Dictyostelium discoideum — это вид почвенных амеб, принадлежащих к типу Amoebozoa , infraphylum Mycetozoa . Обычно называемый слизистой плесенью , D. discoideum — это эукариот , который в течение своей жизни переходит из группы одноклеточных амеб в многоклеточную слизь, а затем в плодовое тело. Его уникальный бесполый жизненный цикл состоит из четырех стадий: вегетативная, агрегация, миграция и кульминация. Жизненный цикл D. discoideum относительно короткий, что позволяет своевременно просматривать все стадии. Клетки, участвующие в жизненном цикле, подвергаются движению, химической сигнализации и развитию, что применимо к исследованиям рака у человека. Простота его жизненного цикла делает D. discoideum ценным модельным организмом для изучения генетических, клеточных и биохимических процессов в других организмах. [2]
В дикой природе D. discoideum можно найти в почве и влажной опавшей листве. Его основной рацион состоит из бактерий , таких как Escherichia coli , которые находятся в почве и разлагающихся органических веществах. Одноядерные амебы D. discoideum потребляют бактерии, которые находятся в их естественной среде обитания, которая включает почву лиственных лесов и разлагающиеся листья. [3]
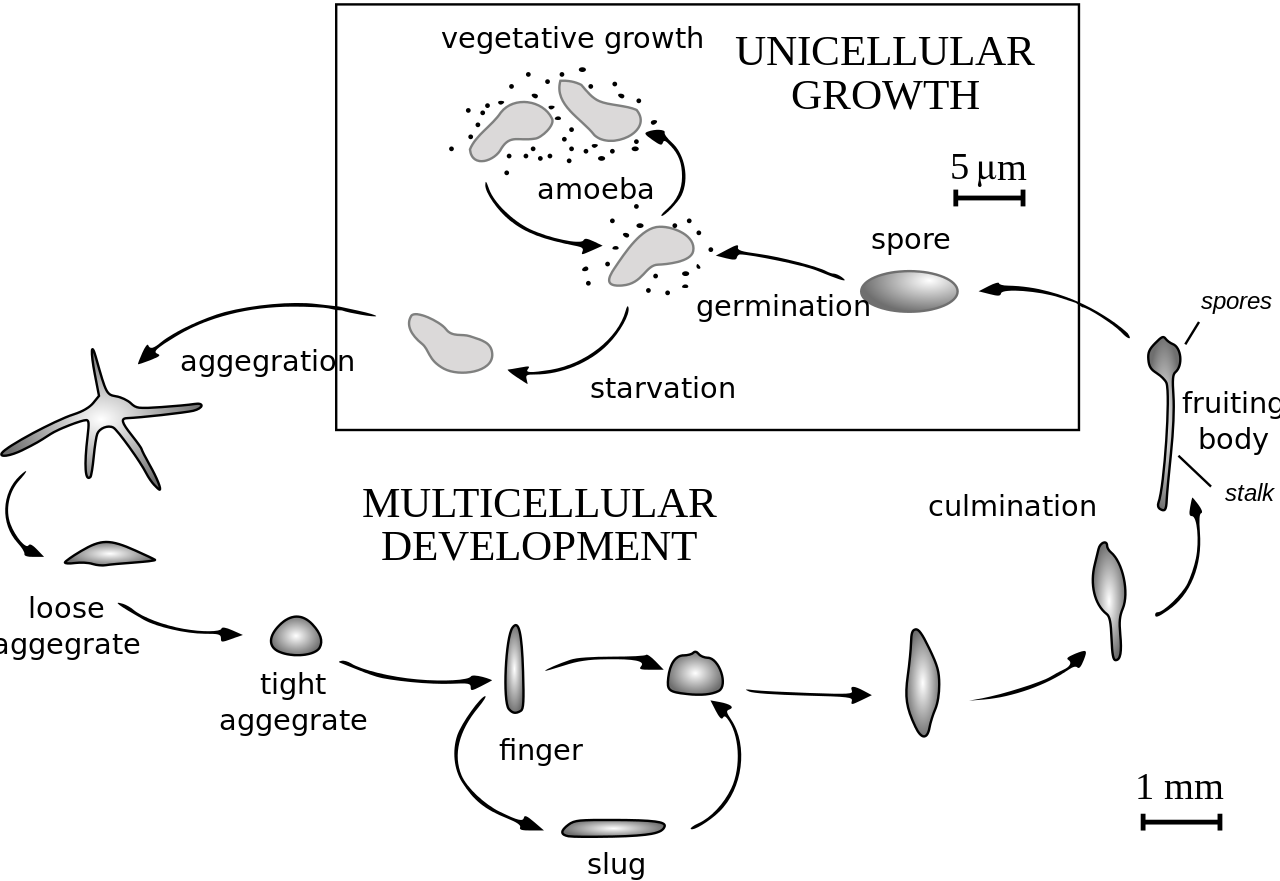
Жизненный цикл D. discoideum начинается , когда споры высвобождаются из зрелого сорокарпия (плодового тела). Миксамебы вылупляются из спор в теплых и влажных условиях. Во время вегетативной стадии миксамебы делятся митозом , питаясь бактериями. Бактерии выделяют фолиевую кислоту , которая привлекает миксамебы. Когда запас бактерий истощается, миксамебы переходят в стадию агрегации.
Во время агрегации голодание инициирует производство белковых соединений, таких как гликопротеины и аденилатциклаза . [4] Гликопротеины обеспечивают межклеточную адгезию , а аденилатциклаза создает циклический АМФ . Циклический АМФ выделяется амебами для привлечения соседних клеток в центральное местоположение. По мере того, как они движутся к сигналу, они сталкиваются друг с другом и склеиваются с помощью молекул адгезии гликопротеинов.
Стадия миграции начинается, как только амебы образуют плотный агрегат, и удлиненная куча клеток опрокидывается, чтобы лечь на землю. Амебы работают вместе как подвижный псевдоплазмодий, также известный как слизняк. Слизняк имеет длину около 2–4 мм, состоит из до 100 000 клеток [5] и способен двигаться, производя целлюлозную оболочку в своих передних клетках, через которую движется слизняк. [6] Часть этой оболочки остается позади в виде скользкого следа, поскольку он движется к аттрактантам, таким как свет , тепло и влажность, в направлении только вперед. [6] Циклический АМФ и вещество, называемое фактором, индуцирующим дифференциацию , помогают формировать различные типы клеток. [6] Слизняк дифференцируется на престебельные и преспоровые клетки, которые движутся к переднему и заднему концам соответственно. Как только слизень находит подходящую среду, передний конец слизняка образует стебель плодового тела, а задний конец образует споры плодового тела. [6] Переднеподобные клетки, которые были обнаружены совсем недавно, также рассеяны по всей задней области слизняка. Эти переднеподобные клетки образуют самое дно плодового тела и шляпки спор. [6] После того, как слизень оседает в одном месте, задний конец расправляется, а передний конец поднимается в воздух, образуя то, что называется «мексиканской шляпой», и начинается стадия кульминации.
Клетки предстебелька и клетки предспоры меняются местами на стадии кульминации, образуя зрелое плодовое тело. [6] Передний конец мексиканской шляпы образует целлюлозную трубку, которая позволяет более задним клеткам перемещаться вверх по внешней стороне трубки к вершине, а клетки предстебелька перемещаются вниз. [6] Эта перестройка формирует стебель плодового тела, состоящий из клеток переднего конца слизняка, а клетки заднего конца слизняка находятся наверху и теперь образуют споры плодового тела. В конце этого 8–10-часового процесса зрелое плодовое тело полностью формируется. [6] Это плодовое тело имеет высоту 1–2 мм и теперь способно начать весь цикл заново, высвобождая зрелые споры, которые становятся миксамебами.
Хотя D. discoideum обычно размножается бесполым путем , D. discoideum все еще способен к половому размножению, если соблюдены определенные условия. У D. discoideum есть три различных типа спаривания, и исследования определили половой локус, который определяет эти три типа спаривания. Штаммы типа I определяются геном, называемым MatA, штаммы типа II имеют три различных гена: MatB (гомологичный Mat A), Mat C и Mat D, а штаммы типа III имеют гены Mat S и Mat T (гомологичные Mat C и Mat D). [7] Эти полы могут спариваться только с двумя различными полами, но не со своим собственным. [7]
При инкубации с бактериальной пищей может происходить гетероталличное или гомоталличное половое развитие, что приводит к образованию диплоидной зиготы. [8] [9] Гетероталлическое спаривание происходит, когда две амебы разных типов спаривания находятся в темной и влажной среде, где они могут слиться во время агрегации, образуя гигантскую клетку зиготы. Затем гигантская клетка высвобождает цАМФ, чтобы привлечь другие клетки, а затем поглощает другие клетки каннибалистически в агрегате. Потребленные клетки служат для заключения всего агрегата в толстую целлюлозную стенку, чтобы защитить его. Это известно как макроциста . Внутри макроцисты гигантская клетка сначала делится посредством мейоза , затем посредством митоза, чтобы произвести множество гаплоидных амеб, которые будут выпущены для питания, как это делают обычные амебы. Гомоталлические штаммы D. discoideum AC4 и ZA3A также способны производить макроцисты. [10] Каждый из этих штаммов, в отличие от гетероталличных штаммов, вероятно, выражает оба аллеля типа спаривания ( matA и mata ). Хотя половое размножение возможно, очень редко можно увидеть успешное прорастание макроцисты D. discoideum в лабораторных условиях. Тем не менее, рекомбинация широко распространена в природных популяциях D. discoideum , что указывает на то, что пол, вероятно, является важным аспектом их жизненного цикла. [9]
Поскольку многие из его генов гомологичны человеческим генам, но его жизненный цикл прост, D. discoideum обычно используется в качестве модельного организма . Его можно наблюдать на организменном, клеточном и молекулярном уровнях, прежде всего из-за его ограниченного количества типов клеток и поведения, а также его быстрого роста. [6] Он используется для изучения клеточной дифференциации, хемотаксиса и апоптоза , которые являются нормальными клеточными процессами. Он также используется для изучения других аспектов развития, включая сортировку клеток, формирование паттернов, фагоцитоз, подвижность и передачу сигналов. [11] Эти процессы и аспекты развития либо отсутствуют, либо их слишком сложно увидеть в других модельных организмах. D. discoideum тесно связан с высшими метазоа. Он несет похожие гены и пути, что делает его хорошим кандидатом на нокаут генов . [12]
Процесс дифференциации клеток происходит, когда клетка становится более специализированной, чтобы развиться в многоклеточный организм. Изменения в размере, форме, метаболической активности и отзывчивости могут происходить в результате корректировок в экспрессии генов. Разнообразие клеток и дифференциация у этого вида включают решения, принимаемые на основе межклеточных взаимодействий в путях либо к стебельчатым клеткам, либо к споровым клеткам. [13] Судьбы этих клеток зависят от их среды и формирования паттерна. Таким образом, организм является превосходной моделью для изучения дифференциации клеток.

Хемотаксис определяется как перемещение организма к химическому стимулу или от него по градиенту концентрации химических веществ. Некоторые организмы демонстрируют хемотаксис, когда они движутся к источнику питательных веществ. У D. discoideum амеба выделяет сигнал, цАМФ, из клетки, привлекая другие амебы к миграции к источнику. Каждая амеба движется к центральной амебе, которая выделяет наибольшее количество секреции цАМФ. Секреция цАМФ затем проявляется всеми амебами и является призывом для них начать агрегацию. Эти химические выбросы и движение амеб происходят каждые шесть минут. Амебы движутся к градиенту концентрации в течение 60 секунд и останавливаются, пока не будет отправлена следующая секреция. Такое поведение отдельных клеток имеет тенденцию вызывать колебания в группе клеток, и химические волны различной концентрации цАМФ распространяются через группу по спирали. [14] : 174–175
Элегантный набор математических уравнений, воспроизводящих спирали и потоковые узоры D. discoideum, был открыт математическими биологами Томасом Хёфером и Мартином Бёрлийстом. Математический биолог Корнелис Дж. Вейер доказал, что подобные уравнения могут моделировать его движение. Уравнения этих узоров в основном зависят от плотности популяции амеб, скорости выработки циклического АМФ и чувствительности отдельных амеб к циклическому АМФ. Спиральный узор формируется амебами в центре колонии, которые вращаются, посылая волны циклического АМФ. [15] [16]
Использование цАМФ в качестве хемотаксического агента не установлено ни в одном другом организме. В биологии развития это один из понятных примеров хемотаксиса, который важен для понимания воспаления человека, артрита, астмы, перемещения лимфоцитов и аксонного руководства. Фагоцитоз используется в иммунном надзоре и презентации антигенов, в то время как определение типа клеток, сортировка клеток и формирование паттерна являются основными особенностями эмбриогенеза, которые можно изучать с помощью этих организмов. [6]
Однако следует отметить, что колебания цАМФ могут не быть необходимыми для коллективной миграции клеток на многоклеточных стадиях. Исследование показало, что опосредованная цАМФ сигнализация изменяется от распространяющихся волн к устойчивому состоянию на многоклеточной стадии D. discoideum. [17]
Термотаксис — это движение по градиенту температуры. Было показано, что слизни мигрируют по крайне пологим градиентам всего 0,05 °C/см, но выбранное направление сложное; оно, по-видимому, находится вдали от температуры примерно на 2 °C ниже температуры, к которой они акклиматизировались. Это сложное поведение было проанализировано с помощью компьютерного моделирования поведения и периодической картины изменений температуры в почве, вызванных ежедневными изменениями температуры воздуха. Вывод заключается в том, что поведение перемещает слизней на несколько сантиметров ниже поверхности почвы к поверхности. Это удивительно сложное поведение примитивного организма, не имеющего явного чувства гравитации. [14] : 108–109
Апоптоз (запрограммированная гибель клеток) является нормальной частью развития видов. [4] Апоптоз необходим для правильного расположения и формирования сложных органов. Около 20% клеток D. discoideum альтруистически жертвуют собой при формировании зрелого плодового тела. На стадии псевдоплазмодия (слизняка или грекса) своего жизненного цикла организм сформировал три основных типа клеток: предстебельные, предспоровые и переднеподобные клетки. Во время кульминации предстебельные клетки выделяют целлюлозную оболочку и вытягиваются в виде трубки через грекс. [4] По мере дифференциации они образуют вакуоли и увеличиваются, поднимая преспоровые клетки. Стебельковые клетки подвергаются апоптозу и умирают, когда преспоровые клетки поднимаются высоко над субстратом. Затем преспоровые клетки становятся споровыми клетками, каждая из которых становится новой миксамебой при рассеивании. [6] Это пример того, как апоптоз используется в формировании репродуктивного органа — зрелого плодового тела.
Недавний крупный вклад в исследования Dictyostelium был получен благодаря новым методам, позволяющим визуализировать активность отдельных генов в живых клетках. [18] Это показало, что транскрипция происходит «всплесками» или «импульсами» ( транскрипционный всплеск ), а не следует простому вероятностному или непрерывному поведению. Всплеск транскрипции теперь, по-видимому, сохраняется у бактерий и людей. Еще одной замечательной особенностью организма является то, что у него есть наборы ферментов репарации ДНК, обнаруженных в клетках человека, которые отсутствуют во многих других популярных модельных системах метазойных организмов. [19] Дефекты в репарации ДНК приводят к разрушительным человеческим ракам, поэтому возможность изучать человеческие белки репарации в простой поддающейся обработке модели окажется бесценной.
Способность этого организма легко изолироваться и культивироваться в лабораторных условиях [6] [20] добавляет ему привлекательности в качестве модельного организма. Хотя D. discoideum можно выращивать в жидкой культуре, обычно его выращивают в чашках Петри, содержащих питательный агар , а поверхности поддерживаются влажными. Культуры лучше всего растут при температуре 22–24 °C (комнатная температура). D. discoideum питается в основном E. coli , что достаточно для всех стадий жизненного цикла. Когда поступление пищи уменьшается, миксамебы объединяются, образуя псевдоплазмодии. Вскоре чашка покрывается различными стадиями жизненного цикла. Проверка чашки часто позволяет проводить подробные наблюдения за развитием. Клетки можно собирать на любой стадии развития и быстро выращивать.
При выращивании D. discoideum в лабораторных условиях важно учитывать его поведенческие реакции. Например, он имеет сродство к свету, высоким температурам, высокой влажности, низким ионным концентрациям и кислой стороне градиента pH. Часто проводятся эксперименты, чтобы увидеть, как манипуляции этими параметрами затрудняют, останавливают или ускоряют развитие. Изменения этих параметров могут изменить скорость и жизнеспособность роста культуры. Кроме того, плодовые тела, поскольку это самая высокая стадия развития, очень чувствительны к потокам воздуха и физическим стимулам. Неизвестно, есть ли стимул, связанный с высвобождением спор.
Подробный анализ экспрессии белка у Dictyostelium был затруднен из-за больших сдвигов в профиле экспрессии белка между различными стадиями развития и общей нехватки коммерчески доступных антител к антигенам Dictyostelium . [21] В 2013 году группа из онкологического центра Beatson West of Scotland сообщила о стандарте визуализации белка без антител для иммуноблоттинга, основанном на обнаружении MCCC1 с использованием конъюгатов стрептавидина . [22]
Бактериальный род Legionella включает виды, вызывающие болезнь легионеров у людей. D. discoideum также является хозяином для Legionella и представляет собой подходящую модель для изучения процесса заражения. [23] В частности, D. discoideum разделяет с клетками-хозяевами млекопитающих схожий цитоскелет и клеточные процессы, имеющие отношение к заражению Legionella , включая фагоцитоз , мембранный транспорт, эндоцитоз , сортировку везикул и хемотаксис.
В отчете журнала Nature за 2011 год были опубликованы результаты, демонстрирующие «примитивное фермерское поведение» в колониях D. discoideum . [24] [25] Описанный как « симбиоз » между D. discoideum и бактериальной добычей, около трети собранных в дикой природе колоний D. discoideum занимались « разведением » бактерий, когда бактерии были включены в плодовые тела слизевика. [25] Включение бактерий в плодовые тела позволяет «засевать» источник пищи в месте распространения спор, что особенно ценно, если новое место имеет мало пищевых ресурсов. [25] Колонии, полученные из «фермерских» спор, обычно также демонстрируют такое же поведение при споруляции. Это включение имеет свою стоимость: те колонии, которые не потребляют все бактерии-жертвы, производят более мелкие споры, которые не могут распространяться так широко. Кроме того, гораздо меньше пользы для спор, содержащих бактерии, которые приземляются в богатом пищей регионе. Этот баланс издержек и выгод поведения может способствовать тому, что меньшинство колоний D. discoideum практикует эту практику. [24] [25]
Известно, что D. discoideum поедает как грамположительные , так и грамотрицательные бактерии , но некоторые из фагоцитированных бактерий, включая некоторые патогены человека, [26] способны жить в амебах и выходить, не убивая клетку. Когда они попадают в клетку, где они находятся и когда они покидают клетку, неизвестно. Исследование еще не окончательное, но можно нарисовать общий жизненный цикл D. discoideum, адаптированный для фермерских клонов, чтобы лучше понять этот симбиотический процесс.

На рисунке можно увидеть различные стадии. Сначала, на стадии голодания, бактерии заключены в D. discoideum , [26] после попадания в амебу, в фагосоме слияние с лизосомами блокируется, и эти незрелые фагосомы окружены органеллами клетки хозяина, такими как митохондрии, везикулы и многослойная мембрана, полученная из шероховатого эндоплазматического ретикулума (RER) амеб. Роль RER во внутриклеточной инфекции неизвестна, но RER не требуется как источник белков для бактерий. [27] Бактерии находятся внутри этих фагосом во время стадий агрегации и многоклеточного развития. Амебы сохраняют свою индивидуальность, и у каждой амебы есть своя собственная бактерия. На кульминационной стадии, когда образуются споры, бактерии переходят из клетки в сорус с помощью цитоскелетной структуры, которая предотвращает разрушение клетки хозяина. [28] Некоторые результаты показывают, что бактерии используют экзоцитоз, не убивая клетку. [28] Свободноживущие амебы, по-видимому, играют решающую роль в сохранении и распространении некоторых патогенов в окружающей среде. Временная ассоциация с амебами была зарегистрирована для ряда различных бактерий, включая Legionella pneumophila , многие виды Mycobacterium , Francisella tularensis и Escherichia coli , среди прочих. [27] Сельское хозяйство, по-видимому, играет решающую роль в выживании патогенов, поскольку они могут жить и размножаться внутри D. discoideum , что делает земледелие. Отчет Nature сделал важный шаг вперед в познании поведения амеб, и известная испанская фраза, переводимая как «ты глупее амебы», теряет смысл, поскольку амебы являются прекрасным примером социального поведения с удивительной координацией и чувством жертвенности ради блага вида. [ необходима цитата ]
Клетки-сторожа у Dictyostelium discoideum являются фагоцитарными клетками, ответственными за удаление токсичных материалов из стадии слизняка социального цикла. Обычно круглые по форме, эти клетки присутствуют внутри оболочки слизняка, где они, как обнаружено, свободно циркулируют. Процесс детоксикации происходит, когда эти клетки поглощают токсины и патогены внутри слизняка посредством фагоцитоза . Затем клетки слипаются в группы по пять-десять клеток, которые затем прикрепляются к внутренней оболочке слизняка. Оболочка отшелушивается, когда слизень мигрирует на новое место в поисках пищевых бактерий.
Дозорные клетки составляют приблизительно 1% от общего числа клеток слизняков, и число дозорных клеток остается постоянным даже по мере их высвобождения. Это указывает на постоянную регенерацию дозорных клеток внутри слизней, поскольку они удаляются вместе с токсинами и патогенами. Дозорные клетки присутствуют в слизняке даже тогда, когда нет токсинов или патогенов, которые нужно удалить. Дозорные клетки были обнаружены у пяти других видов Dictyostelia , что предполагает, что дозорные клетки можно описать как общую характеристику врожденной иммунной системы у социальных амеб. [29]
Количество клеток-дозорных варьируется в зависимости от статуса выращивания дикого D. discoideum . При воздействии токсичной среды, созданной с помощью бромистого этидия, было показано, что количество клеток-дозорных на миллиметр было ниже у фермеров, чем у нефермеров. Этот вывод был сделан путем наблюдения за следами, оставленными при миграции слизней, и подсчета количества клеток-дозорных на миллиметр. Однако количество клеток-дозорных не влияет на производство и жизнеспособность спор у фермеров. Фермеры, подвергавшиеся воздействию токсичной среды, производят такое же количество спор, как и фермеры в нетоксичной среде, а жизнеспособность спор была одинаковой у фермеров и нефермеров. Когда клада 2 Burkholderia или бактерии, связанные с фермерами, удаляются у фермеров, производство и жизнеспособность спор были аналогичны таковым у нефермеров. Таким образом, предполагается, что бактерии, переносимые фермерами, выполняют дополнительную функцию защиты фермеров от потенциального вреда, вызванного токсинами или патогенами. [30]
В более старых классификациях Dictyostelium был помещен в несуществующий полифилетический класс Acrasiomycetes . Это был класс клеточных слизевиков, который характеризовался агрегацией отдельных амеб в многоклеточное плодовое тело, что делало его важным фактором, связывающим акразиды с диктиостелидами. [31]
Более поздние геномные исследования показали, что Dictyostelium сохранил больше своего предкового геномного разнообразия, чем растения и животные, хотя филогения на основе протеома подтверждает, что амебозои отделились от линии животных и грибов после разделения растений и животных. [32] Подкласс Dictyosteliidae, отряд Dictyosteliales представляет собой монофилетическую группу в пределах Mycetozoa, группы, которая включает протостелид, диктиостелид и миксогастридных слизевиков. Анализ данных фактора удлинения-1α (EF-1α) подтверждает, что Mycetozoa является монофилетической группой, хотя деревья рРНК помещают ее в полифилетическую группу. Кроме того, эти данные подтверждают идею о том, что диктиостелид и миксогастриды более тесно связаны друг с другом, чем с протостелидами. Анализ EF-1α также поместил Mycetozoa в качестве непосредственной внешней группы для клады животных и грибов. [33] Последние филогенетические данные помещают диктиостелид прочно в супергруппу Amoebozoa , вместе с миксомицетами . Между тем, протостелиды оказались полифилетическими, их стебельчатые плодовые тела являются конвергентной чертой нескольких неродственных линий. [34]
Проект по секвенированию генома D. discoideum был завершен и опубликован в 2005 году международным сотрудничеством институтов. Это был первый полностью секвенированный геном свободноживущего простейшего. D. discoideum состоит из гаплоидного генома размером 34 Мб с базовым составом 77% [A+T] и содержит шесть хромосом, которые кодируют около 12 500 белков. [3] Секвенирование генома D. discoideum обеспечивает более сложное исследование его клеточной и онтогенетической биологии.
Тандемные повторы тринуклеотидов очень распространены в этом геноме; один класс генома является кластеризованным, что привело исследователей к мысли, что он служит центромерами. Повторы соответствуют повторяющимся последовательностям аминокислот и, как полагают, расширяются за счет расширения нуклеотидов. [3] Расширение тринуклеотидных повторов также происходит у людей, что в целом приводит ко многим заболеваниям. Изучение того, как клетки D. discoideum переносят эти повторы аминокислот, может дать представление о том, как люди могут переносить их.
Каждый секвенированный геном играет важную роль в идентификации генов, которые были приобретены или утеряны с течением времени. Сравнительные геномные исследования позволяют сравнивать эукариотические геномы. Филогения, основанная на протеоме, показала, что амебозои отклонились от линии животных и грибов после разделения растений и животных. [3] Геном D. discoideum примечателен тем, что многие его кодируемые белки обычно встречаются у грибов, растений и животных. [3]
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка ){{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )