Этозавры ( / eɪ ˌ ɛ t oʊ ˈ s ɔːr / ) — тяжелобронированные рептилии, принадлежащие к вымершему отряду Aetosauria ( / eɪ ˌ ɛ t oʊ ˈ s ɔːr i ə / ; от греческого ἀετός (aetos, «орел») и σαυρος ( саурос , «ящерица»)). Это были всеядные или травоядные псевдозухи среднего и крупного размера , часть ветви архозавров, более тесно связанных с крокодилами , чем с птицами и другими динозаврами . Все известные этоозавры относятся к позднему триасу , а в некоторых слоях этого времени они входят в число наиболее многочисленных ископаемых позвоночных . У них маленькие головы, перевернутая морда, прямые конечности и тело, украшенное четырьмя рядами пластинчатых остеодерм (костных щитков ). Ископаемые останки этозавра известны в Европе , Северной и Южной Америке , некоторых частях Африки и Индии . Поскольку их бронированные пластины часто сохраняются и многочисленны в определенных местах, этозавры служат важными окаменелостями-индексами четвероногих позднего триаса . Многие этозавры имели широкие географические ареалы, но их стратиграфические ареалы были относительно короткими. Таким образом, присутствие определенных этозавров позволяет точно датировать место их обнаружения. [1] [2] [3]
Почти все этоозавры (кроме рода Aetosauroides ) относятся к семейству Stagonolepididae . Описано более 20 родов этозавров, и в последнее время возникли разногласия по поводу описания некоторых из этих родов. В настоящее время выделяются два отдельных подразделения этозавров: Desmatosuchia и Aetosaurinae , что основано главным образом на широких различиях в морфологии черепа . [4] Структура остеодермы, как правило, является одной из наиболее полезных черт для более точного определения взаимоотношений этозавров. Среди других архозавров этоозавры наиболее тесно связаны с ревуэльтозавром , небольшой рептилией, первоначально известной по зубам, ошибочно отнесенной к травоядным динозаврам. [3]
Останки этозавра были впервые обнаружены в начале 19 века, хотя самые первые описанные останки были приняты за рыбью чешую . Позже этозавров признали родственниками крокодилов , после чего их стали интерпретировать как полуводных падальщиков, тесно связанных с фитозаврами . Последующие работы установили, что этоозавры были полностью наземными животными и, вероятно, в некоторой степени были травоядными. Некоторые формы имеют характеристики, которые могли быть адаптацией к рытью в поисках пищи. К этозаврам относили и предполагаемые гнездовые сооружения, но эта связь считается неоднозначной. [3]

Череп этозавров относительно невелик по сравнению с телом и весьма своеобразен по форме. Зубы отсутствуют как в передней части предчелюстной кости (кость, образующая кончик морды), так и в передней части зубной кости (зубчатая кость нижней челюсти). [5] [4] Имеющиеся зубы обычно маленькие и выпуклые, от основной конической формы до листовидной формы с большими зубцами. Вероятно, это свидетельствует о всеядности или травоядности , и аналогичные адаптации наблюдаются у других архозавров, которые меньше полагаются на мясо в своем рационе. [6] [3] У некоторых этозавров зубы имеют форму зифодонта, что означает, что зубы загнуты, зазубрены и сплющены сбоку. Эта форма, преобладающая у Aetosauroides и небольшой экземпляр, условно отнесенный к Coahomasuchus , [4] типична для хищных архозавров. [7]
У некоторых этозавров (особенно представителей группы Desmatosuchia) кончик морды расширен вбок и имеет уплощенную форму «лопаты», похожую на морду свиньи . Наружные ноздри (ноздри) удлиненные, намного больше, чем предглазничные отверстия (отверстие на боковой стороне черепа). У многих этозавров на предчелюстной кости есть небольшой выступ, который снизу выступает в ноздри. У всех этозавров, кроме Aetosauroides , задний край ноздри получает вклад от вогнутого переднего края верхнечелюстной кости . В задней верхней части черепа отверстие, известное как надвисочное окно, расположено и открыто сбоку, в отличие от большинства других архозавров, у которых оно в основном видно при взгляде на череп сверху. [8] [5] [4] Черепная коробка довольно стандартна по псевдозуховым стандартам, хотя отверстие для отводящего нерва проходит через парабазисфеноидальную кость (в нижней передней части черепной коробки), а не через проотическую кость (в верхней части черепной коробки). Передняя часть). В остальном эта черта наблюдается только у ревуэльтозавра и крокодиломорфов среди архозавров. [9] [5] [10] Нижняя челюсть (нижняя челюсть) у многих этозавров имеет форму «тапочки». Это происходит из-за комбинации особенностей: передняя часть зубной кости сильно сужается к концу, тогда как нижняя часть зубной кости иногда изгибается в «подбородок» (проекция вниз), что также может обнажать селезеночную кость . Челюстной сустав расположен низко, а суставная часть (кость нижней челюсти, соединяющаяся с черепом) часто имеет высокий выступ сразу за челюстным суставом. [8] [4] [7]

Во многих отношениях, за исключением черепа и доспехов, анатомия скелета этозавров была довольно стандартной среди других крупных псевдозухов триаса. Задние конечности приобрели «вертикальную» позу конечностей, аналогичную той, что наблюдается у « rauisurians », родственной разновидности плотоядных триасовых псевдозухий, предков крокодиломорфов. [1] Поза конечностей с выпрямленным столбом – это поза, при которой бедренная кость сочленяется вертикально с вертлужной впадиной бедра, которая наклонена вниз, так что нога располагается под телом и действует как опора, несущая вес. Тем не менее, вероятно, существовали значительные различия в строении бедер этозавров, а передние конечности могли иметь полураскидистую «гибридную» позицию. Хотя поза задних конечностей аналогична рауизухиям, другие черты более плезиоморфны (типичны для предков псевдозухий), например, толстый таз и широкие пятипалые ступни. [6] Передние конечности были меньше задних, а лучевая кость , в частности, была намного короче плечевой кости . Тем не менее, их низкая и тяжелая форма тела требует, чтобы все этозавры были четвероногими . У них было множество приспособлений для укрепления тела в ответ на их тяжелую броню: подвздошно-фибулярная мышца прикреплялась к нижнему положению на малоберцовой кости , четвертый вертел бедренной кости был увеличен, поперечные отростки (реберные крепления) развились в длинные массивные пьедесталы и самые крупные виды даже приобрели усиление гипосфен-гипантрум между позвонками. [3] [4]
Хотя этоозавры в целом были широкотелыми рептилиями, существуют некоторые различия в степени этой тенденции. Типоторацины , примером которых являются Typothorax и Paratypothorax , имели очень широкий дискообразный панцирь, окаймленный небольшими шипами или килями и переходящий в узкий хвост. Самый крупный вид типоторацинов мог иметь длину около 3 метров (9,8 фута) и вес 110 кг (243 фунта). Десматозухины ( Desmatosurinae sensu stricto ), такие как Desmatosuchus и Longosurus , имели умеренно более узкое тело и не имели брюшного панциря. Однако они также приобрели более шипастую спинную броню, особенно в шейной области. [11] Десматозух , вероятно, был одним из крупнейших известных этозавров, его длина составляла 4–6 м (13–20 футов), а вес - 280 кг (620 фунтов). [12] [3] [13] Этозавры, которые не вписываются в эти две категории, такие как Stagonolepis и Neoaetosauroides , обычно имели узкую форму, тонкие конечности и ограничение панциря над бедром. [3] Этот тип телосложения является плезиоморфным (предковым) по отношению к двум другим формам, при этом некоторые узкотелые этозавры более тесно связаны с типоторацинами, а другие ближе к десматозухинам. [4] Некоторые плезиоморфные роды, такие как широко распространенный норианский род Aetosaurus и карнийский Coahomasuchus , как правило, были небольшими, около метра (3,2 фута) в длину. [14] Другие были крупнее, например, самый базальный этозавр Aetosauroides и ранний десматозухин Calyptosurus . [3]

Этозавры имели очень тяжелую броню, состоящую из рядов больших переплетенных костных пластин, известных как остеодермы , защищающих спину, бока, живот и хвост. [6] Эти остеодермы обычно имеют четырехугольную (четырехстороннюю) форму и наверняка использовались в качестве защиты от хищников. У большинства остеодерм верхняя поверхность сильно изрыта, а нижняя гладкая. Они имеют гетерогенную внутреннюю структуру: внутренняя часть каждой остеодермы состоит из губчатой или губчатой кости (также называемой диплоэ ), а их внешние части состоят из компактной кости. [3] При жизни эти пластинки, вероятно, были покрыты ороговевшим (роговым) покрытием, как и щитки современных крокодилов , которые являются еще одним примером псевдозуховых остеодерм. [15] Остеодермы полезны для диагностики таксонов этозавров, а виды этозавров часто можно идентифицировать по отдельным щиткам на основе их формы, структуры или рисунка орнамента. [16] [17] [3]
У этозавров есть четыре ряда остеодерм, идущих вдоль их дорсальной (задней) стороны, образующих непрерывную пластину, часто называемую панцирем . Два внутренних ряда, обрамляющих среднюю линию позвоночного столба, известны как парамедианные остеодермы. Они, как правило, шире, чем длинны, и сильно украшены расходящимися ямками или бороздками. Почти у всех этозавров на верхней поверхности каждой пластины имеется небольшой выступ или приподнятая поверхность, известная как спинное возвышение. Дорсальное возвышение часто располагается сзади (назад) или медиально (внутри) на соответствующей парамедиальной остеодерме, хотя внутри группы есть много исключений. Парамедианные остеодермы почти всегда имеют приподнятые или вдавленные передние края, где пластинки перекрываются передними. Если передний край приподнят, эта область называется передней перемычкой, а если он вдавлен, то эта область называется передней пластинкой. Хотя два парамедиальных ряда остеодермы обычны для ранних архозавроформ, лишь немногие рептилии приближаются к этозаврам по сложности их остеодерм. Остеодермы досвеллиид , эрпетозухид и некоторых крокодиломорфов иногда путают или сравнивают с остеодермами этозавров. И передняя перемычка, и дорсальное возвышение встречаются у Acaenasuchus , близкого родственника этозавров, первоначально ошибочно идентифицированных как ювенильный десматозухин. [8] [3] [4] [18]

Два внешних ряда остеодерм, лежащих рядом с парамедианами, известны как латеральные остеодермы. Они параллельны парамедианам почти на всей задней стороне, хотя первые два парамедиана позади головы (известные как выйные остеодермы) одиночные. Боковые пластинки обычно разделены на две поверхности или фланцы, согнутые между своими дорсальными выступами. Верхний, или дорсальный, фланец лежит в одной плоскости с парамедианными остеодермами. Нижний/внешний или боковой фланец заворачивается на боковую часть корпуса. Спинное возвышение между этими выступами часто имеет форму невысокого лезвия, набалдашника или шипа. В шейных латеральных остеодермах, расположенных на шее, дорсальное возвышение имеет тенденцию проявляться в виде выступающего шипа. [19] Это доведено до крайности у десматозухинов, таких как Longosurus и Desmatosuchus , у которых шип увеличен в резко изогнутый рог. [8]
У большинства этозавров (за исключением десматозухинов ) нижняя часть тела животного также защищена остеодермами. Эти вентральные (брюшные) остеодермы обычно меньше и более плоские, чем дорсальный ряд, и расположены в большее число рядов (обычно 5–14 рядов), по крайней мере на туловище. Вентральные ряды остеодерм обычно выгибаются наружу и разделяются под бедром, оставляя широкий зазор для клоакального отверстия. Вокруг этого отверстия у Typothorax встречаются большие крючковатые шипы , одно из немногих исключений из общего правила гладких вентральных остеодерм. Брюшные ряды распадаются на шагрень из мелких пластинок на шее и небольшое количество широких рядов под хвостом. Передние и задние конечности покрывал густой набор небольших непересекающихся пластинок, известных как аппендикулярные остеодермы. [3] [20]
Материал этозавра был впервые описан швейцарским палеонтологом Луи Агассисом в 1844 году. Он назвал род Stagonolepis из песчаника Лоссимута в Элгине , Шотландия , но считал, что ископаемое принадлежит девонской рыбе, а не триасовой рептилии. Возможно, это связано с тем, что он считал эти слои частью старого красного песчаника и, следовательно, палеозойского возраста. Агассис принял остеодермы за большие ромбовидные чешуйки, которые, по его мнению, были расположены по такому же принципу, как и у гарса . Он также считал, что эти предполагаемые чешуи очень похожи на чешую лопастной рыбы Megalichthys из-за их большого размера. [21]
Английский биолог Томас Генри Хаксли пересмотрел рыбью чешую, описанную Агассисом, и посчитал, что она принадлежит крокодилу. Впервые он предложил это Лондонскому геологическому обществу в 1858 году и более подробно рассказал об этом в статье 1875 года в ежеквартальном журнале общества. К этому времени в Элгине был обнаружен новый материал, указывающий на то, что Стагонолепис был не рыбой, а рептилией. Однако Stagonolepis все еще был известен прежде всего по щиткам и отпечаткам щитков, многие из которых сохранились плохо. [22]

Более полные останки этозавра были найдены в Нижнем Штубенсандтайне в Германии в 1870-х годах. Среди них были полные сочлененные скелеты 22 этозавров. Эти экземпляры были найдены в большом обнажении песчаника недалеко от Штутгарта и сохранились вместе на площади менее 2 квадратных метров. Животные, вероятно, были погребены под осадком озера вскоре после смерти, а поток воды переместил их тела на дно озера и поставил их в непосредственной близости друг к другу. В 1877 году немецкий палеонтолог Оскар Фраас отнес эти экземпляры к недавно созданному роду Aetosaurus . Фраас назвал род в честь сходства черепа с головой орла, с узким удлиненным черепом и заостренной мордой. [23]
Первые этозавры из Северной Америки также были описаны в 1870-х годах. Американский палеонтолог Эдвард Дринкер Коуп назвал Типоторакса в 1875 году и Эпископозавра в 1877 году, оба были из Нью-Мексико . Однако Коуп считал, что эти роды — фитозавры , крокодилоподобные водные архозавры, которые также были распространены в позднем триасе. Хотя в последние годы на юго-западе Америки был обнаружен новый материал, относящийся к типотораксу , эпископозавр больше не считается действительным. Коуп, вместе с более поздними палеонтологами, такими как Фридрих фон Хюне , признал, что останки типового вида E. horridus на самом деле принадлежали Typothorax . В то время Коуп считал, что этозаврия принадлежит к Rhynchocephalia , отряду рептилий, в который входят современные туатары . Он также считал, что плотно прилегающие остеодермы, которые, как считалось, срослись с ребрами, указывают на то, что этозавры были переходными формами между ринхоцефалами и черепахами . [24] Другой вид, E. haplocerus , был отнесен к Desmatosuchus в 1953 году. [25]

Третий североамериканский род, названный Stegomus , был назван Отниэлем Чарльзом Маршем в 1896 году . конкурируют друг с другом в этой области и в научной литературе. В отличие от этозавров Коупа, Стегомус был найден на востоке США, в Коннектикуте . Марш также признал Стегомуса этозавром, а не фитозавром в своем первоначальном описании рода. Как и Коуп, многие палеонтологи в этот период времени были склонны считать щитки этозавров принадлежащими фитозаврам. [17] Марш считал этоозавров близкими родственниками динозавров на основании их удлиненных плюсневых костей (костей стопы). [24]
Как отдельная группа, этозаврия была названа в 1889 году английским натуралистом Ричардом Лидеккером и зоологом Генри Аллейном Николсоном . [24] Они считали Aetosauria одним из трех подотрядов отряда Crocodilia, два других - Parasuria (группа, включающая фитозавров и другие триасовые формы) и Eusuria (группа, включающая всех посттриасовых крокодиломорфов). Николсон и Лидеккер поместили одно семейство в подотряд Aëtosauridae. Они считали, что этозавры похожи на ныне живущих крокодилов, несмотря на их более длинные плюсневые кости. [24]
Материал этозавра продолжали описывать до начала 20 века, когда образцы собирали известные палеонтологи, такие как Барнум Браун и Чарльз Кэмп . [27] Более качественные останки американских этозавров помогли получить анатомическую информацию о некоторых распространенных видах, таких как Desmatosuchus spurensis (подробно описанный EC Case в 1922 году) и Typothorax meadei (ныне Longosurus , описанный HJ Sawin в 1947 году). [28] Однако останки этозавров все еще путались с останками фитозавров, и их по-прежнему считали преимущественно плотоядными и полуводными «паразухиями». Переописание Stagonolepis robertsoni, сделанное А. Д. Уокером в 1961 году , помогло изменить эти представления и создать более современный взгляд на анатомию и экологию этозавров. [29]
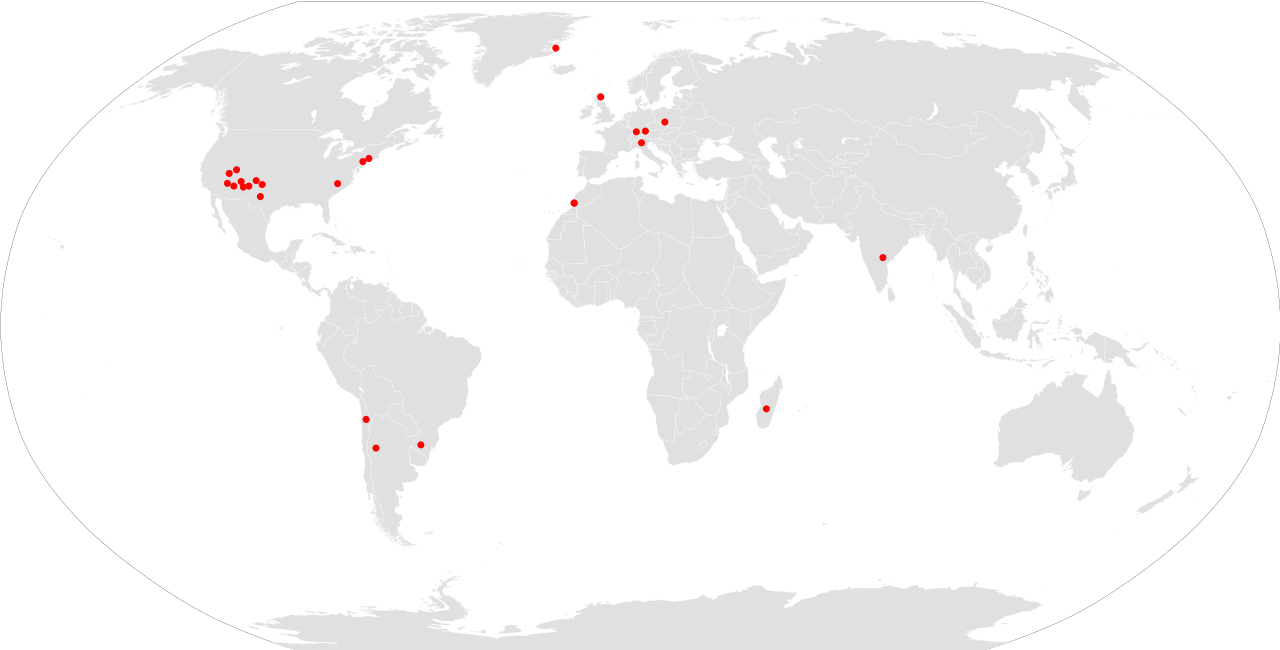
Окаменелости этозавров были обнаружены на всех континентах, кроме Австралии и Антарктиды , что обеспечило их распространение почти по всему миру в позднем триасе. Останки этозавров наиболее распространены на северных ( Лавразийских ) континентах, а окаменелости особенно распространены в формации Чинл и группе Докум на юго-западе США . Большинство окаменелостей было найдено в Аризоне , Нью-Мексико и Техасе . [30] Некоторые останки были также найдены в Вайоминге , Колорадо и Юте , особенно в Национальном парке Каньонлендс и Национальном парке Зайон . [31] Этозавры также известны из супергруппы Ньюарка вдоль восточного побережья Соединенных Штатов, в таких штатах, как Коннектикут и Северная Каролина . [32] Этозавр и паратипоторакс были обнаружены в формации Флеминг-фьорд на Земле Джеймсон , на восточном побережье Гренландии . [33] Эти два рода были первоначально названы из Нижнего Штубенсандштайна в Германии. Шотландия , Польша и Италия также произвели окаменелости этозавра. Несколько этозавров известны из Северной Африки, причем щитки были обнаружены в позднекарнийской формации Таймсгадиуин в Марокко. [34] [ 35] В позднем триасе Марокко находилось в непосредственной близости от Ньюаркской супергруппы Северной Америки на суперконтиненте Пангея . Также возможно, что Desmatosuchus присутствовал в Африке, поскольку к этому роду относят окаменелости из серии Zarzaitine в Алжире . [14]

Этозавры также встречаются в районах, соответствующих Гондване (южная Пангея), хотя они значительно менее распространены и разнообразны, чем на северных континентах. Южноамериканские этозавры известны из Аргентины , Бразилии и Чили . В Аргентине Aetosauroides и Neoaetosauroides происходят из карнийской формации Ишигуаласто и норианской формации Лос-Колорадос соответственно. [36] [1] В Бразилии окаменелости были найдены в формациях Санта-Мария и Катуррита в Риу-Гранди-ду-Сул (Палеоррота). [14] [36] Чилийские этозавры представлены одним фрагментарным родом, Чилинозухами , из региона Антофагаста . [37] Этозавры были также найдены в Индии , которая, наряду с Южной Америкой, была частью Гондваны в позднем триасе. Ранние описания индийских этозавров были основаны на материале из формации Малери на юге центральной Индии, хотя эти останки были слишком недостаточными, чтобы можно было отнести экземпляры к какому-либо конкретному роду. Судя по опубликованным описаниям, окаменелости индийских этозавров наиболее близки к Longosurus и Paratypothorax, которые значительно более специализированы, чем большинство описанных гондванских этозавров. [38] Сообщения об этозаврах с Мадагаскара [39] [40] основаны на вероятных щитках крокодиломорфов . [41] Предполагаемые следы этозавров и скелетный материал из Южной Африки также необоснованны. [42]
Следы, принадлежащие к ихногену Brachychirotherium, часто ассоциируются с этозаврами. [43] [44] Brachychirotherium известны из Риу-Гранди-ду-Сул в Палеорроте, Бразилия [36] , а также в Италии , [45] Германии , [46] на востоке США, [47] и во многих других местах обитания этозавров. Они также распространены на юго-западе США: их нашли в Национальном парке Каньонлендс и Национальном памятнике динозавров . [31] Многие из этих гусениц имеют узкую колею (это означает, что левый и правый отпечатки расположены близко друг к другу) и почти переходят друг друга. Функциональный анализ скелета Typothorax coccinarum, проведенный в 2011 году , показал, что он обладал диапазоном движений, необходимым для создания следов. [48]
Этозавры принадлежат к Pseudosuchia , кладе архозавров, включающей ныне живущих крокодилов и характеризующейся своеобразным строением костей лодыжки . [ сомнительно ] Этозавров традиционно относили к (ныне устаревшей) группе, называемой текодонтами , в которую входили все « примитивные » родственники крокодилов, жившие в триасе. С развитием филогенетики этозавры были позже помещены в группу под названием Сухая , в которую входили многие триасовые круротарзаны, а также более поздние круротарзаны, включая крокодилов. [49]
Первоначально все этоозавры считались членами семейства Stagonolepididae. Ранний филогенетический анализ разделил этозавров на два подсемейства: Aetosaurinae и Desmatosurinae. Этозаврины характеризуются выступами, называемыми возвышениями, на дорсальных парамедиальных остеодермах, расположенных близко к средней линии спины. Десматосухины имеют еще несколько отличительных характеристик, в том числе бороздки на дорсальных парамедианах, которые помогают им плотно прикрепляться к латеральным пластинкам. Многие десматозухины имеют длинные шипы, выступающие из боковых пластинок. Эти шипы особенно заметны у Desmatosuchus . С другой стороны, этозаврины имеют меньше шипов. Многие этозаврины, такие как Aetosaurus и Neoaetosauroides , имеют гладкий панцирь и вообще лишены шипов. [8] Более поздние исследования (см. ниже) отдали предпочтение третьей группе, Typothoracinae, которая, как и Desmatosurinae, имеет длинные шипы, но отличается наличием более остроугольных суставов между остеодермами. Более того, род Aetosauroides теперь часто классифицируется за пределами Stagonolepididae как этозавр, не являющийся стагонолепидидами, в результате чего названия Aetosauria и Stagonolepididae больше не являются синонимами. [50]
Филогения этозавров была впервые исследована в 1994 году палеонтологом Дж. Майклом Пэрришем. В филогенетический анализ были включены Aetosauroides , Aetosaurus , Desmatosuchus , Longosurus , Neoaetosauroides , Stagonolepis и Typothorax . Было обнаружено, что этоозавры образуют кладу с рауизухаями, которую Пэрриш назвал Rauisurchiformes. Rauisuchiformes также включал надотряд Crocodylomorpha , к которому относятся ныне живущие крокодилы . Пэрриш обнаружил, что этозаврия является монофилетической группой и, следовательно, настоящей кладой, состоящей из общего предка этозавра и всех его потомков. Чтобы филогенетически определить этозаврию, Пэрриш выделил пять синапоморфий или общих характеристик. Первая синапоморфия касалась челюсти: предчелюстная кость на кончике была беззубой (беззубой), перевернутой и широкой, образуя «лопату». Кроме того, зубная кость нижней челюсти также беззубая, перевернутая и широкая. Еще одной синапоморфией считали уменьшенный размер и простую коническую форму зубов. Еще две синапоморфии этозавров общие с крокодиломорфами, но не считались признаком тесного филогенетического родства; тело покрыто дорсальным и вентральным панцирем, образующим полный панцирь, а парамедиальные остеодермы намного шире, чем длинны, с характерными ямками. Окончательная синапоморфия обнаружена в строении костей конечностей. У всех этозавров конечности очень крепкие, с крупными мышечными креплениями, такими как дельтопекторальный гребень плечевой кости , четвертый вертел бедренной кости , внутримыщелковый гребень большеберцовой кости и подвздошно-фибулярный вертел малоберцовой кости . [52]
В филогенетическом анализе Пэрриша было обнаружено, что этозавр является самым базальным членом клады, самым ранним, отошедшим от самого недавнего общего предка . После Aetosaurus происходит политомия трех меньших клад, в которой неизвестно, какая клада отделилась от группы первой. В этой политомии был Neoaetosauroides , клада, содержащая Aetosauroides и Stagonolepis , и другая политомия, которая включала Longosurus , Desmatosuchus и кладу, содержащую Paratypothorax и Typothorax .
Более позднее исследование, проведенное палеонтологами Эндрю Б. Хеккертом и Спенсером Г. Лукасом в 1999 году, увеличило число синапоморфий, диагностирующих этозаврию, до 18. Новые синапоморфии включали височные окна или отверстия, которые открывались на боковой стороне черепа, а не на верхней, боковой части черепа. остеодермы, сочленяющиеся с парамедианами, и остеодермы, покрывающие конечности. Aetosaurus по-прежнему оставался наиболее базальным представителем, но филогения более производных этозавров отличалась тем, что Typothorax и Paratypothorax были разделены на две разные клады, а их сестринскими таксонами были Desmatosuchus и Longosurus соответственно. Что еще более важно, в анализ был включен новый этозавр по имени Коахомазух . Коахомазух оказался базальным этозавром, тесно связанным со Stagonolepis , а также появился на ранней стадии в летописи окаменелостей этозавров. Раньше базальные представители были известны только из более поздних времен, произошедших от более продвинутых этозавров. [14]
В 2003 году палеонтологи Саймон Р. Харрис, Дэвид Дж. Гауэр и Марк Уилкинсон изучили предыдущие филогенетические исследования этозавров и раскритиковали то, как они использовали определенные символы для создания кладограмм. Они пришли к выводу, что только три гипотезы о взаимоотношениях этозавров из предыдущих исследований все еще верны: что Aetosaurus является самым базальным этозавром, что Aetosauroides является сестринским таксоном Stagonolepis robertsoni и что Longosurus и Desmatosuchus более тесно связаны друг с другом, чем любой из них. Неоэтозавроиды . Они также исправили деревья из всех предыдущих анализов. [53]
Совсем недавно анализ 2007 года, проведенный палеонтологом Уильямом Дж. Паркером, привел к созданию более крупного дерева филогенетики этозавров с включением Heliocanthus . Основываясь на дереве, Паркер определил клады Typothoracisinae и Paratypothoracisini , обе внутри Aetosaurinae. Паркер также дал пересмотренное филогенетическое определение этозаврии, отметив, что предыдущее определение, данное Хекертом и Лукасом в 2000 году, было несколько двусмысленным. [8] Хеккерт и Лукас (2000) определили Aetosauria как таксон, основанный на стебле , утверждая, что Aetosauria включает в себя все crurotarsans, которые были более тесно связаны с Desmatosuchus , чем с непосредственной сестринской группой Aetosauria. [54] Поскольку непосредственная сестринская группа Aetosauria была неопределенна, Паркер предложил новое определение с несколькими родами круротарсанов, не относящимися к этозаврам, а не с одной сестринской группой. По мнению Паркера, этозаврия включала в себя все таксоны, более тесно связанные с Aetosaurus и Desmatosurus , чем с Leptosurus , Postosurus , Prestosuchus , Poposaurus , Sphenosuchus , Alligator , Gracilisuchus и Revueltosaurus . [8]
В 2012 году был назван новый род этозавров, Aetobarbakinoides . Филогенетический анализ в этом исследовании показал, что Aetosaurinae представляет собой парафилетическую группу. Как парафилетическая группа, этозаврины имели бы общего предка, который также является предком других этозавров, не являющихся этозавринами, и, таким образом, не могли образовать свою собственную кладу . [50] Анализ Паркера 2007 года принял это определение. В 2002 году Хеккерт и Лукас определили Aetosaurinae как « таксон на основе стебля, содержащий все таксоны, более тесно связанные с Aetosaurus, чем с последним общим предком Aetosaurus и Desmatosuchus ». [8] Исследование 2012 года поместило Aetosaurus в основание клады стагонолепидидов, а традиционные таксоны этозаврин заняли последовательно более производные позиции. Согласно анализу, эти таксоны на самом деле более тесно связаны с Desmatosuchus, чем с Aetosaurus . Таким образом, согласно определению Хекерта и Лукаса, Aetosaurinae может ограничиваться только самим Aetosaurus .
Еще одним открытием этого исследования было то, что Aetosauroides находится за пределами Stagonolepididae. Если эта филогения верна, Stagonolepididae и Aetosauria не будут эквивалентными группами, а Aetosauroides будет первым этозавром, не являющимся стагонолепидидами. Следующая кладограмма упрощена после анализа, представленного Девином К. Хоффманом, Эндрю Б. Хеккертом и Линдси Э. Занно. [55]
В 2016 году Уильям Паркер провел новый филогенетический анализ этозавров, предложив альтернативную гипотезу взаимоотношений этозавров. Ниже представлена кладограмма: [4]

Хотя этоозавры известны исключительно из позднего триаса, их нынешнее общепринятое положение в филогении архозавров указывает на то, что они произошли от более базальных псевдозуховых архозавров в раннем или среднем триасе. Учитывая, что этозавры высокоспециализированы и обладают многими анатомическими особенностями, не встречающимися у других псевдозухов, эволюционное происхождение группы плохо изучено. Недавнее открытие полных экземпляров позднетриасового псевдозухия Revueltosaurus callenderi указывает на то, что он, возможно, был близок к предку этозавров. Некоторые филогенетические анализы относят его к сестринскому таксону или ближайшему родственнику этозаврии. Как и этозавры, Revueltosaurus имеет два ряда парамедиальных остеодерм вдоль спины и в щечной области черепа верхнюю челюсть , которая вписывается в борозду скуловой кости . [3] Один филогенетический анализ помещает Turfanosuchus dabanensis , среднетриасовый псевдозухий, в качестве сестринского таксона Revueltosaurus и Aetosauria, что потенциально делает его самым ранним известным « стволовым этозавром» («ствол» означает, что он лежит на ветви, которая включает этозавров, но сам по себе не этозавр). [56] Однако другие исследования считали Turfanosuchus частью другого семейства псевдозухий, грацилизухид . [57]
В 2012 году еще один «стволовой этозавр» был описан из слоев Манда среднего триаса в Танзании. Он отличается от других псевдозухий среднего триаса длинным черепом, небольшим анторбитальным окном , которое вписывается в большую анторбитальную ямку перед глазницей, острыми и изогнутыми зубами и остеодермами, покрывающими большую часть его тела. Подобно этозаврам и ревуэльтозаврам , у него есть верхняя челюсть, которая вписывается в скуловую кость. Revueltosaurus , Turfanosuchus и безымянный танзанийский псевдозух хорошо подходят для предполагаемого предка этозавров, поскольку у них обоих есть двойные ряды листовидных остеодерм вдоль спины, которые потенциально могли эволюционировать в плотно прилегающие парамедиальные остеодермы этозавров. [56]
В 1904 году американский палеонтолог Генри Фэйрфилд Осборн описал этозавров как хищных водных животных отряда Parasuria, отметив, что «[Parasuchia] представляет собой независимый отряд, вероятно, пресноводный, прибрежный, плотоядный, с короткой мордой (Aetosaurus) или длинной мордой (Phytosaurus, Mystriosuchus). формы, аналогичные по своим повадкам современным крокодилам». [58] Ранние останки этозавров часто находили в глине рядом со скелетами водных животных, таких как фитозавры, и наземных животных, таких как динозавры и трилофозавры . Возможно, это заставило некоторых палеонтологов поверить в то, что животные погибли в болотистой местности. Поскольку в этих глинах находилось большое количество скелетов животных, которые обычно не обитали бы в болотах, некоторые палеонтологи даже предположили, что этоозавры собирали трупы животных, которые попали в болота и умерли. Позже возникли сомнения по поводу такого образа жизни, поскольку на зубах этозавров мало признаков плотоядности, а вес доспехов позволяет предположить, что этозавры вели «пассивный образ жизни». Однако до середины 20 века этозавры по-прежнему считались частично водными животными. [28]

Этозавры были травоядными животными, вероятно, питавшимися папоротниками и семенными папоротниками , которые были распространены в триасе. Перевернутая форма их морды предполагает, что этоозавры, возможно, выкапывали корни и клубни. [59] У этозавров есть несколько анатомических особенностей, которые, возможно, были адаптацией к рытью, в том числе короткий радиус относительно плечевой кости (что наблюдается у многих других роющих четвероногих) и большой дельтопекторальный гребень на плечевой кости, служивший местом прикрепления мышц. У этозавров также были большие задние лапы, или стопы , с большими когтями, которые, вероятно, использовались для рытья царапин. У одного этозавра, Typothorax , на плечевой кости имеется энтепикондиль, который является источником пронатора предплечья и мышц-сгибателей рук, часто используемых при рытье. Кроме того, на локтевой кости имеется большой локтевой отросток , который выступает назад за пределы локтя, обеспечивая большую площадь для прикрепления трехглавой мышцы. [44] Хотя многие исследования показали, что этозавры вели роющий или роющий образ жизни, [52] у этозавров мало характеристик, которые есть у ископаемых животных в качестве приспособлений к рытью. Таким образом, вполне вероятно, что этоозавры были способны в некоторой степени копать, возможно, ища пищу, но не могли рыть норы. [44]
Хотя особенности конечностей указывают на то, что этоозавры, вероятно, копали пищу, особенности черепа и зубов могут указывать на то, какую пищу они ели. У этозавров есть множество производных особенностей, отсутствующих у других круротарсанов, что указывает на то, что они адаптированы к другой диете. В отличие от острых, загнутых зубов других архозавров триаса, у этозавров были простые конические зубы. Кончики челюстей были беззубыми или беззубыми и, вероятно, поддерживали клюв. Зубы изношены очень незначительно, что позволяет предположить, что этозавры не потребляли жесткий и жесткий растительный материал. Более вероятно, что они потребляли неабразивную растительность, например, мягкие листья. [52]
Были предложены альтернативные теории питания этозавров. В 1947 году Х. Дж. Савин предположил, что этозавр Longosurus был падальщиком, основываясь на непосредственной близости некоторых экземпляров к большому количеству скелетов, которые, вероятно, были тушами. [28] Исследование биомеханики челюстей южноамериканского рода Neoaetosauroides , проведенное в 2009 году , показало, что животное, возможно, питалось личинками и насекомыми без твердых экзоскелетов. Это связано с тем, что у Neoaetosauroides отсутствуют зазубрины или изнашиваемые фасетки на зубах, а рычаг челюсти не рассчитан на сильные силы, такие как дробление и измельчение. Исследование показало, что у северных этозавров, таких как Desmatosuchus и Stagonolepis, действительно были челюсти, которые поддерживали сильную мускулатуру и, вероятно, лучше подходили для поедания растительного материала. [60] [61]

По крайней мере, некоторые этоозавры строили гнезда и защищали свои яйца. [62]
В 1996 году геолог Стивен Хасиотис обнаружил окаменелые чашеобразные ямы возрастом 220 миллионов лет в Окаменелом лесу Аризоны, в части формации Чинл , которые, как предполагается, были гнездами этозавров и фитозавров . «Гнезда» уплотнены и очень похожи на гнезда современных крокодилов , охраняющих свои гнезда. [59] Однако кажется, что эти «гнезда» являются результатом выветривания песчаника. [63]
Второе возможное место гнездования этозавров известно на северо-востоке Италии. Гнезда сохранились в виде углублений в карбонатных породах круглой или подковообразной формы с высокими гребнями по бокам. Они кажутся необычайно сложными для гнезд, созданных триасовыми рептилиями. Рядом были обнаружены следы архозавров, напоминающие этозавров, хотя они не находились в том же слое. Поскольку следы были найдены так близко к гнездам, вполне вероятно, что их построили этозавры. [62]
Возраст отдельных этозавров можно определить, исследуя их остеодермы. Утверждалось, что некоторые изолированные остеодермы принадлежат юным этозаврам на основании их размера и формы, но эти гипотезы часто подвергались сомнению. Например, юные остеодермы Calyptosuchus позже были идентифицированы как таковые у псевдозухия Revueltosaurus с небольшим телом (который не является этозавром), а юные остеодермы Desmatosuchus были переинтерпретированы как остеодермы этозавра Acaenasuchus , который в зрелом возрасте имел относительно небольшой размер тела. . Исследования костной структуры парамедиальных остеодерм показывают, что новая кость откладывалась по краям каждой пластинки в течение жизни этозавра. Это означает, что линии остановки роста на нижней стороне парамедиальных остеодерм можно использовать для определения возраста человека. [3] Сравнение возраста отдельных особей с их общей длиной тела показывает, что этозавры увеличивались в длину с относительно постоянной скоростью, но увеличивались в массе тела с разной скоростью в зависимости от того, были ли у них широкие тела, как Typothorax , или узкие тела, как Aetobarbakinoides . Этоозавры также, по-видимому, росли медленнее, чем современные крокодилы. [64] Анализ костей конечностей этозавров показывает, что они росли быстро в молодом возрасте и медленнее во взрослом возрасте. Такая модель роста наблюдается у большинства других псевдозухов. [3]
Поскольку виды этозавров обычно имеют ограниченный ареал окаменелостей и многочисленны в слоях, в которых они обитают, они полезны в биохронологии . Остеодермы являются наиболее распространенными останками, связанными с этозаврами, поэтому по одному опознаваемому щитку можно точно датировать слой, в котором он обнаружен.
Один этозавр, Typothorax coccinarum , был использован для определения ревуэльтской фауны наземных позвоночных . Фауны наземных позвоночных (LVF) — это временной интервал, который определяется датой первого появления (FAD) или первым появлением окаменелости- индекса четвероногих и обычно используется для датировки наземных слоев позднего триаса и ранней юры . [67] [68] Поскольку FAD T. coccinarum находится в начале норийской стадии , ревуэльтская LVF начинается в начале норийского периода около 216 миллионов лет назад . Ревуэльтский период заканчивается следующим FAD, которым оказался фитозавр Redondasaurus и начало апачского LVF. [2] [69]
Биохроны родов этозавров были разработаны для датирования слоев группы Чинл на юго-западе США. Из группы Чинле известно до 13 родов этозавров, большинство из которых встречаются в разных местах и в течение коротких периодов времени. В 1996 году палеонтологи Спенсер Г. Лукас и Эндрю Б. Хекерт выделили пять биохронов, основанных на присутствии этозавров по всей группе Чинл. [70] Число биохронов выросло до 11 в исследовании 2007 года, проведенном Хеккертом и Лукасом вместе с Адрианом П. Хантом и Джастином А. Спилманном. Эти биохроны произошли от отишалкского LVF до апачского LVF и включали такие роды, как Longosurus , Tecovasuchus и Typothorax . [66]
В 2007 году палеонтологи из Музея естественной истории и науки Нью-Мексико в Альбукерке, штат Нью-Мексико, были обвинены в плагиате в некоторых опубликованных ими статьях, посвященных этозаврам. В декабре 2006 года в бюллетене музея в качестве замены названия «Desmatosuchus» chamaensis был указан род Rioarribasuchus . [71] Однако четырьмя годами ранее палеонтолог Уильям Паркер переназначил букву «D». chamaensis к недавно названному роду Heliocanthus в неопубликованной диссертации, которая получила широкое распространение среди исследователей этозавров. [72] Поскольку название не было официально опубликовано, оно считалось nomen nudum до января 2007 года, когда описание Гелиоканта Паркером было опубликовано в Журнале систематической палеонтологии . [8] Авторы статьи 2006 года, Спенсер Г. Лукас, Адриан П. Хант и Джастин А. Спилманн, были обвинены в «интеллектуальной краже» палеонтологами Джеффом Марцем, Майком Тейлором , Мэттом Веделем и Дарреном Нэйшем , которые утверждали, что что Лукас и др. знали, что Паркер в конечном итоге переопишет этот вид и формально выделит новый род. По словам Марца, Тейлора, Веделя и Нэйша, авторы поспешили опубликовать свое имя до того, как Паркер успел опубликовать свое. [73] В то время Лукас, Хант и Шпильман входили в редакционную коллегию бюллетеня NMMNHS, а Хант также был директором музея.
Совпадающий спор возник после того, как Шпильман, Хант и Лукас опубликовали в 2006 году статью, в которой упоминалось, что голотип Redondasuchus был не левым парамедианом , а правым. [74] В 2002 году Джефф Марц пришел к такому же выводу в неопубликованной диссертации. [75] Он, вместе с Тейлором, Веделем и Нэйшем, утверждал, что это была еще одна форма плагиата, поскольку диссертация Марца 2002 года была процитирована Шпильманном и др. (2006), даже несмотря на то, что его заключение о Redondasurus не упоминалось. [73]
Обвинения были отклонены на внутреннем заседании NMMNHS, о котором сообщил Лукас в 2008 году. [76] [77] [78] Они также были доведены до сведения независимой стороны, Комитета по этическому образованию Общества палеонтологии позвоночных (SVP). в 2007 году, а ответ был дан в 2008 году. Что касается Redondasurus , СВП не нашел явных доказательств плагиата. В случае с Heliocanthus и Rioarribasuchus СВП не пытался решить проблему, поскольку Лукас и др. и Паркер предложили противоречивые версии относительно общения и намерений. Ответ СВП завершился обновлением его этической политики и рекомендациями о том, как лучше разрешать подобные разногласия в будущем. [79] [80] [81] Вся полемика стала известна как «Этогейт», в связи со знаменитым Уотергейтским скандалом 1970-х годов. [73] Это привлекло широкое внимание местных газет Альбукерке и научных блогов. [82] Этому также была посвящена новостная статья в выпуске журнала Nature за 2008 год . [83]
{{cite journal}}: Требуется цитировать журнал |journal=( помощь )