Уравнения Лотки–Вольтерры , также известные как модель хищник–жертва Лотки–Вольтерры , представляют собой пару нелинейных дифференциальных уравнений первого порядка , часто используемых для описания динамики биологических систем , в которых взаимодействуют два вида, один как хищник , а другой как добыча. Популяции изменяются со временем в соответствии с парой уравнений:
где
Решение дифференциальных уравнений является детерминированным и непрерывным . Это, в свою очередь, подразумевает, что поколения как хищника, так и жертвы постоянно перекрываются. [1]
Система уравнений Лотки–Вольтерры является примером популяционной модели Колмогорова (не путать с более известными уравнениями Колмогорова ) [2] [3] [4] , которая представляет собой более общую структуру, позволяющую моделировать динамику экологических систем с взаимодействиями хищник–жертва, конкуренцией , болезнями и мутуализмом .
Предполагается, что добыча имеет неограниченный запас пищи и размножается экспоненциально, если только не подвергается хищничеству; этот экспоненциальный рост представлен в уравнении выше членом αx . Предполагается, что скорость хищничества добычи пропорциональна скорости, с которой встречаются хищники и добыча; это представлено выше членом βxy . Если x или y равны нулю, то хищничества быть не может. С этими двумя членами уравнение добычи выше можно интерпретировать следующим образом: скорость изменения популяции добычи определяется ее собственной скоростью роста за вычетом скорости, с которой на нее охотятся.
Член δxy представляет рост популяции хищников. (Обратите внимание на сходство со скоростью хищничества; однако используется другая константа, поскольку скорость, с которой растет популяция хищников, не обязательно равна скорости, с которой он потребляет добычу). Член γy представляет скорость потери хищников из-за естественной смерти или эмиграции; это приводит к экспоненциальному спаду при отсутствии добычи. Следовательно, уравнение выражает, что скорость изменения популяции хищника зависит от скорости, с которой он потребляет добычу, за вычетом его собственной скорости смертности.
Модель хищник-жертва Лотки-Вольтерры делает ряд предположений относительно окружающей среды и биологии популяций хищников и жертв: [5]
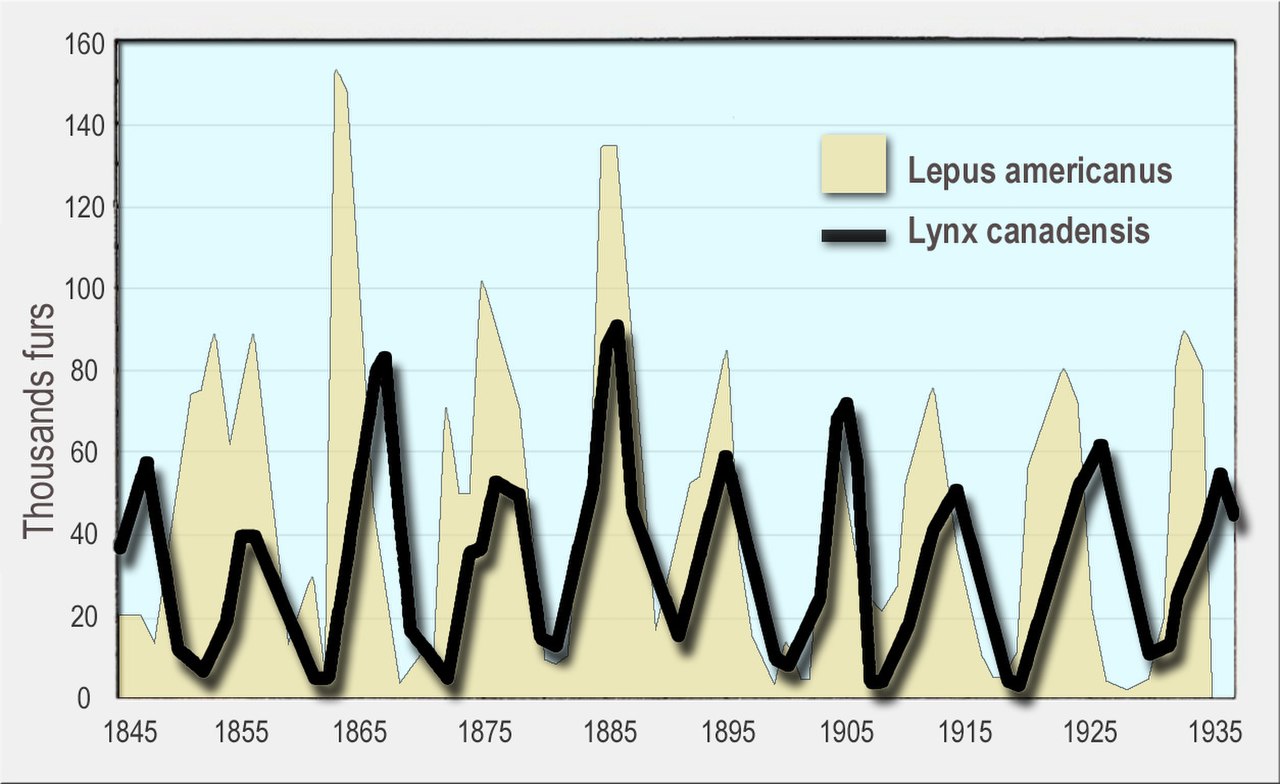
Ни одно из вышеприведенных предположений, вероятно, не будет справедливым для естественных популяций. Тем не менее, модель Лотки–Вольтерры показывает два важных свойства популяций хищников и жертв, и эти свойства часто распространяются на варианты модели, в которых эти предположения смягчены:
Во-первых, динамика популяций хищников и жертв имеет тенденцию к колебаниям. Колебания численности хищников и жертв наблюдались в естественных популяциях, таких как данные по рысям и зайцам-белякам компании Hudson's Bay Company [6] и популяции лосей и волков в национальном парке Isle Royale . [7]
Во-вторых, равновесие популяции этой модели обладает свойством, что равновесная плотность добычи (задается как ) зависит от параметров хищника, а равновесная плотность хищника (задается как ) от параметров жертвы. Это имеет следствием то, что увеличение, например, скорости роста добычи, , приводит к увеличению равновесной плотности хищника, но не равновесной плотности добычи. Улучшение среды обитания для добычи приносит пользу хищнику, а не добыче (это связано с парадоксом пестицидов и парадоксом обогащения ). Демонстрацией этого явления является увеличение процента вылавливаемых хищных рыб в годы Первой мировой войны (1914–18), когда скорость роста добычи увеличилась из-за сокращения рыболовного усилия.
Еще одним примером является экспериментальное удобрение океана железом. В нескольких экспериментах в океане растворялось большое количество солей железа. Ожидалось, что железо, которое является ограничивающим питательным веществом для фитопланктона, будет стимулировать рост фитопланктона и что оно будет поглощать углекислый газ из атмосферы. Добавление железа обычно приводит к кратковременному цветению фитопланктона, который быстро потребляется другими организмами (такими как мелкая рыба или зоопланктон ) и ограничивает эффект обогащения в основном увеличением плотности хищников, что, в свою очередь, ограничивает поглощение углерода. Это предсказывается равновесными плотностями популяции модели хищник-жертва Лотки-Вольтерры и является особенностью, которая переносится на более сложные модели, в которых ограничивающие предположения простой модели смягчены. [8]
Модель Лотки–Вольтерры имеет дополнительные приложения в таких областях, как экономика [9] и маркетинг. [10] [11] Ее можно использовать для описания динамики на рынке с несколькими конкурентами, взаимодополняющими платформами и продуктами, экономикой совместного потребления и т. д. Существуют ситуации, в которых один из конкурентов вытесняет других конкурентов с рынка, и другие ситуации, в которых рынок достигает равновесия, где каждая фирма стабилизируется на своей доле рынка. Также можно описать ситуации, в которых происходят циклические изменения в отрасли или хаотические ситуации без равновесия, а изменения происходят часто и непредсказуемо.
Модель хищник–жертва Лотки–Вольтерры была первоначально предложена Альфредом Дж. Лотка в теории автокаталитических химических реакций в 1910 году. [12] [13] Это было фактически логистическое уравнение , [14] первоначально выведенное Пьером Франсуа Ферхюльстом . [15] В 1920 году Лотка расширил модель с помощью Андрея Колмогорова на «органические системы», используя в качестве примера вид растений и вид травоядных животных [16] , а в 1925 году он использовал уравнения для анализа взаимодействий хищник–жертва в своей книге по биоматематике . [17] Тот же набор уравнений был опубликован в 1926 году Вито Вольтеррой , математиком и физиком, который заинтересовался математической биологией . [13] [18] [19] Исследование Вольтерры было вдохновлено его взаимодействием с морским биологом Умберто Д'Анкона , который в то время ухаживал за его дочерью, а позже стал его зятем. Д'Анкона изучал уловы рыбы в Адриатическом море и заметил, что процент пойманной хищной рыбы увеличился в годы Первой мировой войны (1914–18). Это озадачило его, так как промысловое усилие было очень сильно сокращено в военные годы, и, поскольку предпочтительнее была добыча рыбы, можно было бы интуитивно ожидать, что это увеличит процент добычи рыбы. Вольтерра разработал свою модель, чтобы объяснить наблюдение Д'Анконы, и сделал это независимо от Альфреда Лотки. Он действительно отдал должное более ранней работе Лотки в своей публикации, после чего модель стала известна как «модель Лотки-Вольтерры». [20]
Позднее модель была расширена, включив в нее рост добычи, зависящий от плотности, и функциональную реакцию в форме, разработанной К. С. Холлингом ; модель, которая стала известна как модель Розенцвейга–МакАртура. [21] Обе модели, Лотки–Вольтерры и Розенцвейга–МакАртура, использовались для объяснения динамики естественных популяций хищников и добычи.
В конце 1980-х годов появилась альтернатива модели хищник-жертва Лотки-Вольтерры (и ее обобщениям, зависящим от общей жертвы), модель, зависящая от отношения, или модель Ардити-Гинзбурга . [22] Обоснованность моделей, зависящих от отношения или добычи, была предметом многочисленных споров. [23]
Уравнения Лотки–Вольтерры имеют долгую историю использования в экономической теории ; их первоначальное применение обычно приписывают Ричарду Гудвину в 1965 [24] или 1967 годах. [25] [26]
Уравнения имеют периодические решения. Эти решения не имеют простого выражения в терминах обычных тригонометрических функций , хотя они вполне поддаются обработке. [27] [28] [29]
Если ни один из неотрицательных параметров α , β , γ , δ не обращается в нуль, три из них можно поглотить в нормализацию переменных, оставив только один параметр: поскольку первое уравнение однородно по x , а второе — по y , параметры β / α и δ / γ поглощаются в нормализациях y и x соответственно, а γ — в нормализацию t , так что произвольным остается только α / γ . Это единственный параметр, влияющий на характер решений.
Линеаризация уравнений дает решение, похожее на простое гармоническое движение [30], при этом популяция хищников отстает от популяции жертв на 90° в цикле.

.svg/1280px-Lotka-Volterra_model_(1.1,_0.4,_0.4,_0.1).svg.png)

Предположим, что есть два вида животных: кролик (добыча) и лиса (хищник). Если начальная плотность составляет 10 кроликов и 10 лис на квадратный километр, можно построить график изменения численности двух видов с течением времени; при условии, что показатели роста и смертности кроликов составляют 1,1 и 0,4, а лис — 0,1 и 0,4 соответственно. Выбор временного интервала произволен.
Можно также параметрически построить решения в виде орбит в фазовом пространстве , не отображая время, но с одной осью, представляющей количество добычи, а другой осью, представляющей плотность хищников для всех моментов времени.
Это соответствует исключению времени из двух дифференциальных уравнений выше для получения одного дифференциального уравнения
связывая переменные x (хищник) и y (жертва). Решения этого уравнения — замкнутые кривые. Оно поддается разделению переменных : интегрирование
дает неявную связь
где V — постоянная величина, зависящая от начальных условий и сохраняющаяся на каждой кривой.
Примечание: Эти графики иллюстрируют серьезное потенциальное ограничение в применении в качестве биологической модели: для этого конкретного выбора параметров в каждом цикле популяция кроликов сокращается до крайне низких значений, но восстанавливается (в то время как популяция лис остается значительной при самой низкой плотности кроликов). Однако в реальных ситуациях случайные колебания дискретного числа особей могут привести к фактическому вымиранию кроликов, а следовательно, и лис. Эта проблема моделирования была названа «проблемой атто-лисы», атто - лиса представляет собой условные 10−18 лисы . [31] [32] Плотность 10−18 лис на квадратный километр соответствует в среднем приблизительно 5×10−10 лис на поверхности Земли, что на практике означает, что лисы вымерли.
Поскольку величина сохраняется с течением времени, она играет роль функции Гамильтона системы. [33] Чтобы увидеть это, мы можем определить скобку Пуассона следующим образом . Тогда уравнения Гамильтона будут иметь вид Переменные и не являются каноническими, так как . Однако, используя преобразования [34] , мы пришли к канонической форме уравнений Гамильтона, включающей гамильтониан : Скобка Пуассона для канонических переменных теперь принимает стандартный вид .

Менее экстремальный пример:
α = 2/3 , β = 4/3 , γ = 1 = δ . Предположим, что x , y количественно измеряют тысячи каждый. Круги представляют начальные условия добычи и хищника от x = y = 0,9 до 1,8 с шагом 0,1. Фиксированная точка находится в точке (1, 1/2).
В модельной системе хищники процветают, когда добыча в изобилии, но в конечном итоге превосходят свои запасы пищи и приходят в упадок. Когда популяция хищников низкая, популяция добычи снова увеличится. Эта динамика продолжается в цикле роста и упадка популяции .
Равновесие численности населения в модели достигается, когда ни один из уровней численности населения не изменяется, т.е. когда обе производные равны 0:
Приведенная выше система уравнений дает два решения: и
Следовательно, существует два равновесия.
Первое решение фактически представляет вымирание обоих видов. Если обе популяции равны 0, то они будут оставаться такими неопределенно долго. Второе решение представляет фиксированную точку, в которой обе популяции поддерживают свои текущие, ненулевые численности, и, в упрощенной модели, делают это неопределенно долго. Уровни популяции, при которых достигается это равновесие, зависят от выбранных значений параметров α , β , γ и δ .
Устойчивость неподвижной точки в начале координат можно определить, выполнив линеаризацию с использованием частных производных .
Матрица Якоби модели «хищник-жертва» известна как матрица сообщества .
При оценке в устойчивом состоянии (0, 0) матрица Якоби J становится
Собственные значения этой матрицы равны
В модели α и γ всегда больше нуля, и, таким образом, знак собственных значений выше всегда будет отличаться. Следовательно, неподвижная точка в начале координат является седловой точкой .
Нестабильность этой фиксированной точки имеет значение. Если бы она была стабильной, ненулевые популяции могли бы притягиваться к ней, и, как таковая, динамика системы могла бы привести к вымиранию обоих видов для многих случаев начальных уровней популяции. Однако, поскольку фиксированная точка в начале координат является седловой точкой и, следовательно, нестабильна, из этого следует, что вымирание обоих видов в модели затруднено. (На самом деле, это могло бы произойти только в том случае, если бы добыча была полностью уничтожена искусственно, в результате чего хищники умерли бы от голода. Если бы хищники были уничтожены, популяция добычи неограниченно росла бы в этой простой модели.) Популяции добычи и хищника могут приближаться к нулю на бесконечно малую величину и все равно восстанавливаться.
Оценка J во второй фиксированной точке приводит к
Собственные значения этой матрицы равны
Поскольку собственные значения являются чисто мнимыми и сопряженными друг с другом, эта неподвижная точка должна быть либо центром для замкнутых орбит в локальной окрестности, либо притягивающей или отталкивающей спиралью. В консервативных системах должны быть замкнутые орбиты в локальной окрестности неподвижных точек, которые существуют в минимумах и максимумах сохраняющейся величины. Сохраняющаяся величина выведена выше как находящаяся на орбитах. Таким образом, орбиты вокруг неподвижной точки являются замкнутыми и эллиптическими , поэтому решения являются периодическими, колеблющимися на малом эллипсе вокруг неподвижной точки с частотой и периодом .
Как показано на циркуляционных колебаниях на рисунке выше, кривые уровня представляют собой замкнутые орбиты, окружающие фиксированную точку: уровни популяций хищников и жертв циклически и колеблются без затухания вокруг фиксированной точки с частотой .
Значение постоянной движения V , или, что то же самое, K = exp(− V ) , можно найти для замкнутых орбит вблизи неподвижной точки.
Увеличение K приближает замкнутую орбиту к неподвижной точке. Наибольшее значение константы K получается путем решения задачи оптимизации. Максимальное значение K при этом достигается в стационарной (неподвижной) точке и составляет где e — число Эйлера .





































