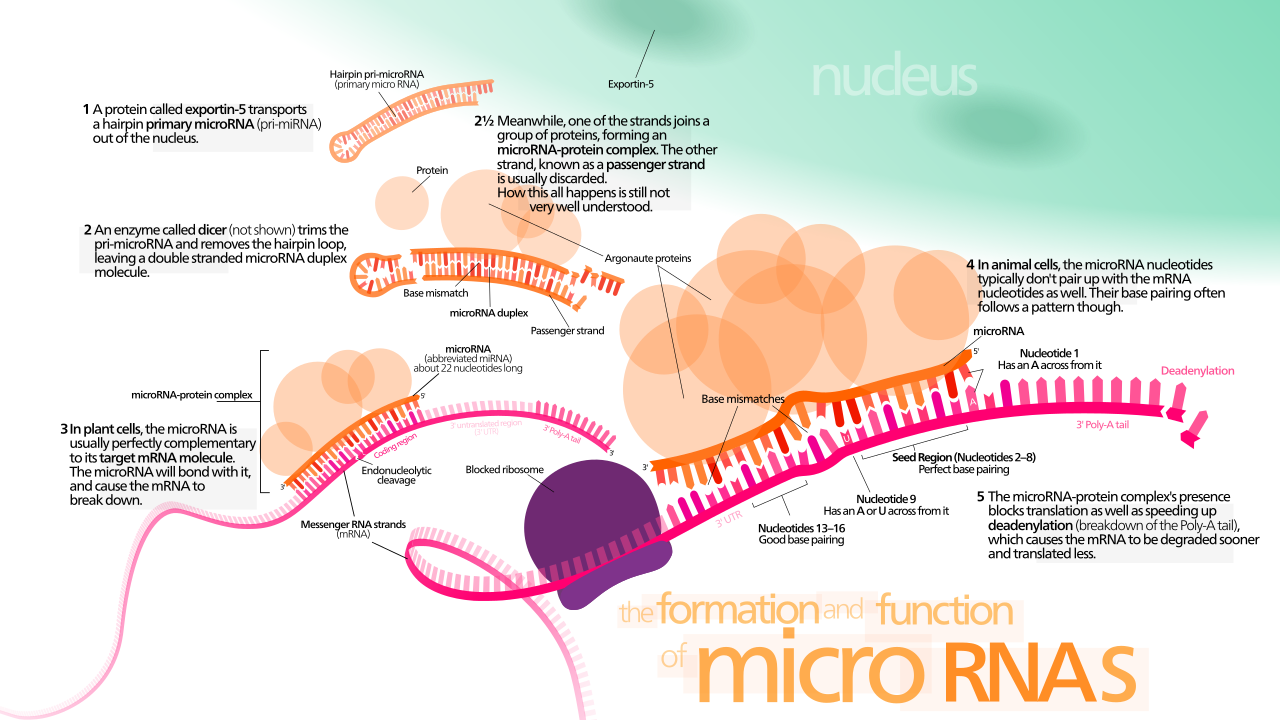
Комплекс подавления генов, индуцированный РНК , или RISC , представляет собой многобелковый комплекс , в частности рибонуклеопротеин , который функционирует в подавлении генов посредством различных путей на транскрипционном и трансляционном уровнях. [1] Используя фрагменты одноцепочечной РНК (ssRNA), такие как микроРНК (miRNA) или двухцепочечные малые интерферирующие РНК (siRNA), комплекс функционирует как ключевой инструмент в регуляции генов. [2] Одноцепочечная РНК действует как шаблон для RISC для распознавания комплементарного транскрипта информационной РНК (мРНК) . После обнаружения один из белков в RISC, Argonaute , активирует и расщепляет мРНК. Этот процесс называется РНК-интерференцией (RNAi) и встречается у многих эукариот ; это ключевой процесс в защите от вирусных инфекций , поскольку он запускается присутствием двухцепочечной РНК (dsRNA). [3] [4] [1]
Хэннон и его коллеги попытались идентифицировать механизмы РНК-интерференции, участвующие в подавлении генов , с помощью dsRNAs в клетках Drosophila . Клетки Drosophila S2 были трансфицированы вектором экспрессии lacZ для количественной оценки экспрессии генов с активностью β-галактозидазы . Их результаты показали, что котрансфекция с lacZ dsRNA значительно снижает активность β-галактозидазы по сравнению с контрольной dsRNA. Следовательно, dsRNAs контролируют экспрессию генов посредством комплементарности последовательностей .
Затем клетки S2 были трансфицированы dsRNA циклина E дрозофилы . Циклин E является важным геном для перехода клеточного цикла в фазу S. dsRNA циклина E останавливает клеточный цикл в фазе G 1 (перед фазой S). Таким образом, РНК-интерференция может воздействовать на эндогенные гены.
Кроме того, dsRNA циклина E только уменьшила РНК циклина E — аналогичный результат был также показан с использованием dsRNA, соответствующей циклину A , которая действует в фазах S, G 2 и M клеточного цикла. Это демонстрирует характерную особенность РНК-интерференции: сниженные уровни мРНК соответствуют уровням добавленной dsRNA.
Чтобы проверить, было ли наблюдаемое ими снижение уровней мРНК результатом прямого воздействия на мРНК (как предполагалось на основании данных из других систем), клетки Drosophila S2 были трансфицированы либо двуцепочечной РНК циклина E дрозофилы, либо двуцепочечной РНК lacZ , а затем инкубированы с синтетическими мРНК для циклина E или lacZ .
Клетки, трансфицированные dsRNA циклина E, показали деградацию только в транскриптах циклина E — транскрипты lacZ были стабильными. Наоборот, клетки, трансфицированные dsRNA lacZ , показали деградацию только в транскриптах lacZ , но не в транскриптах циклина E. Их результаты привели Хэннона и его коллег к предположению, что РНК-интерференция разрушает целевую мРНК посредством «специфической для последовательности нуклеазной активности». Они назвали этот фермент нуклеазы RISC. [5] Позже Девананд Саркар и его коллеги Прасанна К. Сантекадур и Бёнг Квон Ю из Университета Содружества Вирджинии выяснили активность RISC и ее молекулярный механизм в раковых клетках, и они идентифицировали еще один новый компонент RISC, названный AEG-1 [47].
Функция в РНК-интерференции
Домен PIWI белка Argonaute в комплексе с двухцепочечной РНК.
Включение siRNA/miRNA
RNase III Dicer является критическим членом RISC, который инициирует процесс интерференции РНК, производя двухцепочечную siRNA или одноцепочечную miRNA. Ферментативное расщепление dsRNA внутри клетки производит короткие фрагменты siRNA длиной 21-23 нуклеотида с двухнуклеотидным 3'- выступом. [6] [7] Dicer также обрабатывает pre-miRNA, которая образует структуру шпильки, имитирующую dsRNA, аналогичным образом. Фрагменты dsRNA загружаются в RISC, причем каждая нить имеет различную судьбу на основе феномена правила асимметрии, выбора одной нити в качестве направляющей нити по сравнению с другой на основе термодинамической стабильности. [8] [9] [10] [11] Вновь созданные miRNA или siRNA действуют как одноцепочечные направляющие последовательности для RISC, чтобы нацелить мРНК на деградацию. [12] [13]
Цепь с менее термодинамически стабильным 5'-концом выбирается белком Argonaute и интегрируется в RISC. [11] [14] Эта цепь известна как направляющая цепь и нацеливает мРНК на деградацию.
Другая цепь, известная как пассажирская цепь, разрушается RISC. [15]
Часть пути РНК-интерференции с различными способами, которыми RISC может подавлять гены с помощью их информационной РНК.
Регуляция генов
AGO2 (серый) в комплексе с микроРНК (светло-голубой) и ее целевой мРНК (темно-синий)
Основные белки RISC, Ago2, SND1 и AEG-1, играют решающую роль в функции подавления генов комплекса. [16]
RISC использует направляющую цепь miRNA или siRNA для нацеливания на комплементарные 3'-нетранслируемые области (3'UTR) транскриптов мРНК посредством спаривания оснований Уотсона-Крика , что позволяет регулировать экспрессию генов транскрипта мРНК несколькими способами. [17] [1]
деградация мРНК
Наиболее понятная функция RISC — это деградация целевой мРНК, которая снижает уровни транскрипта, доступного для трансляции рибосомами . Эндонуклеолитическое расщепление мРНК, комплементарной направляющей цепи RISC, белком Argonaute является ключом к инициации РНК-интерференции. [18] Для деградации мРНК необходимо два основных условия:
почти идеальное комплементарное соответствие между направляющей цепью и целевой последовательностью мРНК, и,
каталитически активный белок Argonaute, называемый «резчиком», для расщепления целевой мРНК. [1]
Существует два основных пути деградации мРНК после того, как произошло расщепление. Оба инициируются деградацией поли(А)-хвоста мРНК , что приводит к удалению 5'-кэпа мРНК.
Деградация транскрипта от 3' до 5' осуществляется экзосомой и комплексом Ski . [18]
Трансляционная репрессия
RISC может модулировать загрузку рибосомы и вспомогательных факторов в трансляции для подавления экспрессии связанного транскрипта мРНК. Трансляционная репрессия требует только частичного соответствия последовательности между направляющей цепью и целевой мРНК. [1]
Перевод можно регулировать на начальном этапе с помощью:
Было показано, что RITS направляет формирование гетерохроматина в центромерах посредством распознавания центромерных повторов. Благодаря спариванию оснований siRNA (направляющей нити) с целевыми последовательностями хроматина могут быть привлечены ферменты, модифицирующие гистоны. [25]
Механизм не очень хорошо изучен; однако, RITS разрушает зарождающиеся транскрипты мРНК. Было высказано предположение, что этот механизм действует как «самоусиливающаяся петля обратной связи », поскольку зарождающиеся транскрипты используются РНК-зависимой РНК-полимеразой (RdRp) для генерации большего количества siRNA. [26]
В Schizosaccharomyces pombe и Arabidopsis процессинг dsRNA-мишеней в siRNA РНКазами Dicer может инициировать путь подавления генов путем образования гетерохроматина. Белок Argonaute, известный как AGO4, взаимодействует с малыми РНК, которые определяют гетерохроматиновые последовательности. Гистонметилтрансфераза (HMT), H3K9 , метилирует гистон H3 и привлекает хромодоменные белки к сайтам метилирования. Метилирование ДНК поддерживает подавление генов, поскольку гетерохроматиновые последовательности могут быть установлены или распространены. [27]
удаление ДНК
siRNA, генерируемая RISC, по-видимому, играет роль в деградации ДНК во время развития соматического макронуклеуса у инфузорий рода Tetrahymena . Это похоже на эпигенетический контроль образования гетерохроматина и подразумевается как защита от вторжения генетических элементов. [27]
Подобно образованию гетерохроматина в S. pombe и Arabidopsis , белок Tetrahymena, относящийся к семейству Argonaute, Twi1p, катализирует удаление ДНК целевых последовательностей, известных как внутренние последовательности удаления (IES). Используя метилтрансферазы и хромодоменные белки, IES гетерохроматизируются и удаляются из ДНК. [27]
Белки, ассоциированные с RISC
Полная структура RISC до сих пор не решена. Во многих исследованиях сообщалось о диапазоне размеров и компонентов для RISC, но не совсем ясно, связано ли это с тем, что существует несколько комплексов RISC, или с разными источниками, которые используют разные исследования. [28]
Полноразмерный белок-аргонавт из археи вида Pyrococcus furiosus .
Несмотря на это, очевидно, что белки Argonaute присутствуют и необходимы для функционирования. Кроме того, есть понимание некоторых ключевых белков (в дополнение к Argonaute) в комплексе, которые позволяют RISC выполнять свою функцию.
Белки-аргонавты
Белки Argonaute — это семейство белков, обнаруженных у прокариот и эукариот. Их функция у прокариот неизвестна, но у эукариот они отвечают за РНК-интерференцию. [43] У человеческих аргонавтов есть восемь членов семейства, из которых только Argonaute 2 участвует исключительно в целевом расщеплении РНК в RISC. [40]
Комплекс загрузки RISC позволяет загружать фрагменты dsRNA (сгенерированные Dicer) на Argonaute 2 (с помощью TRBP) в рамках пути РНК-интерференции.
RISC-загрузочный комплекс
Комплекс загрузки RISC (RLC) является необходимой структурой, необходимой для загрузки фрагментов dsRNA в RISC для нацеливания на мРНК. RLC состоит из dicer, трансактивирующего белка РНК-связывания ответа ( TRBP ) и Argonaute 2.
Dicer — это эндонуклеаза РНКаза III , которая генерирует фрагменты дцРНК для загрузки, которые направляют РНК-интерференцию.
TRBP — это белок с тремя двухцепочечными РНК-связывающими [белковыми доменами|доменами].
Argonaute 2 — это РНКаза, являющаяся каталитическим центром RISC.
Dicer ассоциируется с TRBP и Argonaute 2, чтобы облегчить перенос фрагментов dsRNA, сгенерированных Dicer, в Argonaute 2. [44] [45]
Более поздние исследования показали, что человеческая РНК-хеликаза А может способствовать RLC. [46]
Другие белки
Недавно идентифицированные члены RISC — SND1 и MTDH . [47] SND1 и MTDH являются онкогенами и регулируют экспрессию различных генов. [48]
Пока неясно, как активированный комплекс RISC находит мишени мРНК в клетке, хотя было показано, что этот процесс может происходить в ситуациях, выходящих за рамки текущей трансляции белка из мРНК. [50]
Эндогенно экспрессируемая miRNA у метазоа обычно не идеально комплементарна большому количеству генов, и поэтому они модулируют экспрессию посредством трансляционной репрессии. [51] [52] Однако у растений этот процесс имеет гораздо большую специфичность к целевой мРНК, и обычно каждая miRNA связывается только с одной мРНК. Большая специфичность означает, что деградация мРНК происходит с большей вероятностью. [53]
^ abcdef Pratt AJ, MacRae IJ (2009). «Комплекс подавления экспрессии РНК: универсальная машина подавления экспрессии генов». Журнал биологической химии . 284 (27): 17897–17901. doi : 10.1074/jbc.R900012200 . PMC 2709356. PMID 19342379 .
^ ab Filipowicz W, Bhattacharyya SN, Sonenber N (2008). «Механизмы посттранскрипционной регуляции микроРНК: есть ли ответы?». Nature Reviews Genetics . 9 (2): 102–114. doi :10.1038/nrg2290. PMID 18197166. S2CID 11824239.
^ ab Fire A, Xu S, Montgomery MK, Kostas SA, Driver SE, Mello CC (1998). «Мощное и специфическое генетическое вмешательство двухцепочечной РНК в Caenorhabditis elegans». Nature . 391 (6669): 806–811. Bibcode :1998Natur.391..806F. doi :10.1038/35888. PMID 9486653. S2CID 4355692.
^ Уотсон, Джеймс Д. (2008). Молекулярная биология гена . Сан-Франциско, Калифорния: Cold Spring Harbor Laboratory Press. С. 641–648. ISBN978-0-8053-9592-1.
^ Zamore PD, Tuschl T, Sharp PA, Bartel DP (2000). «РНК-интерференция: двухцепочечная РНК направляет АТФ-зависимое расщепление мРНК с интервалом от 21 до 23 нуклеотидов». Cell . 101 (1): 25–33. doi : 10.1016/S0092-8674(00)80620-0 . PMID 10778853.
^ Vermeulen A, Behlen L, Reynolds A, Wolfson A, Marshall W, Karpilow J, Khvorova A (2005). «Вклад структуры dsRNA в специфичность и эффективность Dicer». RNA . 11 (5): 674–682. doi :10.1261/rna.7272305. PMC 1370754 . PMID 15811921.
^ Hutvagner, Gyorgy (2005). "Асимметрия малых РНК в РНКi: функция в сборке RISC и регуляции генов". FEBS Letters . 579 (26): 5850–5857. Bibcode :2005FEBSL.579.5850H. doi : 10.1016/j.febslet.2005.08.071 . hdl : 10453/15313 . ISSN 1873-3468. PMID 16199039.
^ Schwarz DS, Hutvágner G, Du T, Xu Z, Aronin N, Zamore PD (2003). «Асимметрия в сборке комплекса ферментов РНК-интерференции». Cell . 115 (2): 199–208. doi : 10.1016/S0092-8674(03)00759-1 . PMID 14567917.
^ Хворова А., Рейнольдс А., Джаясена С.Д. (2003). «Функциональные siRNA и miRNA проявляют смещение нитей». Cell . 115 (2): 209–216. doi : 10.1016/S0092-8674(03)00801-8 . PMID 14567918. S2CID 2500175.
^ ab Siomi H, Siomi MC (2009). «На пути к чтению кода РНК-интерференции». Nature . 457 (7228): 396–404. Bibcode :2009Natur.457..396S. doi :10.1038/nature07754. PMID 19158785. S2CID 205215974.
^ Preall, Jonathan B.; Sontheimer, Erik J. (2005-11-18). «RNAi: RISC Gets Loaded». Cell . 123 (4): 543–545. doi : 10.1016/j.cell.2005.11.006 . ISSN 0092-8674. PMID 16286001.
^ Preall, Jonathan B.; He, Zhengying; Gorra, Jeffrey M.; Sontheimer, Erik J. (2006-03-07). «Выбор короткой интерферирующей цепи РНК не зависит от полярности процессинга дцРНК во время РНКi у дрозофилы». Current Biology . 16 (5): 530–535. Bibcode : 2006CBio...16..530P. doi : 10.1016/j.cub.2006.01.061 . ISSN 0960-9822. PMID 16527750.
^ Gregory RI, Chendrimada TP, Cooch N, Shiekhattar R (2005). "Human RISC couples microRNA biogenesis and posttransscriptional gene silencing". Cell . 123 (4): 631–640. doi : 10.1016/j.cell.2005.10.022 . PMID 16271387.
^ Santhekadur, Prasanna K.; Kumar, Divya P. (2020-06-01). «Сборка RISC и посттранскрипционная регуляция генов при гепатоцеллюлярной карциноме». Genes & Diseases . 7 (2): 199–204. doi : 10.1016/j.gendis.2019.09.009 . ISSN 2352-3042. PMC 7083748 . PMID 32215289.
^ ab Wakiyama M, Takimoto K, Ohara O, Yokoyama S (2007). «Деаденилирование мРНК, опосредованное микроРНК Let-7, и трансляционная репрессия в бесклеточной системе млекопитающих». Genes & Development . 21 (15): 1857–1862. doi :10.1101/gad.1566707. PMC 1935024. PMID 17671087 .
^ ab ORBAN, TAMAS I.; IZAURRALDE, ELISA (апрель 2005 г.). «Распад мРНК, нацеленный на RISC, требует XRN1, комплекса Ski и экзосомы». РНК . 11 (4): 459–469. doi :10.1261/rna.7231505. ISSN 1355-8382. PMC 1370735 . PMID 15703439.
^ Сен GL, Блау HM (2005). «Argonaute2/RISC находится в местах распада мРНК млекопитающих, известных как цитоплазматические тельца». Nature Cell Biology . 7 (6): 633–636. doi :10.1038/ncb1265. PMID 15908945. S2CID 6085169.
^ Verdel A, Jia S, Gerber S, Sugiyama T, Gygi S, Grewal SI, Moazed D (2004). «RITS действует в цис-положении, способствуя транскрипции, опосредованной РНК-интерференцией, и посттранскрипционному подавлению». Nature Genetics . 36 (11): 1174–1180. doi : 10.1038/ng1452 . PMID 15475954.
^ Шимада, Юкико; Мон, Фабио; Бюлер, Марк (2016-12-01). «Транскрипционный комплекс подавления, индуцированный РНК, воздействует на хроматин исключительно посредством взаимодействия с формирующимися транскриптами». Гены и развитие . 30 (23): 2571–2580. doi :10.1101/gad.292599.116. ISSN 0890-9369. PMC 5204350. PMID 27941123 .
^ Sugiyama T, Cam H, Verdel A, Moazed D, Grewal SI (2005). «РНК-зависимая РНК-полимераза является важным компонентом самоподдерживающейся петли, связывающей сборку гетерохроматина с продукцией siRNA». Труды Национальной академии наук Соединенных Штатов Америки . 102 (1): 152–157. Bibcode : 2005PNAS..102..152S. doi : 10.1073/pnas.0407641102 . PMC 544066. PMID 15615848 .
^ abc Mochizuki K, Gorovsky MA (2004). "Малые РНК в расположении генома у Tetrahymena ". Current Opinion in Genetics & Development . 14 (2): 181–187. doi :10.1016/j.gde.2004.01.004. PMID 15196465.
^ abc Sontheimer EJ (2005). «Сборка и функция комплексов подавления РНК». Nature Reviews Molecular Cell Biology . 6 (2): 127–138. doi :10.1038/nrm1568. PMID 15654322. S2CID 27294007.
^ ab Liu Q, Rand TA, Kalidas S, Du F, Kim HE, Smith DP, Wang X (2003). "R2D2, мост между этапами инициации и эффекторными этапами пути РНК- интерференции дрозофилы ". Science . 301 (5641): 1921–1925. Bibcode :2003Sci...301.1921L. doi :10.1126/science.1088710. PMID 14512631. S2CID 41436233.
^ abcdefghij Pham JW, Pellio JL, Lee YS, Carthew RW, Sontheimer EJ (2004). «Комплекс 80S, зависимый от Dicer-2, расщепляет целевые мРНК во время РНК-интерференции у дрозофилы». Cell . 117 (1): 83–94. doi : 10.1016/S0092-8674(04)00258-2 . PMID 15066284.
^ abcd Tomari Y, Du T, Haley B, Schwarz DS, Bennett R, Cook HA, Koppetsch BS, Theurkauf WE, Zamore PD (2004). «Дефекты сборки RISC в мутантном armitage RNAi Drosophila». Cell . 116 (6): 831–841. doi : 10.1016/S0092-8674(04)00218-1 . PMID 15035985.
^ abc Hammond SM, Boettcher S, Caudy AA, Kobayashi R, Hannon GJ (2001). "Argonaute2, связь между генетическим и биохимическим анализом РНКi". Science . 293 (5532): 1146–1150. doi :10.1126/science.1064023. PMID 11498593. S2CID 5271290.
^ abc Caudy AA, Myers M, Hannon GJ, Hammond SM (2002). «Fragile X-related protein and VIG associated with the RNA interference machine». Genes & Development . 16 (19): 2491–2496. doi :10.1101/gad.1025202. PMC 187452. PMID 12368260 .
^ ab Rand TA, Ginalski K, Grishin NV, Wang X (2004). "Биохимическая идентификация Argonaute 2 как единственного белка, необходимого для активности комплекса подавления, индуцированного РНК". Труды Национальной академии наук Соединенных Штатов Америки . 101 (40): 14385–14389. Bibcode :2004PNAS..10114385R. doi : 10.1073/pnas.0405913101 . PMC 521941 . PMID 15452342.
^ abcde Ishizuka A, Siomi MC, Siomi H (2002). «Фрагментарный белок X дрозофилы взаимодействует с компонентами РНК-интерференции и рибосомальными белками». Genes & Development . 16 (19): 2497–2508. doi :10.1101/gad.1022002. PMC 187455. PMID 12368261 .
^ abc Martinez J, Patkaniowska A, Urlaub H, Luhrmann R, Tuschl T (2002). «Одноцепочечные антисмысловые siRNA направляют расщепление целевой РНК в РНКi». Cell . 110 (5): 563–574. doi :10.1016/S0092-8674(02)00908-X. hdl : 11858/00-001M-0000-0012-F2FD-2 . PMID 12230974. S2CID 10616773.
^ ab Liu J, Carmell MA, Rivas FV, Marsden CG, Thomson JM, Song JJ, Hammond SM, Joshua-Tor L, Hannon GJ (2004). «Argonaute2 — каталитический двигатель РНК-интерференции млекопитающих». Science . 305 (5689): 1437–1441. Bibcode :2004Sci...305.1437L. doi : 10.1126/science.1102513 . PMID 15284456. S2CID 2778088.
^ Мартинес Дж., Тушл Т. (2004). «RISC — это РНК-эндонуклеаза, продуцирующая 5′ фосфомоноэстер». Гены и развитие . 18 (9): 975–980. doi :10.1101/gad.1187904. PMC 406288. PMID 15105377 .
^ abc Мейстер Г., Ландталер М., Патканёвска А., Дорсетт Ю., Тенг Г., Тушл Т. (2004). «Человеческий Argonaute2 опосредует расщепление РНК, на которое направлены микроРНК и миРНК». Молекулярная клетка . 15 (2): 1403–1408. doi : 10.1016/j.molcel.2004.07.007 . ПМИД 15260970.
^ abc Mourelatos Z, Dostie J, Paushkin S, Sharma A, Charroux B, Abel L, Rappsilber J , Mann M, Dreyfuss G (2002). "miRNPs: a novel class of ribonucleoproteins contain multiple microRNAs". Genes & Development . 16 (6): 720–728. doi :10.1101/gad.974702. PMC 155365 . PMID 11914277.
^ Холл TM (2005). «Структура и функция белков Argonaute». Cell . 13 (10): 1403–1408. doi : 10.1016/j.str.2005.08.005 . PMID 16216572.
^ Чендримада TP, Грегори RI, Кумарасвами E, Норман J, Куч N, Нишикура K, Шиехаттер R (2005). «TRBP рекрутирует комплекс Dicer в Ago2 для обработки микроРНК и подавления генов». Nature . 436 (7051): 740–744. Bibcode :2005Natur.436..740C. doi :10.1038/nature03868. PMC 2944926 . PMID 15973356.
^ Wang HW, Noland C, Siridechadilok B, Taylor DW, Ma E, Felderer K, Doudna JA, Nogales E (2009). «Структурное понимание процесса обработки РНК комплексом загрузки RISC человека». Nature Structural & Molecular Biology . 16 (11): 1148–1153. doi :10.1038/nsmb.1673. PMC 2845538. PMID 19820710 .
^ Fu Q, Yuan YA (2013). «Структурное понимание сборки RISC, облегчаемое доменами связывания dsRNA спиралей РНК человека A (DHX9)». Nucleic Acids Research . 41 (5): 3457–3470. doi :10.1093/nar/gkt042. PMC 3597700. PMID 23361462 .
^ Yoo BK, Emdad L, Lee SG, Su Z, Santhekadur P, Chen D, Gredler R, Fisher PB, Sarkar D (2011). «Астроцитарный повышенный ген (AEG-1): многофункциональный регулятор нормальной и аномальной физиологии». Pharmacology & Therapeutics . 130 (1): 1–8. doi :10.1016/j.pharmthera.2011.01.008. PMC 3043119 . PMID 21256156.
^ Djikeng A, Shi H, Tschudi C, Shen S, Ullu E (2003). «Обнаружено, что siRNA-рибонуклеопротеин связан с полирибосомами Trypanosoma brucei». РНК . 9 (7): 802–808. doi :10.1261/rna.5270203. PMC 1370447 . PMID 12810914.
^ Сен GL, Верман TS, Блау HM (2005). «трансляция мРНК не является предпосылкой для расщепления мРНК, опосредованного малыми интерферирующими РНК». Дифференциация . 73 (6): 287–293. doi :10.1111/j.1432-0436.2005.00029.x. PMID 16138829.
^ Saumet A, Lecellier CH (2006). «Антивирусное подавление РНК: похожи ли мы на растения?». Retrovirology . 3 : 3. doi : 10.1186/1742-4690-3-3 . PMC 1363733. PMID 16409629 .
^ Bartel DP (2009). «МикроРНК: распознавание целей и регуляторные функции». Cell . 136 (2): 215–233. doi :10.1016/j.cell.2009.01.002. PMC 3794896 . PMID 19167326.
^ Jones-Rhoades MW, Bartel DP, Bartel B (2006). «МикроРНК и их регуляторные роли в растениях». Annual Review of Plant Biology . 57 : 19–53. doi :10.1146/annurev.arplant.57.032905.105218. PMID 16669754.
Дальнейшее чтение
Зонтхаймер, Э. Дж. (2005). «Сборка и функция комплексов подавления РНК». Nature Reviews Molecular Cell Biology . 6 (2): 127–138. doi :10.1038/nrm1568. PMID 15654322. S2CID 27294007.
Fu Q, Yuan YA (март 2013 г.). «Структурное понимание сборки RISC, облегчаемое доменами связывания dsRNA человеческой РНК-хеликазы A (DHX9)». Nucleic Acids Research . 41 (5): 3457–70. doi :10.1093/nar/gkt042. PMC 3597700. PMID 23361462 .
Schwarz DS, Tomari Y, Zamore PD (2004). «Комплекс подавления, вызванный РНК, представляет собой Mg2+-зависимую эндонуклеазу». Current Biology . 14 (9): 787–91. Bibcode :2004CBio...14..787S. doi : 10.1016/j.cub.2004.03.008 . PMID 15120070.
.jpg/1280px-Drosophila_melanogaster_-_side_(aka).jpg)