В археогенетике западный охотник-собиратель ( WHG , также известный как западноевропейский охотник-собиратель , западноевропейский охотник-собиратель или кластер Оберкасселя ) ( ок. 15 000–5 000 лет назад ) является отчетливым наследственным компонентом современных европейцев, представляющим происхождение от населения . мезолитических охотников-собирателей , рассеявшихся по западной , южной и центральной Европе , от Британских островов на западе до Карпат на востоке, после отступления ледникового покрова Последнего ледникового максимума . [2] Это тесно связано, а иногда и считается синонимом концепции кластера Виллабруна , названного в честь пещеры Рипари Виллабруна в Италии, известной из конечного плейстоцена Европы, которая в значительной степени является предком более поздних популяций WHG.
WHG имеют более близкое генетическое родство с древними и современными народами Ближнего Востока и Кавказа, чем более ранние европейские охотники-собиратели. Считается, что WHG мигрировали в Европу примерно в конце последнего ледникового максимума , распространившись по Западной Европе в конце плейстоцена, в значительной степени вытеснив более ранние группы кроманьонцев, такие как мадленцы .
Наряду со скандинавскими охотниками-собирателями (SHG) и восточными охотниками-собирателями (EHG), WHG составляли одну из трех основных генетических групп в послеледниковый период ранней голоценовой Европы. [3] Граница между WHG и EHG проходила примерно от нижнего Дуная , на север вдоль западных лесов Днепра к западной части Балтийского моря . [2]
ГСП, в свою очередь, представляли собой почти равное сочетание WHG и EHG. Когда-то WHG были основным населением всей Европы, но были в значительной степени вытеснены последовательным расширением ранних европейских фермеров (EEF) в раннем неолите , но пережили возрождение во время среднего неолита. В период позднего неолита и раннего бронзового века западные степные пастухи (WSH) из Понтийско-Каспийской степи начали массовую экспансию, которая еще больше вытеснила WHG. Среди современного населения происхождение WHG наиболее распространено среди населения восточно-Балтийского региона. [4]
Западные охотники-собиратели (WHG) признаны отдельным предковым компонентом, вносящим вклад в происхождение большинства современных европейцев . [5] Большинство европейцев можно смоделировать как смесь WHG, EEF и WSH из Понтийско-Каспийской степи . [6] WHG также внесли свой вклад в происхождение других древних групп, таких как ранние европейские фермеры (EEF), которые, однако, были в основном анатолийского происхождения. [5] С распространением неолита EEF стал доминировать в генофонде в большинстве частей Европы, хотя родословная WHG возродилась в Западной Европе от раннего неолита до среднего неолита. [7]
WHGs представляют собой крупный сдвиг населения в Европе в конце ледникового периода, вероятно, расширение населения в континентальную Европу из рефугиумов Юго-Восточной Европы или Западной Азии . [8] Считается, что их предки отделились от восточных евразийцев около 40 000 лет назад, а от древних жителей Северной Евразии (ANE) - до 24 000 лет назад (оценочная дата возраста мальтийского мальчика ). Впоследствии эта дата была перенесена еще дальше во времени благодаря находкам на стоянке рогов носорога Яна , примерно на 38 тыс. лет назад, вскоре после расхождения западно-евразийских и восточно-евразийских линий. [5] [9] Валлини и др. 2022 утверждает, что рассредоточение и раскол западноевразийских линий произошли не ранее ок. 38 000 лет назад, причем более древние европейские образцы начального верхнего палеолита , такие как Златы Кун , Пештера ку Оасе и Бачо Киро , не были связаны с западными охотниками-собирателями, но были ближе к древневосточным евразийцам или были базальными для обоих. [10]
Генетические данные свидетельствуют о том, что предки популяции WHG генетически разошлись с охотниками-собирателями на Ближнем Востоке и на Кавказе около 26 000 лет назад, во время последнего ледникового максимума . [11] Геномы WHG демонстрируют более высокую близость к древним и современным популяциям Ближнего Востока по сравнению с европейцами раннего палеолита, такими как граветты . Близость к древним популяциям Ближнего Востока в Европе увеличилась после последнего ледникового максимума , что коррелирует с расширением происхождения WHG (Виллабруна или Оберкассель). Есть также свидетельства двунаправленного потока генов между популяциями WHG и Ближнего Востока еще 15 000 лет назад. Связанные с WHG останки принадлежали в первую очередь человеческим гаплогруппам Y-хромосомы I-M170 с более низкой частотой C-F3393 (в частности, кладе C-V20/C1a2 ), которые обычно обнаруживались среди более ранних палеолитических европейских останков, таких как Костенки-14 и Сунгирь . Отцовскую гаплогруппу C-V20 до сих пор можно обнаружить у мужчин, живущих в современной Испании , что свидетельствует о давнем присутствии этой линии в Западной Европе. Их митохондриальные хромосомы принадлежали преимущественно к гаплогруппе U5 . [12] [13]
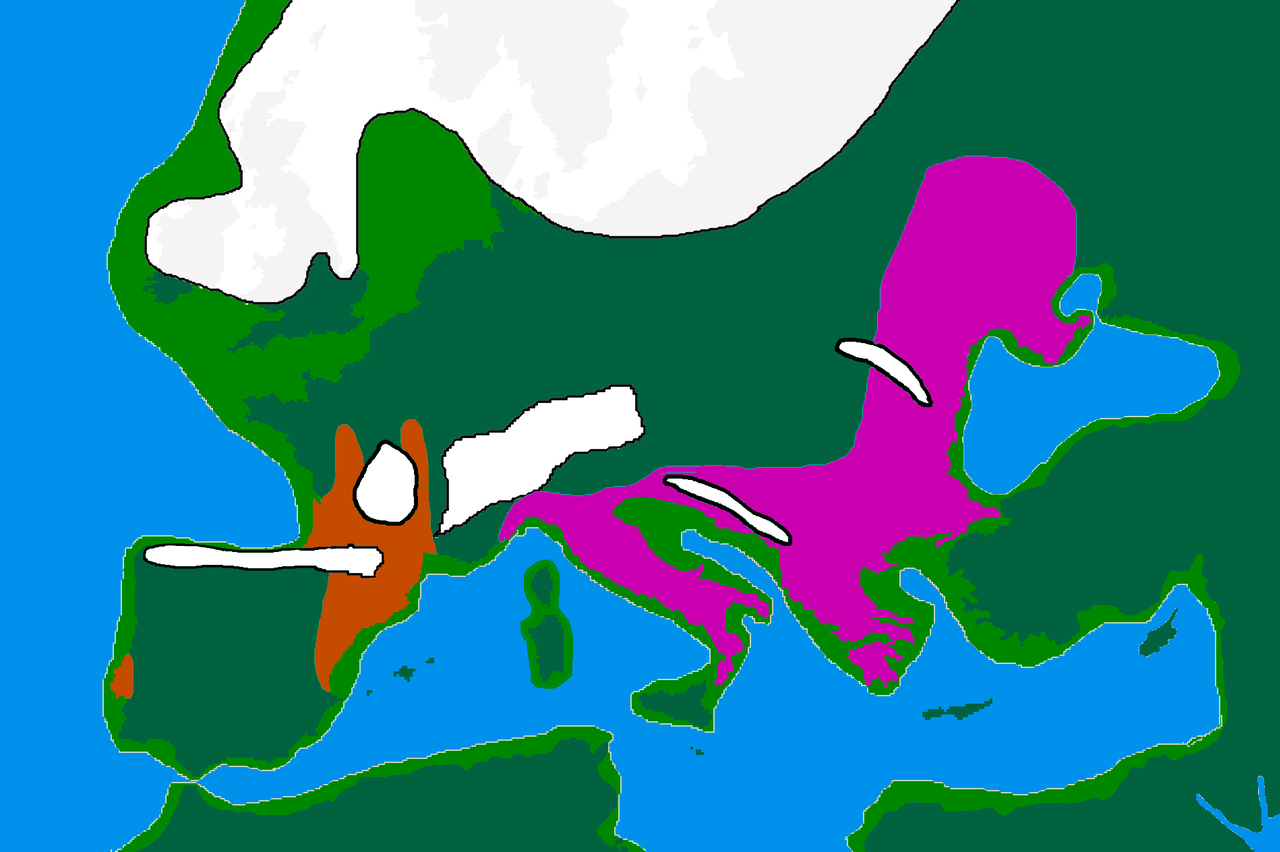
Самые ранние известные особи происхождения WHG/Виллабруна в Европе известны из Италии и датируются примерно 17 000 лет назад. [14] Ранние популяции WHG/Виллабруна связаны с эпиграветтской археологической культурой, которая в значительной степени заменила популяции, связанные с мадленской культурой около 14 000 лет назад (предками мадленских особей были популяции, связанные с западным граветтским , солютрейским и ориньякским периодами ). культуры). [12] [15] Родословная WHG впервые обнаружена к северу от Альп у двух особей возрастом 14 000 лет в Оберкасселе , Германия, которых можно смоделировать как примесь предков Виллабруны (которая сама моделируется как примесь линии, связанной с кластером Вестонице ). и предковая линия от особей Костенки-14 и Goyet Q116-1 ), а также родословная Goyet-Q2, связанная с особями, обнаруженными в Европе до последнего ледникового максимума. В исследовании говорится, что всех особей кластера Оберкассель можно смоделировать как c. 75% Виллабруна и 25% происхождение Гойе-Q2 или, альтернативно, как c. 90% предков Виллабруна и 10% предков Фурнола, недавно идентифицированного кластера, описанного как сестринская линия предков Гойета Q116-1, обнаруженного у людей, связанных с граветтской культурой юго-западной Европы. [12] Исследование предполагает, что родословная Оберкасселя в основном уже сформировалась до того, как распространилась, возможно, вокруг западной стороны Альп, в Западную и Центральную Европу и Великобританию, где отобранные особи WHG генетически однородны. Это контрастирует с прибытием предков Виллабруны и Оберкасселя в Иберию, которое, по-видимому, включало в себя повторяющиеся события примеси с местным населением, несущим высокий уровень предков Goyet-Q2. Это, а также выживание специфических клад гаплогруппы C1 Y-ДНК, ранее наблюдавшихся среди ранних европейских охотников-собирателей, предполагает относительно более высокую генетическую преемственность на юго-западе Европы в этот период. [12]
Есть признаки того, что WHG несет в себе «аллели риска диабета и болезни Альцгеймера ». [16]

Также было обнаружено, что WHG внесли свой вклад в происхождение популяций на границах Европы, таких как ранние анатолийские фермеры и древние африканцы Северо-Запада, [18] , а также другие европейские группы, такие как восточные охотники-собиратели . [19] Отношения WHGs с EHGs остаются неубедительными. [19] EHG моделируются так, чтобы получить различную степень происхождения от линии, связанной с WHG, в диапазоне от всего лишь 25% до 91%, а остальная часть связана с потоком генов от палеолитических сибиряков (ANE) и, возможно, кавказских охотников-собирателей . Другая линия, известная как скандинавские охотники-собиратели (SHG), представляет собой смесь EHG и WHG. [а] [21] [22]
Люди мезолитической кундинской культуры и нарвской культуры восточной Балтики представляли собой смесь WHG и EHG, [23] проявляя самое близкое родство с WHG. Было обнаружено , что образцы украинского мезолита и неолита плотно сгруппированы между WHG и EHG, что позволяет предположить генетическую преемственность в Днепровских порогах в течение 4000 лет. Украинские образцы принадлежали исключительно к материнской гаплогруппе U , которая встречается примерно в 80% всех европейских образцов охотников-собирателей. [24]
С ЭХГ были тесно связаны представители культуры ямочно-гребенчатой керамики (КСС) Восточной Балтики. [25] В отличие от большинства WHG, WHG восточной Балтики не получили примеси европейских фермеров во время неолита. Таким образом, современное население восточной Балтики имеет большее количество предков WHG, чем любое другое население Европы. [23]
Было обнаружено, что SHG содержат смесь компонентов WHG, которые, вероятно, мигрировали в Скандинавию с юга, и EHG, которые позже мигрировали в Скандинавию с северо-востока вдоль норвежского побережья. Эта гипотеза подтверждается данными о том, что у ГСП из западной и северной Скандинавии было меньше предков WHG (около 51%), чем у людей из восточной Скандинавии (около 62%). Считается, что WHG, проникшие в Скандинавию, принадлежали к аренсбургской культуре . EHG и WHG показали более низкие частоты аллелей SLC45A2 и SLC24A5 , которые вызывают депигментацию, и OCA/Herc2 , который вызывает светлый цвет глаз , чем SHG. [26]

ДНК одиннадцати WHG из верхнего палеолита и мезолита в Западной Европе , Центральной Европе и на Балканах была проанализирована на предмет их гаплогрупп Y-ДНК и гаплогрупп мтДНК . Анализ показал, что когда-то WHG были широко распространены от побережья Атлантического океана на западе до Сицилии на юге и до Балкан на юго-востоке на протяжении более шести тысяч лет. [27] Исследование также включало анализ большого количества людей из доисторической Восточной Европы. Тридцать семь образцов собрано с территории мезолитической и неолитической Украины (9500-6000 гг. до н.э.). Было установлено, что они являются промежуточными между EHG и SHG, хотя происхождение WHG в этой популяции увеличилось в эпоху неолита. Образцы Y-ДНК, выделенные от этих людей, принадлежали исключительно гаплотипам R (особенно субкладам R1b1 ) и гаплотипам I (особенно субкладам I2 ). мтДНК принадлежала почти исключительно U (особенно субкладам U5 и U4 ). [27] Было проанализировано большое количество лиц из могильника Звейниеки , которые в основном принадлежали кундинской культуре и нарвской культуре в восточной Балтике. На более ранних стадиях эти люди в основном были потомками WHG, но со временем происхождение EHG стало преобладающим. Y-ДНК этого сайта принадлежала почти исключительно гаплотипам гаплогруппы R1b1a1a и I2a1 . МтДНК принадлежала исключительно гаплогруппе U (особенно субкладам U2 , U4 и U5 ). [27] Также были проанализированы сорок человек из трех стоянок мезолита Железных ворот на Балканах . По оценкам, эти люди имели 85% WHG и 15% EHG происхождения. Самцы на этих участках несли исключительно гаплотипы гаплогруппы R1b1a и I (в основном субклады I2a ). мтДНК принадлежала в основном U (особенно субкладам U5 и U4 ). [27] Было обнаружено, что люди балканского неолита на 98% имеют анатолийское происхождение и на 2% - WHG-происхождение. К энеолиту люди Кукутенско -Трипольской культурыБыло обнаружено, что около 20% имеют предков охотников-собирателей, что занимает промежуточное положение между EHG и WHG. Было обнаружено , что люди культуры шаровидных амфор укрывают ок. 25% предков WHG, что значительно выше, чем у групп среднего неолита Центральной Европы. [27]


В плодотворном исследовании 2014 года впервые был выявлен вклад трех основных компонентов в современные европейские линии: западных охотников-собирателей (WHG, в пропорциях до 50% у северных европейцев), древних северных евразийцев (ANE, сибиряки верхнего палеолита, позже связанные с более поздняя индоевропейская экспансия , присутствующая в пропорциях до 20%), и, наконец, ранние европейские земледельцы (EEF, земледельцы в основном ближневосточного происхождения, которые мигрировали в Европу примерно с 8000 лет назад, сейчас присутствуют в пропорциях примерно 30% в Прибалтике). регионе до примерно 90% в Средиземноморье). Компонент «Ранние европейские фермеры» (EEF) был идентифицирован на основе генома женщины, похороненной ок. 7000 лет назад в могиле культуры линейной керамики в Штутгарте , Германия. [29]
Это исследование 2014 года обнаружило доказательства генетического смешения между WHG и EEF по всей Европе, с наибольшим вкладом EEF в Средиземноморской Европе (особенно на Сардинии, Сицилии, Мальте и среди евреев-ашкенази), а также наибольшим вкладом WHG в Северной Европе и среди басков. люди. [30]
С 2014 года дальнейшие исследования уточнили картину скрещивания EEF и WHG. В ходе анализа 180 наборов древних данных ДНК периодов энеолита и неолита из Венгрии, Германии и Испании в 2017 году были обнаружены доказательства длительного периода скрещивания. Смешение происходило на региональном уровне, из местных популяций охотников-собирателей, так что популяции из трех регионов (Германия, Иберия и Венгрия) были генетически различимы на всех этапах периода неолита, с постепенно увеличивающимся соотношением предков WHG среди земледельческих популяций с течением времени. . Это говорит о том, что после первоначальной экспансии первых земледельцев не было дальнейших дальних миграций, достаточно существенных, чтобы гомогенизировать земледельческое население, и что земледельческое население и население охотников-собирателей существовали бок о бок в течение многих столетий, с продолжающимся постепенным смешением на протяжении 5-го века. до 4-го тысячелетия до нашей эры (а не единичное событие примеси при первом контакте). [31] Уровень примеси варьировался в зависимости от географического положения; в эпоху позднего неолита доля WHG у фермеров в Венгрии составляла около 10%, в Германии - около 25%, а в Иберии - до 50%. [32]
Анализ останков из Грота Континенца в Италии показал, что из шести останков три захоронены между ок. 10 000 г. до н. э. и 7 000 г. до н. э. принадлежали I2a-P214 ; и вдвое больше материнских гаплогрупп U5b1 и одна U5b3 . [33] [34] Около 6000 г. до н.э. WHG Италии были почти полностью генетически заменены EEF (два G2a2 ) и одной гаплогруппой R1b , хотя в последующие тысячелетия родословная WHG немного увеличилась. [35]
Люди неолита на Британских островах были близки к иберийским и центральноевропейским популяциям раннего и среднего неолита, которые, согласно модели, имели около 75% предков от EEF, а остальная часть - от WHG в континентальной Европе. Впоследствии они заменили большую часть населения WHG на Британских островах, не смешиваясь с ними. [36]
По оценкам, WHG внесли 20-30% предков в неолитические группы EEF по всей Европе. Специфические приспособления против местных патогенов могли быть привнесены посредством мезолитической примеси WHG в неолитические популяции EEF. [37]
Исследование мезолитических охотников-собирателей из Дании показало, что они были связаны с современными западными охотниками-собирателями и связаны с культурами Маглемосе , Конгемосе и Эртебёлле . Они демонстрировали «генетическую однородность примерно от 10 500 до 5 900 лет назад», пока «не прибыли неолитические фермеры анатолийского происхождения». Переход к периоду неолита был «очень резким и привел к смене населения с ограниченным генетическим вкладом местных охотников-собирателей. Последующая неолитическая популяция была связана с культурой воронкообразных клювов» . [38]

По словам Дэвида Райха , анализ ДНК показал, что западные охотники-собиратели обычно были темнокожими, темноволосыми и голубоглазыми. [41] Темная кожа была следствием их внеафриканского происхождения (у всех популяций Homo sapiens изначально была темная кожа), а голубые глаза были результатом вариации гена OCA2, что вызывало депигментацию радужной оболочки . [42]
Археолог Грэм Уоррен сказал, что цвет их кожи варьировался от оливкового до черного, и предположил, что у них могли быть некоторые региональные различия цвета глаз и волос. [43] Это разительно отличается от дальних родственников восточных охотников-собирателей (EHG), которые, как предполагалось, были светлокожими, кареглазыми или голубоглазыми, темноволосыми или светловолосыми. [44]
По прогнозам , два скелета WHG с неполными SNP , Ла Бранья и Человек из Чеддера , имели темную или от темного до черного цвета кожу, тогда как два других скелета WHG с полными SNP, «Свен» и человек из Лошбора , по прогнозам, имели темную или средне-черную кожу. к темной и промежуточной коже соответственно. [45] [26] [b] Испанский биолог Карлес Лалуэса-Фокс сказал, что у особи Ла Бранья-1 была темная кожа, «хотя мы не можем знать точный оттенок». [47]
Согласно исследованию 2020 года, прибытие ранних европейских фермеров (EEF) из Западной Анатолии от 8500 до 5000 лет назад вместе с пастухами западных степей в бронзовом веке вызвало быструю эволюцию европейского населения в сторону более светлой кожи и волос. [42] Смешение охотников-собирателей и земледельцев, по-видимому, было случайным, но не обширным. [48]
.png/1280px-Archaeogenetic_analysis_of_human_skin_pigmentation_in_Europe_(with_Asia_geographic_extension).png)
Некоторые авторы выражают осторожность в отношении реконструкций пигментации кожи : Quillen et al. (2019) признают исследования, которые в целом показывают, что «более светлый цвет кожи был редкостью на большей части Европы во время мезолита», включая исследования, касающиеся предсказаний «темного или темного или темно-черного» человека Чеддера, но предупреждают, что «реконструкции мезолита и неолита Фенотип пигментации с использованием локусов, распространенных в современных популяциях, следует интерпретировать с некоторой осторожностью, поскольку вполне возможно, что другие, еще не изученные локусы, также могли влиять на фенотип». [49]
Генетик Сьюзан Уолш из Университета Индианы – Университета Пердью в Индианаполисе , которая работала над проектом «Человек Чеддер», сказала, что «мы просто не знаем цвет его кожи». [50] Немецкий биохимик Йоханнес Краузе заявил, что мы не знаем, был ли цвет кожи западноевропейских охотников-собирателей больше похож на цвет кожи людей из современной Центральной Африки или людей из арабского региона . Достоверно лишь то, что они не несли какой-либо известной мутации, ответственной за светлую кожу у последующих популяций европейцев. [51]
Исследование геномного происхождения и социальной динамики последних охотников-собирателей Атлантической Франции, проведенное в 2024 году, показало, что «фенотипически мы обнаруживаем некоторое разнообразие во время позднего мезолита во Франции», при котором два из секвенированных в исследовании WHG «вероятно имели пигментация кожи от бледной до средней», но «большинство людей имеют темную кожу и голубые глаза, характерные для WHG» изученных образцов. [52]
Начиная с скопления Виллабруна, по крайней мере, ~ 14 000 лет назад, все проанализированные европейские особи демонстрируют близость к Ближнему Востоку. По времени это коррелирует с интерстадиалом Бёллинг-Аллерёд, первым значительным периодом потепления после ледникового периода. Археологически это коррелирует с культурными переходами внутри эпиграветта в Южной Европе и переходом от Магдалины к Азилию в Западной Европе. Таким образом, появление скопления Виллабруна может отражать миграции или популяционные сдвиги внутри Европы в конце ледникового периода, и это наблюдение также согласуется с данными об обновлении последовательностей митохондриальной ДНК в это время. Одним из сценариев, который мог бы объяснить эти закономерности, является расширение населения из рефугиумов Юго-Восточной Европы или Западной Азии после ледникового периода, объединяющее генетическое происхождение Европы и Ближнего Востока. В-шестых, в кластере Виллабруна некоторые, но не все, люди имеют близость к выходцам из Восточной Азии. Важным направлением будущей работы является получение аналогичных данных древней ДНК из Юго-Восточной Европы и Ближнего Востока, чтобы получить более полную картину истории верхнего палеолита населения Западной Евразии.
{{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка ){{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )Благодаря этому анализу мы идентифицировали хромосому из южной Европы как новую глубокую ветвь гаплогруппы C (C-V20 или C7, рисунок S1). Ранее в Южной Европе было обнаружено лишь несколько примеров С-хромосом (определяемых только маркером RPS4Y711) [32], [33]. Чтобы улучшить наши знания о распространении гаплогруппы C в Европе, мы опросили 1965 европейцев на наличие мутации RPS4Y711 и идентифицировали одну дополнительную хромосому гаплогруппы C из южной Европы, которая также была классифицирована как C7 (данные не показаны).
... тогда как аллели риска диабета и болезни Альцгеймера обогащены западными предками охотников-собирателей.
Большинство современных европейцев происходят как минимум из трех высокодифференцированных популяций: западноевропейских охотников-собирателей, которые внесли свой вклад в происхождение всех европейцев, но не жителей Ближнего Востока; древние северные евразийцы, родственные сибирякам верхнего палеолита, которые внесли свой вклад как в европейцев, так и в жителей Ближнего Востока; и ранние европейские фермеры, которые были в основном выходцами с Ближнего Востока, но также имели предков, связанных с западноевропейскими охотниками-собирателями.
Человек разумный прибыл в Европу с Ближнего Востока около 42 000 лет назад. Как и в случае с африканским происхождением, эти люди имели темную кожу, но из-за вариаций гена OCA2 (вызывающего депигментацию радужной оболочки) у многих из них были голубые глаза» (...) «в южной и центральной Европе, где они [аллели светлой кожи] были завезен фермерами из Западной Анатолии, расширявшимися от 8500 до 5000 лет назад. Это было началом неолитической революции в этих регионах, характеризующейся более оседлым образом жизни и одомашниванием некоторых видов животных и растений. (...) «Быстрый рост численности населения в результате неолитической революции, такой как использование молочных продуктов в качестве источника пищи для взрослых и развитие сельского хозяйства, а также массовое распространение ямных скотоводов, вероятно, вызвали быструю селекцию европейское население стремится к светлой коже и волосам
Как обнаружила команда Экскофье, после того, как они обосновались в Анатолии, ранние земледельческие популяции начали постепенно перемещаться на запад, в Европу, начиная примерно с 8000 лет назад. Время от времени они смешивались, но не слишком часто, с местными охотниками-собирателями.
Их анализ показывает, что цвет кожи обоих людей, вероятно, был темным, а цвет кожи мезолитического человека из Чеддера, по прогнозам, был «темным или от темного до черного». Эти данные позволяют предположить, что более светлый цвет кожи был редкостью на большей части территории Европы в эпоху мезолита. Однако это не противоречит оценкам дат <20 тыс. лет назад, которые касаются начала отбора, а не времени фиксации предпочтительных аллелей (Beleza et al., 2013; Beleza, Johnson и др., 2013). . Хотя исследования древнего генома предсказывают, что среди мезолитических европейцев в целом более темный цвет кожи, производные аллели rs1426654 и rs16891982 разделялись в европейских популяциях во время мезолита (González-Fortes et al., 2017; Günther et al., 2018; Mittnik et al., 2018). ), что позволяет предположить, что к этому времени, вероятно, уже присутствовали фенотипические вариации, обусловленные этими локусами. Однако реконструкции фенотипа пигментации мезолита и неолита с использованием локусов, распространенных в современных популяциях, следует интерпретировать с некоторой осторожностью, поскольку возможно, что другие, еще не изученные локусы также могли повлиять на фенотип.
_to_Villabruna_(orange)_genetic_types_in_European_sites,_according_to_timeline_and_climate_evolution.jpg/1280px-Transition_from_Goyet_(green)_to_Villabruna_(orange)_genetic_types_in_European_sites,_according_to_timeline_and_climate_evolution.jpg){kind=link}