

В молекулярной биологии комплементарность описывает взаимосвязь между двумя структурами, каждая из которых следует принципу «замок и ключ». В природе комплементарность является базовым принципом репликации и транскрипции ДНК, поскольку это свойство, общее для двух последовательностей ДНК или РНК , так что когда они выровнены антипараллельно друг другу, нуклеотидные основания в каждой позиции в последовательностях будут комплементарными , как если бы вы смотрели в зеркало и видели обратную сторону вещей. Это комплементарное спаривание оснований позволяет клеткам копировать информацию из одного поколения в другое и даже находить и восстанавливать повреждения информации, хранящейся в последовательностях.
Степень комплементарности между двумя цепями нуклеиновых кислот может варьироваться от полной комплементарности (каждый нуклеотид находится напротив своей противоположности) до отсутствия комплементарности (каждый нуклеотид не находится напротив своей противоположности) и определяет стабильность последовательностей , которые должны быть вместе. Кроме того, различные функции восстановления ДНК, а также регуляторные функции основаны на комплементарности пар оснований. В биотехнологии принцип комплементарности пар оснований позволяет создавать гибриды ДНК между РНК и ДНК и открывает дверь для современных инструментов, таких как библиотеки кДНК . Хотя большая часть комплементарности наблюдается между двумя отдельными цепочками ДНК или РНК, также возможно, что последовательность имеет внутреннюю комплементарность, что приводит к связыванию последовательности с самой собой в свернутой конфигурации.

.png/1280px-Complementarity_(DNA).png)
Комплементарность достигается за счет различных взаимодействий между азотистыми основаниями : аденином , тимином ( урацилом в РНК ), гуанином и цитозином . Аденин и гуанин являются пуринами , в то время как тимин, цитозин и урацил являются пиримидинами . Пурины больше пиримидинов. Оба типа молекул дополняют друг друга и могут образовывать пары оснований только с противоположным типом азотистого основания. В нуклеиновой кислоте азотистые основания удерживаются вместе водородными связями , которые эффективно работают только между аденином и тимином и между гуанином и цитозином. Комплемент оснований A = T разделяет две водородные связи, в то время как пара оснований G ≡ C имеет три водородные связи. Все другие конфигурации между азотистыми основаниями будут препятствовать образованию двойной спирали. Цепи ДНК ориентированы в противоположных направлениях, они называются антипараллельными . [1]
Комплементарная цепь ДНК или РНК может быть сконструирована на основе комплементарности азотистых оснований. [2] Каждая пара оснований, A = T против G ≡ C, занимает примерно одинаковое пространство, тем самым обеспечивая образование скрученной двойной спирали ДНК без каких-либо пространственных искажений. Водородные связи между азотистыми основаниями также стабилизируют двойную спираль ДНК. [3]
Комплементарность цепей ДНК в двойной спирали позволяет использовать одну цепочку в качестве шаблона для построения другой. Этот принцип играет важную роль в репликации ДНК , закладывая основу наследственности , объясняя, как генетическая информация может передаваться следующему поколению. Комплементарность также используется в транскрипции ДНК , которая генерирует цепочку РНК из шаблона ДНК. [4] Кроме того, вирус иммунодефицита человека , одноцепочечный РНК-вирус , кодирует РНК-зависимую ДНК-полимеразу ( обратную транскриптазу ), которая использует комплементарность для катализа репликации генома. Обратная транскриптаза может переключаться между двумя родительскими РНК- геномами с помощью рекомбинации с выбором копии во время репликации. [5]
Механизмы репарации ДНК, такие как корректурное считывание , основаны на комплементарности и позволяют исправлять ошибки во время репликации ДНК путем удаления несовпадающих азотистых оснований. [1] В общем, повреждения в одной цепи ДНК можно исправить путем удаления поврежденного участка и его замены с помощью комплементарности для копирования информации с другой цепи, как это происходит в процессах репарации несовпадений , репарации эксцизии нуклеотидов и репарации эксцизии оснований . [6]
Цепи нуклеиновых кислот также могут образовывать гибриды , в которых одноцепочечная ДНК может легко отжигаться с комплементарной ДНК или РНК. Этот принцип лежит в основе широко применяемых лабораторных методов, таких как полимеразная цепная реакция , ПЦР. [1]
Две цепи комплементарной последовательности называются смысловой и антисмысловой . Смысловая цепь, как правило, представляет собой транскрибированную последовательность ДНК или РНК, которая была получена в результате транскрипции, в то время как антисмысловая цепь представляет собой цепь, комплементарную смысловой последовательности.

Самокомплементарность относится к тому факту, что последовательность ДНК или РНК может складываться сама на себя, создавая структуру, похожую на двойную цепочку. В зависимости от того, насколько близко друг к другу расположены части последовательности, которые являются самокомплементарными, цепочка может образовывать шпильковые петли, соединения, выпуклости или внутренние петли. [1] РНК с большей вероятностью образует такие типы структур из-за связывания пар оснований, не наблюдаемого в ДНК, например, связывания гуанина с урацилом. [1]

Комплементарность может быть обнаружена между короткими участками нуклеиновой кислоты и кодирующей областью или транскрибированным геном, и приводит к спариванию оснований. Эти короткие последовательности нуклеиновой кислоты обычно встречаются в природе и имеют регуляторные функции, такие как подавление генов. [1]
Антисмысловые транскрипты представляют собой участки некодирующей мРНК, которые комплементарны кодирующей последовательности. [7] Исследования по всему геному показали, что антисмысловые транскрипты РНК часто встречаются в природе. Обычно считается, что они увеличивают кодирующий потенциал генетического кода и добавляют общий уровень сложности к регуляции генов. На данный момент известно, что 40% человеческого генома транскрибируется в обоих направлениях, что подчеркивает потенциальную значимость обратной транскрипции. [8] Было высказано предположение, что комплементарные области между смысловыми и антисмысловыми транскриптами позволят генерировать двухцепочечные гибриды РНК, которые могут играть важную роль в регуляции генов. Например, мРНК фактора 1α, индуцированного гипоксией, и мРНК β-секретазы транскрибируются двунаправленно, и было показано, что антисмысловой транскрипт действует как стабилизатор смыслового сценария. [9]
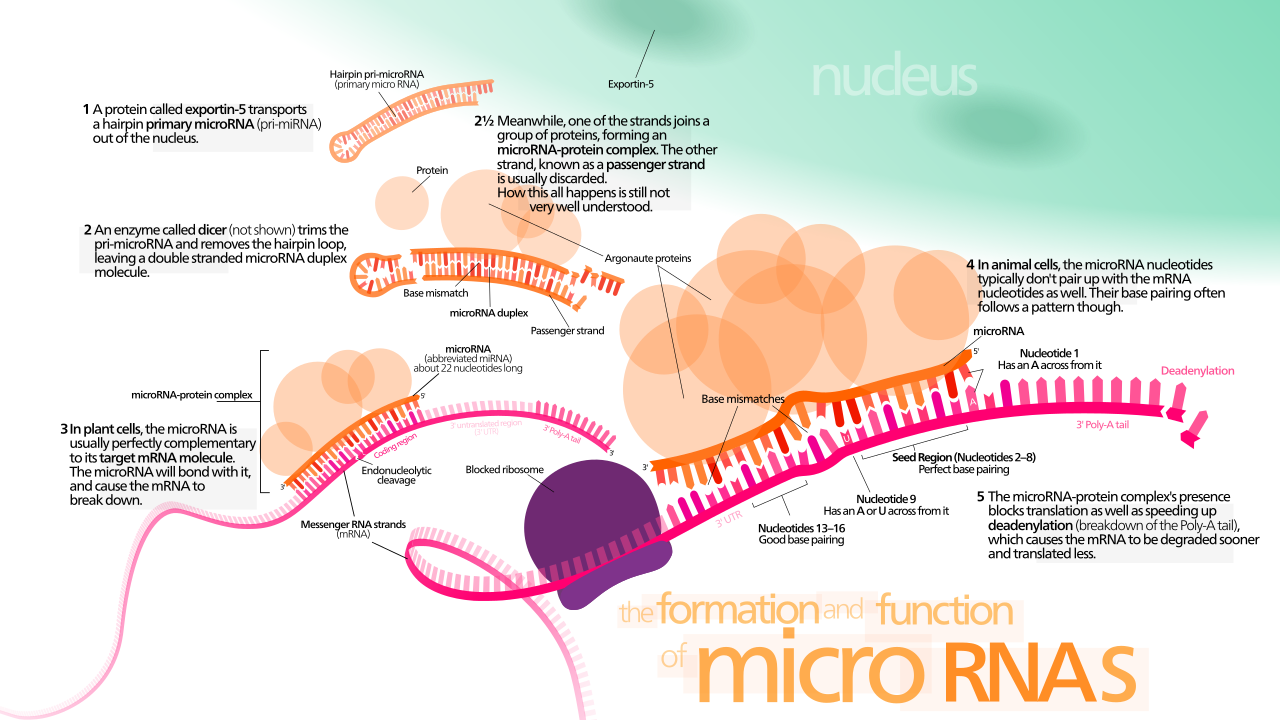
miRNAs , микроРНК, представляют собой короткие последовательности РНК, которые комплементарны областям транскрибируемого гена и выполняют регуляторные функции. Текущие исследования показывают, что циркулирующие miRNA могут использоваться в качестве новых биомаркеров, следовательно, демонстрируют многообещающие доказательства для использования в диагностике заболеваний. [10] MiRNAs образуются из более длинных последовательностей РНК, которые освобождаются ферментом Dicer от последовательности РНК, которая принадлежит регуляторному гену. Эти короткие нити связываются с комплексом RISC . Они совпадают с последовательностями в восходящей области транскрибируемого гена из-за своей комплементарности, чтобы действовать как сайленсер для гена тремя способами. Один из них заключается в том, чтобы не допустить связывания рибосомы и инициирования трансляции. Два — в том, чтобы деградировать мРНК, с которой связался комплекс. И третий — в том, чтобы предоставить новую последовательность двухцепочечной РНК (dsRNA), на которую Dicer может воздействовать, чтобы создать больше miRNA, чтобы найти и деградировать больше копий гена. Малые интерферирующие РНК (siRNA) по своей функции похожи на miRNA; они происходят из других источников РНК, но служат той же цели, что и miRNA. [1] Учитывая их короткую длину, правила комплементарности означают, что они все еще могут быть очень разборчивыми в своих целях выбора. Учитывая, что существует четыре выбора для каждого основания в цепи и длина mi/siRNA 20bp - 22bp, это приводит к более чем1 × 10 12 возможных комбинаций . Учитывая, что длина человеческого генома составляет ~3,1 миллиарда оснований, [11] это означает, что каждая miRNA должна найти совпадение во всем человеческом геноме только один раз случайно.
Целующиеся шпильки образуются, когда одна нить нуклеиновой кислоты дополняет сама себя, создавая петли РНК в форме шпильки. [12] Когда две шпильки вступают в контакт друг с другом in vivo , комплементарные основания двух нитей формируются и начинают раскручивать шпильки до тех пор, пока не образуется комплекс двухцепочечной РНК (dsRNA) или комплекс не раскручивается обратно на две отдельные нити из-за несоответствий в шпильках. Вторичная структура шпильки до поцелуя обеспечивает стабильную структуру с относительно фиксированным изменением энергии. [13] Целью этих структур является балансирование стабильности петли шпильки против прочности связывания с комплементарной нитью. Слишком сильное начальное связывание с плохим местом — и нити не раскрутятся достаточно быстро; слишком слабое начальное связывание — и нити никогда полностью не сформируют желаемый комплекс. Эти шпильковые структуры позволяют раскрыть достаточное количество оснований, чтобы обеспечить достаточно сильную проверку начального связывания и достаточно слабую внутреннюю связь, чтобы обеспечить развертывание после нахождения благоприятного соответствия. [13]
---С Г--- ЦГ ---ЦГ--- УАКГ GCUA CGGC АГЦГ АААГ КУАА U CUU ---CCUGCAACUUAGGCAGG--- А ГАА ---ГГАКГУУГААУККГУКК--- ГАУУ УУУК УКГЦ GCCG CGAU АУГК ГЦ ---Г Ц------Г Ц---Целующиеся шпильки встречаются наверху петель. Взаимодополняемостьиз двух головок побуждает шпильку разворачиваться и выпрямляться, чтобыстать одной плоской последовательностью из двух прядей, а не двумя шпильками.
Комплементарность позволяет хранить информацию, содержащуюся в ДНК или РНК, в одной цепи. Комплементарная цепь может быть определена из шаблона и наоборот, как в библиотеках кДНК. Это также позволяет проводить анализ, например, сравнивать последовательности двух разных видов. Были разработаны стенографии для записи последовательностей при наличии несоответствий (коды неоднозначности) или для ускорения чтения противоположной последовательности в комплементе (амбиграммы).
Библиотека кДНК представляет собой коллекцию экспрессированных генов ДНК, которые рассматриваются как полезный справочный инструмент в процессах идентификации и клонирования генов. Библиотеки кДНК создаются из мРНК с использованием РНК-зависимой ДНК-полимеразы обратной транскриптазы (RT), которая транскрибирует шаблон мРНК в ДНК. Поэтому библиотека кДНК может содержать только вставки, которые предназначены для транскрибации в мРНК. Этот процесс основан на принципе комплементарности ДНК/РНК. Конечным продуктом библиотек является двухцепочечная ДНК, которая может быть вставлена в плазмиды. Таким образом, библиотеки кДНК являются мощным инструментом в современных исследованиях. [1] [14]
При написании последовательностей для систематической биологии может потребоваться иметь коды IUPAC , которые означают «любой из двух» или «любой из трех». Код IUPAC R (любой пурин ) является дополнительным к Y (любой пиримидин ), а M (амино) — к K (кето). W (слабый) и S (сильный) обычно не меняются местами [15], но были заменены в прошлом некоторыми инструментами. [16] W и S обозначают «слабый» и «сильный» соответственно и указывают на количество водородных связей, которые нуклеотид использует для спаривания со своим комплементарным партнером. Партнер использует то же количество связей для создания комплементарной пары. [17]
Код IUPAC, который специально исключает один из трех нуклеотидов, может быть комплементарным коду IUPAC, который исключает комплементарный нуклеотид. Например, V (A, C или G - "не T") может быть комплементарным B (C, G или T - "не A").
Определенные символы могут быть использованы для создания подходящей ( амбиграфической ) записи нуклеиновой кислоты для комплементарных оснований (например, гуанин = b , цитозин = q , аденин = n и тимин = u ), что позволяет дополнять целые последовательности ДНК, просто поворачивая текст «вверх ногами». [19] Например, с предыдущим алфавитом buqn (GTCA) будет читаться как ubnq (TGAC, обратный комплемент), если перевернуть его вверх ногами.
Амбиграфические обозначения легко визуализируют комплементарные участки нуклеиновых кислот, такие как палиндромные последовательности. [20] Эта функция улучшается при использовании пользовательских шрифтов или символов вместо обычных символов ASCII или даже Unicode. [20]
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite journal}}: CS1 maint: числовые имена: список авторов ( ссылка )