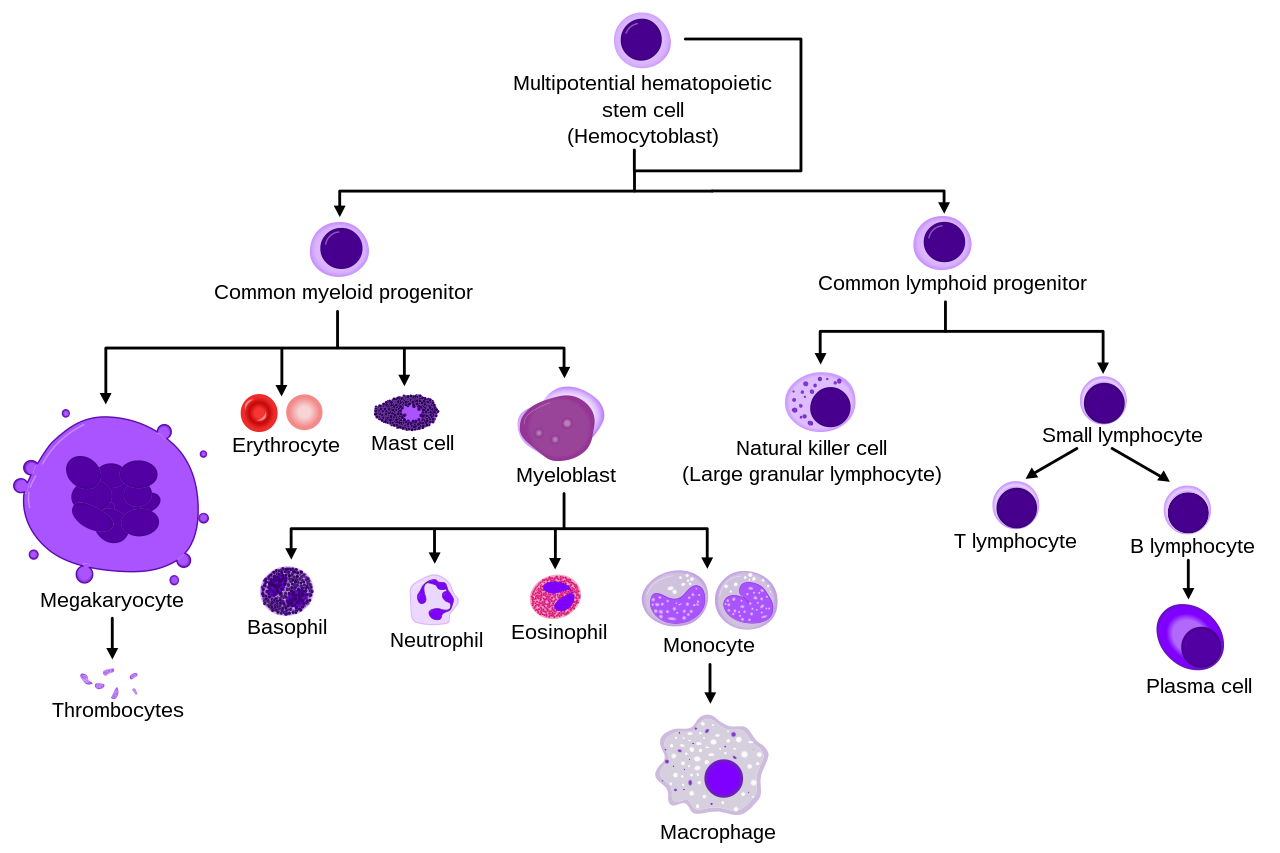
Диаграмма, показывающая развитие различных клеток крови от гемопоэтических стволовых клеток до зрелых клеток.
Гематопоэз ( / hɪ ˌ mætəpɔɪ ˈ iːsɪs , ˌ h iːmət oʊ -, ˌ hɛmə - / ; [1] [ 2] от древнегреческого αἷμα ( haîma ) « кровь » и ποιεῖν ( poieîn ) « делать»; также hematopoiesis в американском английском , иногда h(a)emopoiesis ) — это образование клеточных компонентов крови . Все клеточные компоненты крови происходят из гемопоэтических стволовых клеток . [3] У здорового взрослого человека примерно десять миллиардов ( 1010 ) до ста миллиардов (1011 ) новые клетки крови вырабатываются каждый день, чтобы поддерживать постоянный уровень в периферическом кровообращении. [4] [5] [ нужная страница ]
Процесс
Гемопоэтические стволовые клетки (ГСК)
Гемопоэтические стволовые клетки (ГСК) находятся в мозговом веществе кости ( костном мозге ) и обладают уникальной способностью давать начало всем различным типам зрелых клеток крови и тканям. [3] ГСК являются самообновляющимися клетками: когда они дифференцируются, по крайней мере некоторые из их дочерних клеток остаются как ГСК, поэтому пул стволовых клеток не истощается. [6] Это явление называется асимметричным делением. [7] Другие дочерние клетки ГСК ( миелоидные и лимфоидные клетки-предшественники) могут следовать любым другим путям дифференциации, которые приводят к производству одного или нескольких определенных типов клеток крови, но не могут обновляться. Пул предшественников неоднороден и может быть разделен на две группы: долгосрочные самообновляющиеся ГСК и только временно самообновляющиеся ГСК, также называемые краткосрочными. [8] Это один из основных жизненно важных процессов в организме.
Типы клеток
Все клетки крови делятся на три линии. [9]
Красные кровяные клетки , которые также называются эритроцитами, являются клетками, переносящими кислород . Эритроциты функциональны и выбрасываются в кровь. Количество ретикулоцитов, которые являются незрелыми эритроцитами, дает оценку скорости эритропоэза .
Гранулопоэз (или гранулоцитопоэз) – это кроветворение гранулоцитов, за исключением тучных клеток , которые являются гранулоцитами, но с экстрамедуллярным созреванием. [10]
В период с 1948 по 1950 год Комитет по уточнению номенклатуры клеток и болезней крови и кроветворных органов выпустил доклады по номенклатуре клеток крови. [11] [12] Обзор терминологии представлен ниже, от самой ранней до последней стадии развития: [ необходима ссылка ]
[корень]взрыв
про[корневой]цит
[корень]цит
мета[корневой]цит
название зрелой клетки
Корень для эритроцитарных колониеобразующих единиц (CFU-E) - "rubri", для гранулоцитарно-моноцитарных колониеобразующих единиц (CFU-GM) - "granulo" или "myelo" и "mono", для лимфоцитарных колониеобразующих единиц (CFU-L) - "lympho", а для мегакариоцитарных колониеобразующих единиц (CFU-Meg) - "megakaryo". Согласно этой терминологии, стадии формирования эритроцитов будут следующими: рубрибласт, прорубрицит, рубрицит, метарубрицит и эритроцит. Однако в настоящее время наиболее распространенной представляется следующая номенклатура:
Остеокласты также возникают из кроветворных клеток моноцитарно-нейтрофильной линии, в частности КОЕ-ГМ.
Расположение
Места кроветворения (у человека) в пре- и постнатальный периоды
В развивающихся эмбрионах кроветворение происходит в агрегатах клеток крови в желточном мешке, называемых кровяными островками . По мере развития кроветворение происходит в селезенке , печени и лимфатических узлах . [13] Когда развивается костный мозг , он в конечном итоге берет на себя задачу формирования большинства клеток крови для всего организма. [3] Однако созревание, активация и некоторая пролиферация лимфоидных клеток происходит в селезенке, тимусе и лимфатических узлах. У детей кроветворение происходит в костном мозге длинных костей, таких как бедренная и большеберцовая кость. У взрослых это происходит в основном в тазу, черепе, позвонках и грудине. [14]
Экстрамедуллярный
В некоторых случаях печень, тимус и селезенка могут возобновить свою кроветворную функцию, если это необходимо. Это называется экстрамедуллярным кроветворением . Это может привести к существенному увеличению размеров этих органов. Во время внутриутробного развития, поскольку кости и, следовательно, костный мозг развиваются позже, печень функционирует как основной кроветворный орган. Поэтому печень увеличивается во время развития. [15] Экстрамедуллярный кроветворение и миелопоэз могут поставлять лейкоциты при сердечно-сосудистых заболеваниях и воспалениях во взрослом возрасте. [16] [17] Селезеночные макрофаги и молекулы адгезии могут участвовать в регуляции экстрамедуллярной генерации миелоидных клеток при сердечно-сосудистых заболеваниях . [18] [19]
Созревание
Более подробная и полная диаграмма, показывающая развитие различных клеток крови у человека:
Морфологические характеристики кроветворных клеток показаны в соответствии с окраской по Райту, Маю-Гимзе или Маю-Грюнвальду-Гимзе. Альтернативные названия некоторых клеток указаны в скобках.
Некоторые клетки могут иметь более одного характерного внешнего вида. В этих случаях было включено более одного представления одной и той же клетки.
Моноциты и лимфоциты вместе составляют агранулоциты, в отличие от гранулоцитов (базофилов, нейтрофилов и эозинофилов), которые вырабатываются в процессе гранулопоэза.
B., N. и E. обозначают базофильный, нейтрофильный и эозинофильный соответственно – как в базофильном промиелоците. Для лимфоцитов фактическими обозначениями являются T и B.
Полихроматический эритроцит (ретикулоцит) справа демонстрирует свой характерный вид при окраске метиленовым синим или азуру В.
Эритроцит справа более точно отражает его внешний вид в реальности, если рассматривать его под микроскопом.
Другие клетки, возникающие из моноцитов: остеокласты, микроглия (центральная нервная система), клетки Лангерганса (эпидермис), клетки Купфера (печень).
Для ясности Т- и В-лимфоциты разделены, чтобы лучше показать, что плазматическая клетка возникает из В-клетки. Обратите внимание, что нет никакой разницы во внешнем виде В- и Т-клеток, если не применяется специфическое окрашивание.
По мере созревания стволовой клетки она претерпевает изменения в экспрессии генов , которые ограничивают типы клеток, которыми она может стать, и приближают ее к определенному типу клеток ( клеточная дифференциация ). Эти изменения часто можно отслеживать, контролируя наличие белков на поверхности клетки. Каждое последующее изменение приближает клетку к конечному типу клеток и еще больше ограничивает ее потенциал стать другим типом клеток. [ необходима цитата ]
Определение судьбы клетки
Были предложены две модели кроветворения: детерминизм и стохастическая теория. [20] Для стволовых клеток и других недифференцированных клеток крови в костном мозге детерминизм обычно объясняется теорией детерминизма кроветворения, утверждающей, что колониестимулирующие факторы и другие факторы кроветворной микросреды определяют, что клетки следуют определенному пути клеточной дифференциации. [3] Это классический способ описания кроветворения. В стохастической теории недифференцированные клетки крови дифференцируются в определенные типы клеток случайным образом. Эта теория была подтверждена экспериментами, показывающими, что в популяции мышиных кроветворных клеток-предшественников, лежащая в основе стохастическая изменчивость в распределении Sca-1 , фактора стволовых клеток , подразделяет популяцию на группы, демонстрирующие различные скорости клеточной дифференциации . Например, под влиянием эритропоэтина (фактора дифференцировки эритроцитов) субпопуляция клеток (определяемая по уровням Sca-1) дифференцировалась в эритроциты в семь раз быстрее, чем остальная часть популяции. [21] Кроме того, было показано, что если позволить этой субпопуляции расти, она восстанавливала исходную субпопуляцию клеток, что подтверждает теорию о том, что это стохастический, обратимый процесс. Другой уровень, на котором может быть важна стохастичность, — это процесс апоптоза и самообновления. В этом случае гемопоэтическая микросреда заставляет некоторые клетки выживать, а некоторые, с другой стороны, выполнять апоптоз и умирать. [3] Регулируя этот баланс между различными типами клеток, костный мозг может изменять количество различных клеток, которые в конечном итоге будут произведены. [22]
Производство красных и белых кровяных клеток регулируется с большой точностью у здоровых людей, а производство лейкоцитов быстро увеличивается во время инфекции. Пролиферация и самообновление этих клеток зависят от факторов роста. Одним из ключевых игроков в самообновлении и развитии гемопоэтических клеток является фактор стволовых клеток (SCF), [25] , который связывается с рецептором c-kit на HSC. Отсутствие SCF является летальным. Существуют и другие важные факторы роста гликопротеинов , которые регулируют пролиферацию и созревание, такие как интерлейкины IL-2 , IL-3 , IL-6 , IL-7 . Другие факторы, называемые колониестимулирующими факторами (CSF), специфически стимулируют производство комитированных клеток. Три CSF — это гранулоцитарно-макрофагальный CSF (GM-CSF), гранулоцитарный CSF (G-CSF) и макрофагальный CSF (M-CSF). [26] Они стимулируют образование гранулоцитов и воздействуют либо на клетки-предшественники , либо на клетки-конечные продукты. [ необходима цитата ]
Эритропоэтин необходим для того, чтобы миелоидная клетка-предшественник стала эритроцитом. [23] С другой стороны, тромбопоэтин заставляет миелоидные клетки-предшественники дифференцироваться в мегакариоциты ( клетки, образующие тромбоциты ). [23] На диаграмме справа приведены примеры цитокинов и дифференцированных клеток крови, которые они дают. [27]
Факторы транскрипции
Факторы роста инициируют пути передачи сигнала , которые приводят к активации факторов транскрипции . Факторы роста вызывают различные результаты в зависимости от комбинации факторов и стадии дифференциации клетки. Например, долгосрочная экспрессия PU.1 приводит к миелоидному обязательству, а краткосрочная индукция активности PU.1 приводит к образованию незрелых эозинофилов. [28] Недавно было сообщено, что факторы транскрипции, такие как NF-κB, могут регулироваться микроРНК (например, miR-125b) в гемопоэзе. [29]
Первым ключевым игроком дифференциации от HSC до мультипотентного предшественника (MPP) является транскрипционный фактор CCAAT-энхансер связывающий белок α ( C/EBP α). Мутации в C/EBPα связаны с острым миелоидным лейкозом . [30] С этого момента клетки могут дифференцироваться либо по эритроидно-мегакариоцитарной линии, либо по лимфоидной и миелоидной линии, которые имеют общего предшественника, называемого лимфоидно-примированным мультипотентным предшественником. Существует два основных фактора транскрипции. PU.1 для эритроидно-мегакариоцитарной линии и GATA-1 , который приводит к лимфоидно-примированному мультипотентному предшественнику. [31]
Другие факторы транскрипции включают Ikaros [32] ( развитие B-клеток ) и Gfi1 [33] (стимулирует развитие Th2 и подавляет Th1) или IRF8 [34] ( базофилы и тучные клетки ). Важно отметить, что некоторые факторы вызывают разные реакции на разных стадиях кроветворения. Например, CEBPα в развитии нейтрофилов или PU.1 в развитии моноцитов и дендритных клеток. Важно отметить, что процессы не являются однонаправленными: дифференцированные клетки могут восстанавливать свойства клеток-предшественников. [1]
Примером является фактор PAX5 , который важен для развития В-клеток и связан с лимфомами. [35] Удивительно, но мыши с условным нокаутом гена pax5 позволили периферическим зрелым В-клеткам дедифференцироваться в ранние костномозговые предшественники. Эти результаты показывают, что факторы транскрипции действуют как смотрители уровня дифференциации, а не только как инициаторы. [36]
У некоторых позвоночных кроветворение может происходить везде, где есть рыхлая строма соединительной ткани и медленное кровоснабжение, например, в кишечнике , селезенке или почках . [40]
В отличие от плацентарных млекопитающих, печень новорожденных сумчатых активно кроветворна. [41] [42] [43] [44]
^ abcde Бирбрейр, Александр; Френетт, Пол С. (1 марта 2016 г.). «Нишевая гетерогенность в костном мозге». Анналы Нью-Йоркской академии наук . 1370 (1): 82–96. Bibcode : 2016NYASA1370...82B. doi : 10.1111/nyas.13016. ISSN 1749-6632. PMC 4938003. PMID 27015419 .
^ Медицинские лекции 4-го семестра в Университете Уппсалы 2008 г., автор Лейф Янссон
^ Монга И, Каур К, Дханда С (март 2022 г.). «Возвращаясь к гемопоэзу: применение транскриптомики массовых и отдельных клеток для анализа транскрипционной гетерогенности в гемопоэтических стволовых клетках». Briefings in Functional Genomics . 21 (3): 159–176. doi : 10.1093/bfgp/elac002. PMID 35265979.
^ Моррисон, Дж.; Джудит Кимбл (2006). «Асимметричные и симметричные деления стволовых клеток в развитии и раке» (PDF) . Nature . 441 (7097): 1068–74. Bibcode :2006Natur.441.1068M. doi :10.1038/nature04956. hdl : 2027.42/62868 . PMID 16810241. S2CID 715049.
^ Morrison SJ, Weissman IL (ноябрь 1994). «Долгосрочное повторное заселение подмножества гемопоэтических стволовых клеток детерминировано и изолируется по фенотипу». Immunity . 1 (8): 661–73. doi :10.1016/1074-7613(94)90037-x. PMID 7541305.
^ "Гематопоэз из плюрипотентных стволовых клеток". Библиотека ресурсов антител . ThermoFisher Scientific . Получено 25 апреля 2020 г.
^ Малер (2013). Хашек, Ванда; Руссо, Колин Г.; Уоллиг, Мэтью А. (ред.). Справочник Хашека и Руссо по токсикологической патологии . Ассоциированные редакторы: Брэд Болон и Рикардо Очоа; редактор иллюстраций: Бет У. (третье изд.). [Sl]: Academic Press. стр. 1863. ISBN978-0-12-415759-0.
^ "ПЕРВЫЙ отчет комитета по разъяснению номенклатуры клеток и заболеваний крови и кроветворных органов" . Американский журнал клинической патологии . 18 (5): 443–50. Май 1948. doi :10.1093/ajcp/18.5_ts.443. PMID 18913573.
^ "ТРЕТИЙ, четвертый и пятый отчеты комитета по уточнению номенклатуры клеток и заболеваний крови и кроветворных органов" . Американский журнал клинической патологии . 20 (6): 562–79. Июнь 1950. doi :10.1093/ajcp/20.6.562. PMID 15432355.
^ Fernández KS, de Alarcón PA (декабрь 2013 г.). «Развитие кроветворной системы и нарушения кроветворения, проявляющиеся в младенчестве и раннем детстве» . Pediatric Clinics of North America . 60 (6): 1273–89. doi :10.1016/j.pcl.2013.08.002. PMID 24237971.
^ Георгиадес К.С., Нейман Э.Г., Фрэнсис И.Р., Снайдер М.Б., Фишман Е.К. (ноябрь 2002 г.). «Типичные и атипичные проявления экстрамедуллярного кроветворения». АЖР. Американский журнал рентгенологии . 179 (5): 1239–43. дои : 10.2214/ajr.179.5.1791239 . ПМИД 12388506.
^ Swirski, Filip K.; Libby, Peter; Aikawa, Elena; Alcaide, Pilar; Luscinskas, F. William; Weissleder, Ralph; Pittet, Mikael J. (2 января 2007 г.). «Ly-6Chi-моноциты доминируют в моноцитозе, связанном с гиперхолестеринемией, и приводят к образованию макрофагов в атеромах». Journal of Clinical Investigation . 117 (1): 195–205. doi : 10.1172/JCI29950 . PMC 1716211 . PMID 17200719.
^ Swirski FK, Nahrendorf M, Etzrodt M, Wildgruber M, Cortez-Retamozo V, Panizzi P, Figueiredo JL, Kohler RH, Chudnovskiy A, Waterman P, Aikawa E, Mempel TR, Libby P, Weissleder R, Pittet MJ (30 июля 2009 г.). «Идентификация моноцитов селезеночного резервуара и их размещение в очагах воспаления». Science . 325 (5940): 612–616. Bibcode :2009Sci...325..612S. doi :10.1126/science.1175202. PMC 2803111 . PMID 19644120.
^ Dutta, P; Hoyer, FF; Grigoryeva, LS; Sager, HB; Leuschner, F; Courties, G; Borodovsky, A; Novobrantseva, T; Ruda, VM; Fitzgerald, K; Iwamoto, Y; Wojtkiewicz, G; Sun, Y; Da Silva, N; Libby, P; Anderson, DG; Swirski, FK; Weissleder, R; Nahrendorf, M (6 апреля 2015 г.). «Макрофаги сохраняют гемопоэтические стволовые клетки в селезенке через VCAM-1». Журнал экспериментальной медицины . 212 (4): 497–512. doi : 10.1084/jem.20141642 . PMC 4387283. PMID 25800955 .
^ Dutta, P; Hoyer, FF; Sun, Y; Iwamoto, Y; Tricot, B; Weissleder, R; Magnani, JL; Swirski, FK; Nahrendorf, M (сентябрь 2016 г.). «Ингибирование E-селектина смягчает активацию HSC селезенки и миелопоэз у мышей с гиперхолестеринемией и инфарктом миокарда». Артериосклероз, тромбоз и сосудистая биология . 36 (9): 1802–8. doi : 10.1161/ATVBAHA.116.307519 . PMC 5001901. PMID 27470513.
^ Киммел, Марек (1 января 2014 г.). «Стохастичность и детерминизм в моделях гемопоэза». Системный биологический подход к крови . Достижения в экспериментальной медицине и биологии. Том 844. С. 119–152. doi :10.1007/978-1-4939-2095-2_7. ISBN978-1-4939-2094-5. ISSN 0065-2598. PMID 25480640.
^ Чанг, Ханна Х.; Хемберг, Мартин; Барахона, Маурисио; Ингбер, Дональд Э.; Хуан, Суй (2008). «Транскриптомный шум контролирует выбор линии в клетках-предшественниках млекопитающих». Nature . 453 (7194): 544–547. Bibcode :2008Natur.453..544C. doi :10.1038/nature06965. PMC 5546414 . PMID 18497826.
^ Alenzi, FQ; Alenazi, BQ; Ahmad, SY; Salem, ML; Al-Jabri, AA; Wyse, RK (март 2009 г.). «Гемопоэтическая стволовая клетка: между апоптозом и самообновлением». Йельский журнал биологии и медицины . 82 (1): 7–18. PMC 2660591. PMID 19325941 .
^ abcd Молекулярная клеточная биология. Lodish, Harvey F. 5-е изд.: – Нью-Йорк: WH Freeman and Co., 2003, 973 с. билл. ISBN 0-7167-4366-3Lodish H, Berk A, Zipursky SL, Matsudaira P, Baltimore D, Darnell J (2000). «Рак возникает в пролиферирующих клетках». Молекулярная клеточная биология (4-е изд.). Нью-Йорк: WH Freeman. Рисунок 24-8: Формирование дифференцированных клеток крови из гемопоэтических стволовых клеток в костном мозге. ISBN 0-7167-3136-3– через книжную полку NCBI.
^ Род Флауэр; Хамфри П. Ранг; Морин М. Дейл; Риттер, Джеймс М. (2007). Фармакология Ранга и Дейла . Эдинбург: Черчилль Ливингстон. ISBN978-0-443-06911-6.
^ Broudy, VC (15 августа 1997 г.). «Фактор стволовых клеток и гемопоэз». Blood . 90 (4): 1345–64. doi : 10.1182/blood.V90.4.1345 . PMID 9269751.
^ Ketley, NJ; AC Newland (1997). «Гемопоэтические факторы роста». Postgrad Med J . 73 (858): 215–221. doi :10.1136/pgmj.73.858.215. PMC 2431295 . PMID 9156123.
^ Хауке, Ральф; Стефано Р. Тарантоло (ноябрь 2000 г.). «Гематопоэтические факторы роста». Лабораторная медицина . 31 (11): 613–5. doi : 10.1309/HNTM-ELUV-AV9G-MA1P .
^ Энгель, И; Мюрре, К (октябрь 1999 г.). «Транскрипционные факторы в гемопоэзе» . Current Opinion in Genetics & Development . 9 (5): 575–9. doi :10.1016/s0959-437x(99)00008-8. PMID 10508690.
^ Ho, PA; Alonzo, TA; Gerbing, RB; Pollard, J; Stirewalt, DL; Hurwitz, C; Heerema, NA; Hirsch, B; Raimondi, SC; Lange, B; Franklin, JL; Radich, JP; Meshinchi, S (25 июня 2009 г.). «Распространенность и прогностические последствия мутаций CEBPA при остром миелоидном лейкозе (ОМЛ) у детей: отчет Детской онкологической группы». Blood . 113 (26): 6558–66. doi :10.1182/blood-2008-10-184747. PMC 2943755 . PMID 19304957.
^ Woolthuis, Carolien M.; Park, Christopher Y. (10 марта 2016 г.). «Приверженность гемопоэтических стволовых/прогениторных клеток линии мегакариоцитов». Blood . 127 (10): 1242–1248. doi : 10.1182/blood-2015-07-607945 . ISSN 0006-4971. PMC 5003506 . PMID 26787736. S2CID 206939258.
^ Томпсон, Элизабет К.; Кобб, Брэдли С.; Саббаттини, Пьерангела; Мейкслспергер, Соня; Парельо, Вания; Либерг, Дэвид; Тейлор, Бенджамин; Диллон, Ниалл; Георгопулос, Катя (1 марта 2007 г.). «ДНК-связывающие белки Ikaros как неотъемлемые компоненты регуляторных цепей, специфичных для стадий развития В-клеток». Иммунитет . 26 (3): 335–344. doi : 10.1016/j.immuni.2007.02.010 . ISSN 1074-7613. PMID 17363301.
^ Сасаки, Харука; Куротаки, Дайсуке; Тамура, Томохико (1 апреля 2016 г.). «Регуляция развития базофилов и тучных клеток факторами транскрипции». Allergology International . 65 (2): 127–134. doi : 10.1016/j.alit.2016.01.006 . ISSN 1440-1592. PMID 26972050.
^ O'Brien, P; Morin, P Jr; Ouellette, RJ; Robichaud, GA (15 декабря 2011 г.). «Ген Pax-5: плюрипотентный регулятор дифференцировки В-клеток и раковых заболеваний». Cancer Research . 71 (24): 7345–50. doi : 10.1158/0008-5472.CAN-11-1874 . PMID 22127921.
^ Cobaleda, C; Jochum, W; Busslinger, M (27 сентября 2007 г.). «Преобразование зрелых В-клеток в Т-клетки путем дедифференциации в некоммитированные предшественники» . Nature . 449 (7161): 473–7. Bibcode : 2007Natur.449..473C. doi : 10.1038/nature06159. PMID 17851532. S2CID 4414856.
^ Ван, Дж. Х.; Нихогианнопулу, А.; Ву, Л.; Сан, Л.; Шарп, А. Х.; Бигби, М.; Георгопулос, К. (декабрь 1996 г.). «Избирательные дефекты в развитии фетальной и взрослой лимфоидной системы у мышей с нулевой мутацией Ikaros». Immunity . 5 (6): 537–49. doi : 10.1016/s1074-7613(00)80269-1 . PMID 8986714.
^ Сан, Л.; Лю, А.; Георгопулос, К. (1 октября 1996 г.). «Взаимодействия белков, опосредованные цинковым пальцем, модулируют активность Икароса, молекулярный контроль развития лимфоцитов». Журнал EMBO . 15 (19): 5358–69. doi :10.1002/j.1460-2075.1996.tb00920.x. PMC 452279. PMID 8895580 .
^ Schjerven, H; McLaughlin, J; Arenzana, TL; Frietze, S; Cheng, D; Wadsworth, SE; Lawson, GW; Bensinger, SJ; Farnham, PJ; Witte, ON; Smale, ST (октябрь 2013 г.). «Избирательная регуляция лимфопоэза и лейкемогенеза отдельными цинковыми пальцами Икароса». Nature Immunology . 14 (10): 1073–83. doi :10.1038/ni.2707. PMC 3800053 . PMID 24013668.
^ Зон, ЛИ (15 октября 1995 г.). «Биология развития кроветворения». Кровь (обзор). 86 (8): 2876–91. doi : 10.1182/blood.V86.8.2876.2876 . PMID 7579378.
^ Old JM (2016). Гемопоэз у сумчатых. Развитие и сравнительная иммунология. 58, 40-46. DOI: 10.1016/j.dci.2015.11.009
^ Old JM, Deane EM (2000). Развитие иммунной системы и иммунологическая защита у детенышей сумчатых сумчатых. Developmental and Comparative Immunology. 24(5), 445-454. DOI: 10.1016/S0145-305X(00)00008-2
^ Old JM, Deane EM (2003). Лимфоидные и иммуногематопоэтические ткани эмбрионального щеткохвостого опоссума ( Trichosurus vulpecula ). Анатомия и эмбриология (теперь называется «Структура и функции мозга»). 206(3), 193-197. DOI: 10.1007/s00429-002-0285-2
^ Old JM, Selwood L , Deane EM (2003). Гистологическое исследование лимфоидных и иммуногематопоэтических тканей взрослого полосатого даннарта ( Sminthopsis macroura ). Клетки Ткани Органы. 173(2), 115-121. DOI: 10.1159/000068946
Дальнейшее чтение
Годин, Изабель; Кумано, Ана, ред. (2006). Развитие гемопоэтических стволовых клеток. Springer. ISBN 978-0-306-47872-7.
Внешние ссылки
В Scholia есть тематический профиль по теме «Гемопоэз» .

_diagram_en.svg/1280px-Hematopoiesis_(human)_diagram_en.svg.png)
