
Абиссальная равнина — это подводная равнина на глубоком дне океана , обычно встречающаяся на глубинах от 3000 до 6000 метров (от 9800 до 19700 футов). Расположенные обычно между подножьем континентального поднятия и срединно-океаническим хребтом , абиссальные равнины покрывают более 50% поверхности Земли . [1] [2] Это одни из самых плоских, гладких и наименее изученных регионов на Земле. [3] Абиссальные равнины являются ключевыми геологическими элементами океанических бассейнов (другими элементами являются приподнятый срединно-океанический хребет и примыкающие к нему абиссальные холмы ).
Создание абиссальной равнины — результат расширения морского дна (тектоники плит) и плавления нижней океанической коры . Магма поднимается над астеносферой (слоем верхней мантии ), и по мере того, как этот базальтовый материал достигает поверхности срединно-океанических хребтов, он образует новую океаническую кору, которая постоянно оттягивается в сторону в результате расширения морского дна. Абиссальные равнины возникают в результате покрытия первоначально неровной поверхности океанической коры мелкозернистыми отложениями , главным образом глиной и илом . Большая часть этих отложений откладывается мутными потоками , которые переносятся с окраин континента вдоль подводных каньонов в более глубокие воды. Остальное состоит в основном из пелагических отложений . В некоторых районах равнин распространены металлические конкреции с различной концентрацией металлов, включая марганец , железо , никель , кобальт и медь . Также имеется некоторое количество углерода, азота, фосфора и кремния из-за материалов, которые оседают и разлагаются.
Отчасти из-за своих огромных размеров абиссальные равнины считаются основными резервуарами биоразнообразия . Они также оказывают значительное влияние на круговорот углерода в океане , растворение карбоната кальция и концентрацию CO 2 в атмосфере в масштабах времени от ста до тысячи лет. На структуру абиссальных экосистем сильное влияние оказывают скорость поступления пищи на морское дно и состав оседающего материала. Такие факторы, как изменение климата , методы рыболовства и удобрение океана, оказывают существенное влияние на структуру первичной продукции в эвфотической зоне . [1] [4] Животные поглощают растворенный кислород из бедных кислородом вод. Большая часть растворенного кислорода на абиссальных равнинах поступала из полярных регионов, которые давно растаяли. Из-за нехватки кислорода абиссальные равнины непригодны для организмов, которые могли бы процветать в обогащенных кислородом водах наверху. Глубоководные коралловые рифы встречаются в основном на глубинах 3000 метров и глубже в абиссальной и хадальной зонах .
Абиссальные равнины не считались отдельными физико-географическими особенностями морского дна до конца 1940-х годов, и до недавнего времени ни одна из них не изучалась на систематической основе. Они плохо сохранились в осадочных отложениях , поскольку имеют тенденцию поглощаться процессом субдукции. Из-за темноты и давления воды, которое может достигать примерно в 750 раз атмосферного давления (76 мегапаскалей), абиссальные равнины изучены недостаточно.

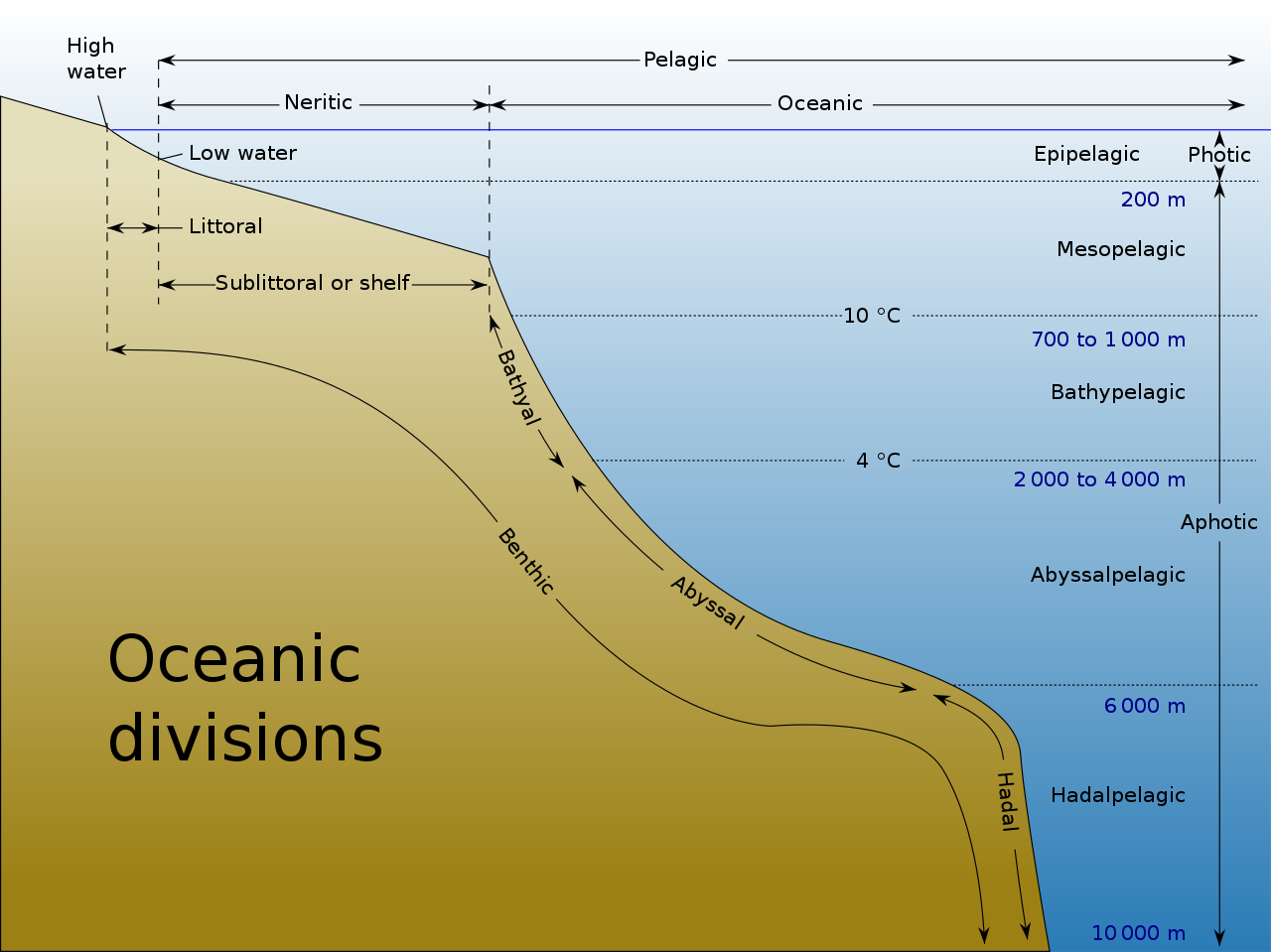
Океан можно представить как зоны в зависимости от глубины и наличия или отсутствия солнечного света . Почти все формы жизни в океане зависят от фотосинтетической деятельности фитопланктона и других морских растений по преобразованию углекислого газа в органический углерод , который является основным строительным блоком органического вещества . Фотосинтез, в свою очередь, требует энергии солнечного света для запуска химических реакций, в результате которых образуется органический углерод. [5]
Слой толщи воды , ближайший к поверхности океана ( уровню моря ), называется фотической зоной . Фотическую зону можно разделить на две разные вертикальные области. Самая верхняя часть фотической зоны, где достаточно света для поддержки фотосинтеза фитопланктона и растений, называется эвфотической зоной (также называемой эпипелагической зоной или поверхностной зоной ). [6] Нижняя часть фотической зоны, где интенсивность света недостаточна для фотосинтеза, называется дисфотической зоной (по-гречески «дисфотический» означает «плохо освещенный»). [7] Дисфотическую зону также называют мезопелагической зоной или сумеречной зоной . [8] Его самая нижняя граница находится на термоклине 12 ° C (54 ° F), который в тропиках обычно находится на высоте от 200 до 1000 метров. [9]
Эфотическую зону несколько условно определяют как простирающуюся от поверхности до глубины, где интенсивность света составляет примерно 0,1–1% от поверхностного солнечного излучения , в зависимости от времени года , широты и степени мутности воды . [6] [7] В самой чистой океанской воде эвфотическая зона может простираться на глубину около 150 метров, [6] или, реже, до 200 метров. [8] Растворенные вещества и твердые частицы поглощают и рассеивают свет, а в прибрежных районах высокая концентрация этих веществ приводит к быстрому ослаблению света с глубиной. В таких районах глубина эвфотической зоны может составлять всего несколько десятков метров или меньше. [6] [8] Дисфотическая зона, где интенсивность света составляет значительно менее 1% от поверхностной освещенности, простирается от основания эвфотической зоны примерно до 1000 метров. [9] От дна фотической зоны до морского дна простирается афотическая зона , область вечной тьмы. [8] [9]
Поскольку средняя глубина океана составляет около 4300 метров, [10] фотическая зона представляет собой лишь небольшую часть общего объема океана. Однако благодаря своей способности к фотосинтезу фотическая зона обладает наибольшим биоразнообразием и биомассой среди всех океанических зон. Здесь происходит почти вся первичная продукция океана. Формы жизни, населяющие афотическую зону, часто способны перемещаться вверх по толще воды в фотическую зону для питания. В противном случае им придется полагаться на опускание материала сверху [ 1] или найти другой источник энергии и питания, как это происходит у хемосинтезирующих архей , обитающих вблизи гидротермальных источников и холодных выходов .
Афотическую зону можно разделить на три различные вертикальные области в зависимости от глубины и температуры. Первая — это батиальная зона , простирающаяся от глубины 1000 метров до 3000 метров, при этом температура воды снижается с 12 °C (54 °F) до 4 °C (39 °F) по мере увеличения глубины. [11] Далее идет абиссальная зона , простирающаяся от глубины 3000 метров до 6000 метров. [11] Последняя зона включает в себя глубокие океанические желоба и известна как зона хадала . Эта самая глубокая океаническая зона простирается от глубины 6000 метров до примерно 11 034 метров, на самом дне Марианской впадины, самой глубокой точки на планете Земля. [2] [11] Абиссальные равнины обычно находятся в абиссальной зоне на глубине от 3000 до 6000 метров. [1]
В таблице ниже представлена классификация океанических зон:


Океаническая кора, образующая основу абиссальных равнин, постоянно создается на срединно-океанических хребтах (тип расходящейся границы ) в результате процесса, известного как декомпрессионное плавление . [20] Декомпрессионное плавление твердой мантии, связанное с плюмами , ответственно за создание океанских островов, таких как Гавайские острова , а также океанской коры на срединно-океанических хребтах. Это явление также является наиболее распространенным объяснением пойменных базальтов и океанических плато (два типа крупных магматических провинций ). Декомпрессионное плавление происходит, когда верхняя мантия частично расплавляется в магму по мере ее продвижения вверх под срединно-океанические хребты. [21] [22] Эта поднимающаяся вверх магма затем охлаждается и затвердевает за счет проводимости и конвекции тепла, образуя новую океаническую кору . Аккреция происходит по мере присоединения мантии к растущим краям тектонической плиты , что обычно связано с расширением морского дна . Таким образом, возраст океанической коры зависит от расстояния от срединно-океанического хребта. [23] Самая молодая океаническая кора находится на срединно-океанических хребтах, и она становится все старше, холоднее и плотнее по мере миграции от срединно-океанических хребтов в рамках процесса, называемого мантийной конвекцией . [24]
Литосфера , лежащая поверх астеносферы , разделена на ряд тектонических плит, которые постоянно создаются и поглощаются на противоположных границах плит . Океаническая кора и тектонические плиты формируются и раздвигаются на срединно-океанических хребтах. Абиссальные холмы образуются в результате растяжения океанической литосферы. [25] Потребление или разрушение океанической литосферы происходит в океанических впадинах (тип сходящейся границы , также известный как деструктивная граница плиты) в результате процесса, известного как субдукция . Океанические желоба встречаются в местах, где встречаются океанические литосферные плиты двух разных плит, и более плотная (более древняя) плита начинает опускаться обратно в мантию. [26] На краю плиты (океанический желоб) океаническая литосфера термически сжалась, стала достаточно плотной и опустилась под собственным весом в процессе субдукции. [27] Процесс субдукции поглощает более старую океаническую литосферу, поэтому возраст океанической коры редко превышает 200 миллионов лет. [28] Общий процесс повторяющихся циклов создания и разрушения океанической коры известен как цикл Суперконтинента , впервые предложенный канадским геофизиком и геологом Джоном Тузо Уилсоном .
Новая океаническая кора, ближайшая к срединно-океаническим хребтам, на мелких уровнях в основном состоит из базальта и имеет пересеченный рельеф . Неровность этой топографии является функцией скорости расширения срединно-океанического хребта (скорости расширения). [29] Величины скорости распространения варьируются весьма существенно. Типичные значения для быстро спрединговых хребтов превышают 100 мм/год, тогда как для медленно спрединговых хребтов обычно менее 20 мм/год. [21] Исследования показали, что чем медленнее скорость распространения, тем шероховатой будет новая океаническая кора, и наоборот. [29] Считается, что это явление связано с разломами на срединно-океаническом хребте, когда формировалась новая океаническая кора. [30] Эти разломы, пронизывающие океаническую кору, вместе с ограничивающими их абиссальными холмами, являются наиболее распространенными тектоническими и топографическими особенностями на поверхности Земли. [25] [30] Процесс расширения морского дна помогает объяснить концепцию дрейфа континентов в теории тектоники плит.
Плоский вид зрелых абиссальных равнин является результатом покрытия этой первоначально неровной поверхности океанической коры мелкозернистыми отложениями, главным образом глиной и илом. Большая часть этих отложений откладывается в результате мутных потоков, которые были перенесены с окраин континента вдоль подводных каньонов вниз, в более глубокие воды. Остальная часть осадков состоит в основном из пыли (частиц глины), вынесенной в море с суши, а также остатков мелких морских растений и животных , опускающихся из верхнего слоя океана, известных как пелагические отложения . Общая скорость отложения осадков в отдаленных районах оценивается в два-три сантиметра за тысячу лет. [31] [32] Покрытые отложениями абиссальные равнины менее распространены в Тихом океане, чем в других крупных океанских бассейнах, поскольку отложения мутных течений задерживаются в океанических желобах, граничащих с Тихим океаном. [33]
Абиссальные равнины обычно покрыты глубоким морем, но во время Мессинского кризиса солености большая часть абиссальной равнины Средиземного моря подвергалась воздействию воздуха как пустая глубокая, горячая и сухая раковина с соляным дном. [34] [35] [36] [37]

Знаменательная научная экспедиция (декабрь 1872 г. – май 1876 г.) исследовательского корабля Британского Королевского флота HMS Challenger дала огромное количество батиметрических данных, большая часть которых была подтверждена последующими исследователями. Батиметрические данные, полученные в ходе экспедиции «Челленджер», позволили ученым составить карты, [38] на которых были представлены приблизительные очертания некоторых основных особенностей подводной местности, таких как края континентальных шельфов и Срединно-Атлантический хребет . Этот прерывистый набор точек данных был получен с помощью простой техники измерения путем опускания длинных тросов с корабля на морское дно. [39]
За экспедицией «Челленджер» последовала экспедиция «Жаннет » 1879–1881 годов под руководством лейтенанта ВМС США Джорджа Вашингтона Делонга . Команда пересекла Чукотское море и записала метеорологические и астрономические данные, а также провела зондирование морского дна. Корабль застрял в паке льда возле острова Врангеля в сентябре 1879 года и в конечном итоге был раздавлен и затоплен в июне 1881 года. [40]
За экспедицией «Жаннет» последовала арктическая экспедиция 1893–1896 годов норвежского исследователя Фритьофа Нансена на борту « Фрама » , которая доказала, что Северный Ледовитый океан представляет собой глубокий океанический бассейн, не прерываемый какими-либо значительными массивами суши к северу от Евразийского континента. [41] [42]
Начиная с 1916 года канадский физик Роберт Уильям Бойль и другие ученые из Комитета по расследованию противолодочного обнаружения ( ASDIC ) предприняли исследования, которые в конечном итоге привели к разработке гидроакустической технологии. Было разработано оборудование для акустического зондирования , которое могло работать гораздо быстрее, чем линии зондирования, что позволило немецкой экспедиции «Метеор» на борту немецкого исследовательского судна « Метеор» (1925–27) проводить частые зондирования на трансектах восток-запад Атлантического океана. Карты, созданные с помощью этих методов, показывают основные атлантические бассейны, но точность глубины этих ранних инструментов была недостаточной, чтобы выявить плоские безликие абиссальные равнины. [43] [44]
По мере совершенствования технологий измерения глубины, широты и долготы становились более точными, и стало возможным собирать более или менее непрерывные наборы точек данных. Это позволило исследователям составить точные и подробные карты больших участков дна океана. Использование эхолота с непрерывной записью позволило Толстому и Юингу летом 1947 года идентифицировать и описать первую абиссальную равнину. Эта равнина, расположенная к югу от Ньюфаундленда , теперь известна как Абиссальная равнина Сом . [45] После этого открытия во всех океанах было обнаружено множество других примеров. [46] [47] [48] [49] [50]
Бездна Челленджера — самая глубокая исследуемая точка из всех океанов Земли; он находится на южном конце Марианской впадины , недалеко от группы Марианских островов . Впадина названа в честь HMS Challenger , исследователи которого сделали первые записи ее глубины 23 марта 1875 года на станции 225. Сообщенная глубина составила 4475 саженей (8184 метра) на основе двух отдельных измерений. 1 июня 2009 года гидролокационное картирование Бездны Челленджера с помощью многолучевой гидролокационной батиметрической системы Simrad EM120 на борту научно-исследовательского судна Kilo Moana показало, что максимальная глубина составляет 10971 метр (6,82 мили). Гидроакустическая система использует фазовое и амплитудное определение дна с точностью не хуже 0,2% от глубины воды (на этой глубине это погрешность около 22 метров). [51] [52]

Редкой, но важной особенностью местности, обнаруженной в батиальной, абиссальной и хадальной зонах, является гидротермальный источник. В отличие от температуры окружающей среды на этих глубинах около 2 °C, вода выходит из этих отверстий при температуре от 60 °C до 464 °C. [13] [14] [15] [16] [17] Из-за высокого барометрического давления на этих глубинах вода может существовать либо в жидкой форме, либо в виде сверхкритической жидкости при таких температурах.
При барометрическом давлении 218 атмосфер критическая точка воды составляет 375°С. На глубине 3000 метров барометрическое давление морской воды составляет более 300 атмосфер (поскольку соленая вода плотнее пресной ). На такой глубине и давлении морская вода становится сверхкритической при температуре 407 °C ( см. изображение ). Однако увеличение солености на этой глубине приближает воду к критической точке. Таким образом, вода, выходящая из самых горячих частей некоторых гидротермальных источников, черных курильщиков и подводных вулканов , может быть сверхкритической жидкостью , обладающей физическими свойствами между свойствами газа и жидкости . [13] [14] [15] [16] [17]
Пик Сестер ( гидротермальное поле Комфортесс-Коув, 4 ° 48 ' ю.ш., 12 ° 22' з.д. / 4,800 ° ю . , 4 ° 48'S 12 ° 23'W / 4,800 ° S 12,383 ° W / -4,800; -12,383 , высота -3047 м), три гидротермальных источника категории черного курильщика на Срединно-Атлантическом хребте. недалеко от острова Вознесения . Предполагается, что они были активны после землетрясения, потрясшего этот регион в 2002 году . В 2008 году в одном из этих источников была зафиксирована устойчивая температура на выходе до 407 °C, а максимальная зарегистрированная температура достигала 464 °C. Эти термодинамические условия превышают критическую точку морской воды и представляют собой самые высокие температуры, зарегистрированные на сегодняшний день на морском дне. Это первое зарегистрированное свидетельство прямого магматико - гидротермального взаимодействия на медленно спрединговом срединно-океаническом хребте. [13] [14] [15] [16] [17] Начальные этапы работы вентиляционного дымохода начинаются с отложения минерального ангидрита. Сульфиды меди, железа и цинка затем оседают в зазорах дымохода, что с течением времени делает его менее пористым. Был зарегистрирован рост вентиляционных отверстий порядка 30 см (1 фут) в день. Исследование глубоководных жерл у побережья Фиджи, проведенное в апреле 2007 года, показало, что эти жерла являются важным источником растворенного железа (см. Цикл железа).
Гидротермальные жерла в глубоком океане обычно образуются вдоль срединно-океанических хребтов, таких как Восточно-Тихоокеанское поднятие и Срединно-Атлантический хребет. Это места, где две тектонические плиты расходятся и формируется новая кора.

Еще одной необычной особенностью, обнаруженной в абиссальной и хадальной зонах, является холодное просачивание , иногда называемое холодным выходом . Это область морского дна, где происходит просачивание сероводорода , метана и других богатых углеводородами жидкостей, часто в виде глубоководных рассолов . Первые холодные просачивания были обнаружены в 1983 году на глубине 3200 метров в Мексиканском заливе . [53] С тех пор холодные просачивания были обнаружены во многих других районах Мирового океана , включая подводный каньон Монтерей недалеко от залива Монтерей , Калифорния, Японского моря , у тихоокеанского побережья Коста-Рики , у атлантического побережья Африка, у берегов Аляски и под шельфовым ледником в Антарктиде . [54]
Хотя когда-то считалось, что равнины представляют собой обширную пустынную среду обитания, исследования, проведенные за последнее десятилетие или около того, показывают, что они изобилуют разнообразной микробной жизнью. [55] [56] Однако структура и функции экосистемы на глубоком морском дне исторически были плохо изучены из-за размера и удаленности пропасти. Недавние океанографические экспедиции, проведенные международной группой ученых из организации « Перепись разнообразия глубоководной морской жизни» (CeDAMar), обнаружили чрезвычайно высокий уровень биоразнообразия на абиссальных равнинах: до 2000 видов бактерий, 250 видов простейших и 500 видов. беспозвоночных ( червей , ракообразных и моллюсков ), обычно встречающихся в отдельных абиссальных участках. [57] Новые виды составляют более 80% тысяч видов морских беспозвоночных, собранных на любой глубинной станции, что подчеркивает наше до сих пор плохое понимание глубинного разнообразия и эволюции. [57] [58] [59] [60] Более богатое биоразнообразие связано с областями известного поступления фитодетрита и более высоким потоком органического углерода. [61]
Abyssobrotula galatheae , вид обыкновенного угря семейства Ophidiidae , является одним из самых глубокоживущих видов рыб. В 1970 году один экземпляр был выловлен с глубины 8370 метров во желобе Пуэрто-Рико . [62] [63] [64] Однако по прибытии на поверхность животное было мертво. В 2008 году улитка-хадал ( Pseudoliparis amblystomopsis ) [65] была замечена и зарегистрирована на глубине 7700 метров в Японском желобе . На сегодняшний день это самая глубокая живая рыба, когда-либо зарегистрированная. [11] [66] Другие рыбы абиссальной зоны включают рыб семейства Ipnopidae , в которое входят абиссальная рыба-паук ( Bathypterois longipes ), тренога ( Bathypterois grallator ), рыба-щуп ( Bathypterois longifilis ) и черная ящерица ( Bathysauropsis gracilis ) . ). Некоторые представители этого семейства были зафиксированы с глубины более 6000 метров. [67]
Ученые CeDAMar продемонстрировали, что некоторые абиссальные и хадальные виды имеют космополитическое распространение. Одним из примеров этого могут быть простейшие фораминиферы , [68] некоторые виды которых распространены от Арктики до Антарктики. Другие группы фауны, такие как многощетинковые черви и равноногие ракообразные, по-видимому, являются эндемиками определенных равнин и бассейнов. [57] Многие, по-видимому, уникальные таксоны нематодных червей также были недавно обнаружены на абиссальных равнинах. Это говорит о том, что глубокие глубины океана способствовали адаптивному излучению . [57] Таксономический состав фауны нематод абиссальной части Тихого океана аналогичен, но не идентичен, таковому в Северной Атлантике. [61] Список некоторых видов, открытых или переописанных CeDAMar, можно найти здесь.
Одиннадцать из 31 описанного вида моноплакофор ( класс моллюсков ) обитают ниже 2000 метров . Из этих 11 видов два обитают исключительно в зоне хадал. [69] Наибольшее количество моноплакофор обитает в восточной части Тихого океана вдоль океанических желобов. Однако в западной части Тихого океана абиссальные моноплакофоры пока не обнаружены, а в Индийском океане идентифицирован только один абиссальный вид. [69] Из 922 известных видов хитонов (из класса моллюсков Polyplacophora ) 22 вида (2,4%), как сообщается, обитают на глубине ниже 2000 метров, а два из них обитают на абиссальной равнине. [69] Хотя генетические исследования отсутствуют, по крайней мере шесть из этих видов считаются эврибатными (способными жить в широком диапазоне глубин), поскольку, как сообщается, они встречаются от сублиторали до абиссальных глубин. Большое количество полиплакофор с больших глубин являются травоядными или ксилофагами , что может объяснить разницу между распространением моноплакофор и полиплакофор в Мировом океане. [69]
Известно, что перакаридные ракообразные, в том числе изоподы, составляют значительную часть сообщества макробентоса, ответственного за поедание крупных остатков пищи, падающей на морское дно. [1] [70] В 2000 году учёные экспедиции «Разнообразие глубоководного атлантического бентоса» (DIVA 1) (рейс М48/1 немецкого научно-исследовательского судна « Метеор III ») обнаружили и собрали три новых вида подотряда донных изопод Asellota . с абиссальных равнин Ангольского бассейна в южной части Атлантического океана . [71] [72] [73] В 2003 году Де Бройер и др. собрал около 68 000 ракообразных-перакарид 62 видов из ловушек с наживкой, расставленных в море Уэдделла , море Скоша и у Южных Шетландских островов . Они обнаружили, что около 98% экземпляров принадлежали к надсемейству амфипод Lysianassoidea , а 2% — к семейству изопод Cirolanidae . Половина этих видов была собрана с глубины более 1000 метров. [70]
В 2005 году дистанционно управляемый аппарат KAIKO Японского агентства по морским наукам и технологиям о Земле (JAMSTEC) собрал керн отложений из бездны Челленджера. В пробах осадков выявлено 432 живых экземпляра мягкостенных фораминифер. [74] [75] Фораминиферы — это одноклеточные протисты , которые строят раковины. По оценкам, существует около 4000 видов живых фораминифер. Из 432 собранных организмов подавляющее большинство выборки состояло из простых фораминифер с мягким панцирем, а остальные представляли виды сложных, многокамерных родов Leptohalysis и Reophax . В целом 85% экземпляров составили аллогромиииды с мягким панцирем . Это необычно по сравнению с образцами организмов, обитающих в отложениях, из других глубоководных сред, где процент фораминифер с органическими стенками колеблется от 5% до 20% от общего числа. Небольшим организмам с твердым известковым панцирем трудно расти на большой глубине, поскольку в воде на этой глубине катастрофически не хватает карбоната кальция. [76] Гигантские (5–20 см) фораминиферы, известные как ксенофиофоры , встречаются только на глубинах 500–10 000 метров, где они могут встречаться в больших количествах и значительно увеличивать разнообразие животных за счет их биотурбации и обеспечения среды обитания для мелких животных. . [77]
Хотя известно, что подобные формы жизни существуют в более мелких океанических впадинах (>7000 м) и на абиссальной равнине, формы жизни, обнаруженные в Бездне Челленджера, могут представлять собой независимые таксоны из этих более мелководных экосистем. Такое преобладание организмов с мягким панцирем в Бездне Челленджера может быть результатом давления отбора. Миллионы лет назад Бездна Челленджера была мельче, чем сейчас. За последние шесть-девять миллионов лет, когда Бездна Челленджера выросла до нынешней глубины, многие виды, присутствующие в отложениях этой древней биосферы, не смогли адаптироваться к возрастающему давлению воды и изменяющейся окружающей среде. Те виды, которые смогли адаптироваться, возможно, были предками организмов, в настоящее время эндемичных для Бездны Челленджера. [74]
Полихеты встречаются во всех океанах Земли на всех глубинах, от форм, обитающих в виде планктона у поверхности, до самых глубоких океанических впадин. 31 мая 2009 года роботизированный океанский зонд Nereus наблюдал экземпляр полихеты размером 2–3 см (пока не классифицированный) на дне Бездны Челленджера. [75] [78] [79] [80] Существует более 10 000 описанных видов полихеты; их можно найти практически в любой морской среде. Некоторые виды живут в океане с самой низкой температурой в хадаловой зоне, в то время как другие можно найти в чрезвычайно горячих водах, прилегающих к гидротермальным источникам.
В абиссальной и хадальной зонах области вокруг подводных гидротермальных источников и холодных просачиваний имеют наибольшую биомассу и биоразнообразие на единицу площади. Подпитываемые химическими веществами, растворенными в жерловых жидкостях, эти области часто являются домом для крупных и разнообразных сообществ термофильных , галофильных и других экстремофильных прокариотических микроорганизмов (таких как представители рода Beggiatoa , окисляющих сульфиды ), часто образующих большие бактериальные маты вблизи холода. просачивается. В этих местах хемосинтезирующие археи и бактерии обычно составляют основу пищевой цепи. Хотя процесс хемосинтеза является полностью микробным, эти хемосинтезирующие микроорганизмы часто поддерживают обширные экосистемы, состоящие из сложных многоклеточных организмов посредством симбиоза . [81] Эти сообщества характеризуются такими видами, как пузырчатые моллюски , митилиды , блюдечки , изоподы, гигантские трубчатые черви , мягкие кораллы , бельдюги , галатеидные крабы и креветки-альвинокариды . Самое глубокое сообщество сипа, обнаруженное на данный момент, находится в Японском желобе на глубине 7700 метров. [11]
Вероятно, наиболее важной экологической характеристикой абиссальных экосистем является ограничение энергии. Сообщества абиссального морского дна считаются ограниченными в питании , поскольку производство бентоса зависит от поступления обломочного органического материала , образующегося в эвфотической зоне, находящейся на тысячи метров над уровнем моря. [82] Большая часть органического потока поступает в виде ослабленного дождя из мелких частиц (обычно только 0,5–2% от чистой первичной продукции в эвфотической зоне), которая уменьшается обратно пропорционально глубине воды. [9] Поток мелких частиц может усиливаться за счет падения более крупных туш и переноса органического материала вниз по склону вблизи окраин континентов. [82]
Помимо высокого биоразнообразия, абиссальные равнины представляют большой текущий и будущий коммерческий и стратегический интерес. Например, их можно использовать для законной и незаконной утилизации крупных сооружений, таких как корабли и нефтяные вышки , радиоактивных отходов и других опасных отходов , таких как боеприпасы . Они также могут быть привлекательными местами для глубоководного рыболовства , добычи нефти, газа и других полезных ископаемых . Будущие мероприятия по глубоководному удалению отходов , которые могут стать значительными к 2025 году, включают размещение сточных вод и ила , секвестрацию углерода и утилизацию грунтов, извлеченных из дноуглубительных работ . [83]
Поскольку рыбные запасы в верхних слоях океана сокращаются, глубоководный промысел все чаще становится объектом эксплуатации. Поскольку глубоководные рыбы долгоживущие и медленно растут, этот глубоководный промысел не считается устойчивым в долгосрочной перспективе, учитывая нынешнюю практику управления. [83] Ожидается, что изменения в первичной продукции в фотической зоне изменят запасы в афотической зоне, ограниченной пищевыми продуктами.
Разведка углеводородов на глубоководных участках иногда приводит к значительному ухудшению состояния окружающей среды , главным образом из-за накопления загрязненного бурового шлама , а также из-за разливов нефти . Хотя выброс нефти , связанный с разливом нефти на платформе Deepwater Horizon в Мексиканском заливе, происходит из устья скважины , находящейся всего в 1500 метрах от поверхности океана, [84] тем не менее, он иллюстрирует тип экологической катастрофы , которая может возникнуть в результате неудач, связанных с морским бурением нефти. и газ.
Отложения некоторых абиссальных равнин содержат богатые минеральные ресурсы, особенно полиметаллические конкреции . Эти конкреции марганца, железа, никеля, кобальта и меди размером с картофелину, распространенные на морском дне на глубинах более 4000 метров [83] , представляют значительный коммерческий интерес. Район максимального коммерческого интереса для добычи полиметаллических конкреций (называемый Тихоокеанской конкреционной провинцией) находится в международных водах Тихого океана, простираясь от 118° до 157° и от 9° до 16° с.ш., площадь более 3 млн. км². [85] Абиссальная зона разлома Кларион-Клиппертон (CCFZ) представляет собой территорию в пределах Тихоокеанской конкреционной провинции, которая в настоящее время исследуется на предмет ее минерального потенциала. [61]
Восемь коммерческих подрядчиков в настоящее время имеют лицензию Международного органа по морскому дну ( межправительственной организации , созданной для организации и контроля всей деятельности, связанной с добычей полезных ископаемых в международной зоне морского дна за пределами национальной юрисдикции ) на разведку ресурсов конкреций и испытание методов добычи в восьми заявленных районах. , каждый площадью 150 000 км². [85] Когда в конечном итоге начнется добыча, каждая горнодобывающая операция, по прогнозам, будет непосредственно разрушать 300–800 км² морского дна в год и нарушать донную фауну на площади, в 5–10 раз превышающей ее площадь, из-за повторного отложения взвешенных отложений. Таким образом, в течение 15-летней прогнозируемой продолжительности одной операции по добыче добыча конкреций может серьезно повредить абиссальные сообщества морского дна на площади от 20 000 до 45 000 км² (зона размером не меньше Массачусетса ) . [85]
Ограниченные знания таксономии , биогеографии и естественной истории глубоководных сообществ не позволяют точно оценить риск исчезновения видов в результате крупномасштабной добычи полезных ископаемых. Данные, полученные в абиссальной части северной части Тихого океана и Северной Атлантики, позволяют предположить, что глубоководные экосистемы могут подвергаться негативному воздействию горнодобывающих операций в масштабах десятилетий. [83] В 1978 году драга на борту Hughes Glomar Explorer , управляемого американским горнодобывающим консорциумом Ocean Minerals Company (OMCO), проложила горный след на глубине 5000 метров в конкрециальных полях CCFZ. В 2004 году Французский научно-исследовательский институт эксплуатации моря ( IFREMER ) провел экспедицию Нодинаута по этому горному следу (который до сих пор виден на морском дне) для изучения долгосрочного воздействия этого физического нарушения на отложения и его донную фауну. . Взятые пробы поверхностного осадка показали, что его физические и химические свойства не восстановились с момента нарушения, произошедшего 26 лет назад. С другой стороны, биологическая активность, измеренная в треке приборами на борту пилотируемого подводного батискафа «Наутиле» , не отличалась от активности на близлежащем невозмущенном участке. Эти данные позволяют предположить, что донная фауна и потоки питательных веществ на границе вода–отложения полностью восстановились. [86]
{{cite book}}: |journal=игнорируется ( помощь ){{cite book}}: |journal=игнорируется ( помощь ){kind=link}