
Насекомые — единственная группа беспозвоночных , у которой развились крылья и способность летать . Насекомые впервые полетели в каменноугольном периоде , примерно 300–350 миллионов лет назад, что сделало их первыми животными, развившими способность летать. Крылья, возможно, развились из придатков по бокам существующих конечностей, у которых уже были нервы, суставы и мышцы, используемые для других целей. Первоначально они могли использоваться для плавания по воде или для замедления скорости спуска при планировании.
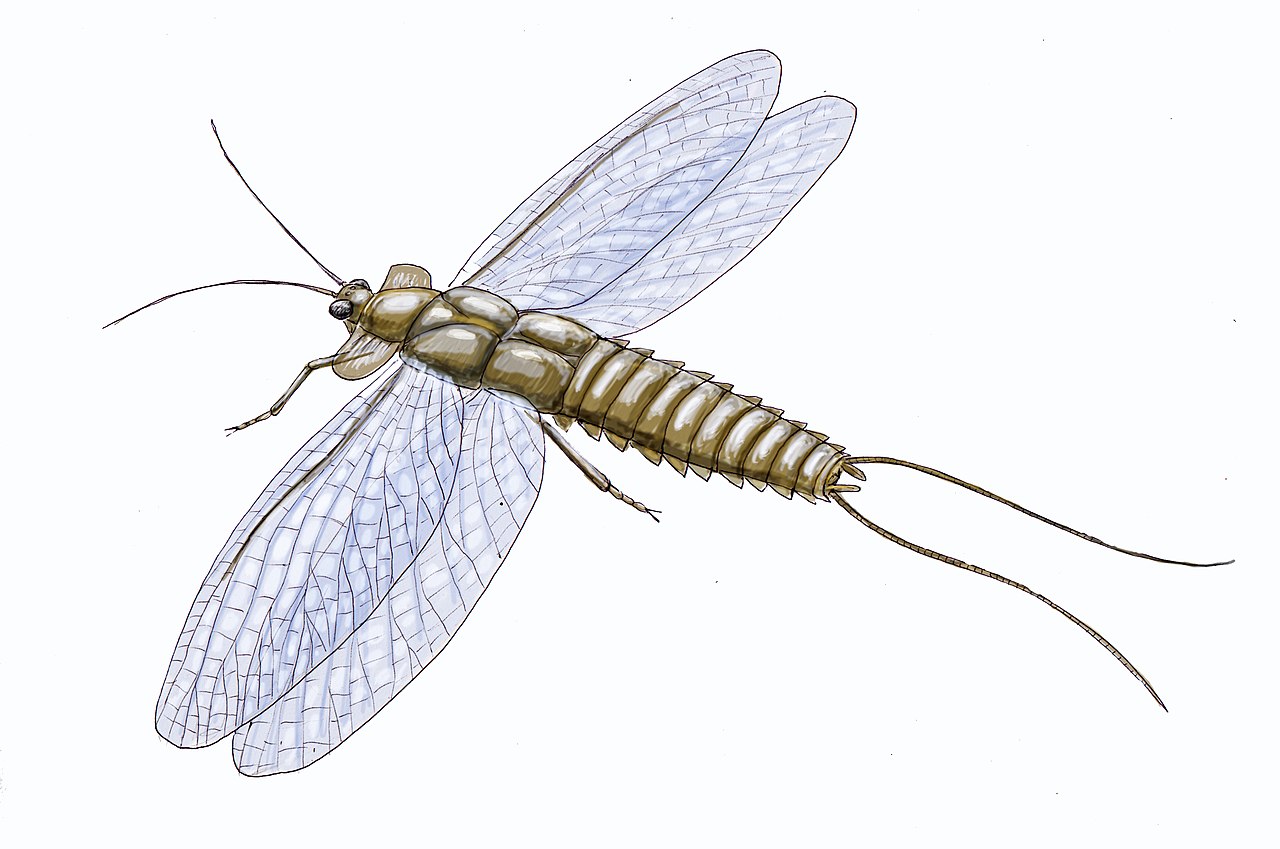
Две группы насекомых, стрекозы и подёнки , имеют летательные мышцы, прикрепленные непосредственно к крыльям. У других крылатых насекомых к грудной клетке прикрепляются летательные мышцы, которые заставляют ее колебаться, заставляя крылья биться. Из этих насекомых некоторые ( мухи и некоторые жуки ) достигают очень высоких частот взмахов крыльев благодаря развитию «асинхронной» нервной системы, в которой грудная клетка колеблется быстрее, чем частота нервных импульсов.
Не все насекомые способны летать. Ряд бескрылых насекомых вторично потеряли свои крылья в ходе эволюции , в то время как другие более базальные насекомые, такие как чешуйница, крылья так и не развились. У некоторых эусоциальных насекомых, таких как муравьи и термиты , только крылатые репродуктивные касты развивают крылья во время брачного сезона , а затем сбрасывают их после спаривания, в то время как представители других каст остаются бескрылыми всю свою жизнь.
Некоторые очень маленькие насекомые используют не стационарную аэродинамику , а механизм Вейса-Фога, создающий большие подъемные силы за счет износа крыльев. Многие насекомые умеют парить, сохраняя высоту и контролируя свое положение. У некоторых насекомых, таких как мотыльки , передние крылья соединены с задними, поэтому они могут работать в унисон.
В отличие от других насекомых, мышцы крыльев Ephemeroptera (подёнки) и Odonata (стрекозы и стрекозы) прикрепляются непосредственно к основаниям крыльев, которые шарнирно соединены так, что небольшое движение основания крыла вниз поднимает само крыло вверх, подобно греблю через воздух. У стрекоз и стрекоз передние и задние крылья схожи по форме и размеру. Каждый из них действует независимо, что дает определенную степень тонкого контроля и мобильности с точки зрения резкости, с которой они могут менять направление и скорость, чего не наблюдается у других летающих насекомых. Все стрекозы — воздушные хищники и всегда охотились на других насекомых, передающихся по воздуху. [1]

За исключением двух отрядов с мышцами прямого полета, все остальные живые крылатые насекомые летают, используя другой механизм, задействующий мышцы непрямого полета. Этот механизм возник однажды и является определяющим признаком ( синапоморфией ) инфракласса Neoptera ; это соответствует, вероятно, не случайно, появлению механизма складывания крыльев, который позволяет насекомым-неоптеранам складывать крылья обратно над брюшком в состоянии покоя (хотя эта способность вторично утеряна у некоторых групп, например у бабочек ) . . [1]
Однако всех Neoptera объединяет то, как работают мышцы грудной клетки : эти мышцы вместо того, чтобы прикрепляться к крыльям, прикрепляются к грудной клетке и деформируют ее; поскольку крылья являются продолжением грудного экзоскелета , деформации грудной клетки также заставляют крылья двигаться. Набор продольных мышц спины сжимает грудную клетку спереди назад, заставляя дорсальную поверхность грудной клетки ( notum ) выгибаться вверх, заставляя крылья переворачиваться вниз. Другая группа мышц, идущая от тергума к грудине , снова тянет нотум вниз, заставляя крылья подниматься вверх. [1] [2]

Насекомые, которые взмахивают крыльями менее ста раз в секунду, используют синхронные мышцы. Синхронная мышца — это тип мышцы, которая сокращается один раз при каждом нервном импульсе. Обычно это производит меньшую мощность и менее эффективно, чем асинхронные мышцы, что объясняет независимую эволюцию асинхронных полетных мышц в нескольких отдельных кладах насекомых. [3]
Насекомые, которые машут крыльями быстрее, например шмели , используют асинхронные мышцы; это тип мышцы, которая сокращается более одного раза за нервный импульс. Это достигается за счет стимуляции повторного сокращения мышцы за счет снятия напряжения в мышце, что может произойти быстрее, чем при простой стимуляции нерва. [4] Это позволяет частоте взмахов крыльев превышать скорость, с которой нервная система может посылать импульсы. Асинхронная мышца — одно из последних усовершенствований, появившихся у некоторых высших неоптовых ( жесткокрылых , двукрылых и перепончатокрылых ). Общий эффект заключается в том, что многие высшие Neoptera могут взмахивать крыльями гораздо быстрее, чем насекомые с мышцами прямого полета. [1]
Асинхронные мышцы по определению находятся под относительно грубым контролем со стороны нервной системы. Чтобы сбалансировать этот эволюционный компромисс, насекомые, которые развили непрямой полет, также развили отдельную нервно-мышечную систему для точного контроля взмахов крыльев. [5] Эти мышцы, известные как «прямые мышцы», прикрепляются непосредственно к склеритам, образующим шарнир крыла, и сокращаются под действием импульсов 1:1 от мотонейронов. [6] Недавние работы начали изучать сложную нелинейную мышечную динамику шарнира крыла и ее влияние на траекторию законцовки крыла. [7]
Существует две основные аэродинамические модели полета насекомых: создание вихря на передней кромке и использование хлопка и броска. [8] [9]
Большинство насекомых используют метод, который создает спиральный вихрь на передней кромке . Эти машущие крылья совершают два основных полухода. Ход вниз начинается вверх и назад, а затем погружается вниз и вперед. Затем крыло быстро переворачивают ( супинация ) так, чтобы передняя кромка была направлена назад. Затем ход вверх толкает крыло вверх и назад. Затем крыло снова переворачивается ( пронация ), и может произойти еще один ход вниз. Диапазон частот у насекомых с мышцами синхронного полета обычно составляет от 5 до 200 герц (Гц). У лиц с асинхронными летательными мышцами частота взмахов крыльев может превышать 1000 Гц. Когда насекомое зависает, оба взмаха занимают одинаковое время. Однако более медленное движение вниз обеспечивает тягу . [8] [9]
Идентификация основных сил имеет решающее значение для понимания полета насекомых. Первые попытки понять машущие крылья предполагали квазистационарное состояние. Это означает, что поток воздуха над крылом в любой момент времени предполагался таким же, как поток воздуха над немашущим, установившимся крылом при том же угле атаки. Разделив машущее крыло на большое количество неподвижных положений и затем проанализировав каждое положение, можно было бы создать временную шкалу мгновенных сил, действующих на крыло в каждый момент. Рассчитанная подъемная сила оказалась слишком маленькой в три раза, поэтому исследователи поняли, что должны существовать нестационарные явления, создающие аэродинамические силы. Было несколько разрабатываемых аналитических моделей, пытающихся аппроксимировать поток, близкий к машущему крылу. Некоторые исследователи предсказывали пики силы при супинации. С помощью динамически масштабируемой модели плодовой мухи эти предсказанные силы позже были подтверждены. Другие утверждали, что пики силы во время супинации и пронации вызваны неизвестным вращательным эффектом, который фундаментально отличается от поступательного явления. С этим аргументом есть некоторое несогласие. С помощью вычислительной гидродинамики некоторые исследователи утверждают, что эффекта вращения нет. Они утверждают, что высокие силы вызваны взаимодействием со следом, оставленным предыдущим ударом. [8] [9]
Подобно эффекту вращения, упомянутому выше, явления, связанные с взмахами крыльев, до конца не поняты и не согласованы. Поскольку каждая модель является приближением, разные модели не учитывают эффекты, которыми можно пренебречь. Например, эффект Вагнера , предложенный Гербертом А. Вагнером в 1925 году [10], говорит о том, что циркуляция медленно возрастает до установившегося состояния из-за вязкости, когда наклонное крыло ускоряется из состояния покоя. Это явление могло бы объяснить значение подъемной силы, которое меньше прогнозируемого. Обычно речь идет о поиске источников дополнительного подъема. Утверждалось, что этот эффект незначителен для потока с числом Рейнольдса , типичным для полета насекомых. Число Рейнольдса является мерой турбулентности ; Течение ламинарное (плавное), когда число Рейнольдса низкое, и турбулентное, когда оно велико. [11] Эффект Вагнера сознательно игнорировался, по крайней мере, в одной модели. [9] Одним из наиболее важных явлений, происходящих во время полета насекомых, является всасывание передней кромки. Эта сила важна для расчета эффективности. Концепция всасывания на передней кромке впервые была предложена Д.Г. Эллисом и Дж.Л. Столлери в 1988 году для описания вихревой подъемной силы на треугольных крыльях с острыми краями . [12] При больших углах атаки поток отделяется по передней кромке, но снова присоединяется, не достигая задней кромки. Внутри этого пузыря разделенного потока находится вихрь. Поскольку угол атаки очень велик, большая часть импульса передается потоку вниз. Эти две особенности создают большую подъемную силу, а также некоторое дополнительное сопротивление. Однако важной особенностью является лифт. Поскольку поток отделился, но все же обеспечивает большую подъемную силу, это явление называется задержкой сваливания , впервые замеченное на воздушных винтах Х. Химмельскампом в 1945 году. [13] Этот эффект наблюдался при полете машущих насекомых, и было доказано, что он быть способным обеспечить достаточную подъемную силу, чтобы компенсировать недостатки квазистационарных моделей. Этот эффект используется каноистами при гребле на гребле . [8] [9]
Все воздействия на машущее крыло можно свести к трем основным источникам аэродинамических явлений: вихрю на передней кромке, установившимся аэродинамическим силам на крыле и контакту крыла со следом от предыдущих ударов. Размер летающих насекомых колеблется от примерно 20 микрограмм до примерно 3 граммов. По мере увеличения массы тела насекомого площадь крыльев увеличивается, а частота взмахов крыльев уменьшается. Для более крупных насекомых число Рейнольдса (Re) может достигать 10 000, при этом поток начинает становиться турбулентным. Для более мелких насекомых оно может достигать 10. Это означает, что вязкие эффекты гораздо важнее для более мелких насекомых. [9] [14] [15]
Еще одна интересная особенность полета насекомых – наклон тела. По мере увеличения скорости полета тело насекомого имеет тенденцию наклоняться носом вниз и становиться более горизонтальным. Это уменьшает лобовую площадь и, следовательно, сопротивление тела. Поскольку сопротивление также увеличивается с увеличением скорости движения, насекомое делает свой полет более эффективным, поскольку эта эффективность становится более необходимой. Кроме того, изменяя геометрический угол атаки при движении вниз, насекомое может сохранять полет с оптимальной эффективностью, выполняя максимально возможное количество маневров. Развитие общей тяги сравнительно невелико по сравнению с подъемной силой. Подъемная сила может более чем в три раза превышать вес насекомого, а тяга даже на самых высоких скоростях может составлять всего 20% от веса. Эта сила развивается главным образом за счет менее мощного подъема взмахов. [9] [16]
Хлопок и бросок, или механизм Вейса-Фога, открытый датским зоологом Торкелем Вейс-Фогом , представляет собой метод создания подъемной силы, используемый во время полета мелких насекомых. [17] Когда размеры насекомых становятся меньше 1 мм, силы вязкости становятся доминирующими, и эффективность подъемной силы аэродинамического профиля резко снижается. Начиная с положения хлопка, два крыла раздвигаются и вращаются вокруг задней кромки. Затем крылья разделяются и перемещаются горизонтально до конца хода вниз. Затем крылья прогибаются и используют переднюю кромку во время гребли вверх. Когда начинается движение хлопка, передние кромки встречаются и вращаются вместе, пока зазор не исчезнет. Первоначально считалось, что крылья соприкасаются, но несколько инцидентов указывают на зазор между крыльями и позволяют предположить, что это обеспечивает аэродинамическое преимущество. [18] [19]
Создание подъемной силы за счет механизма хлопка и броска происходит в ходе нескольких процессов на протяжении всего движения. Во-первых, механизм основан на взаимодействии крыльев, поскольку одно движение крыла не создает достаточной подъемной силы. [20] [21] [22] Когда крылья вращаются вокруг задней кромки при броске, воздух устремляется в образовавшийся зазор и создает сильный вихрь на передней кромке, а второй развивается на законцовках крыла. На задней кромке развивается третий, более слабый вихрь. [19] Сила развивающихся вихрей частично зависит от начального зазора между крыльями в начале броскового движения. Уменьшенный зазор между крыльями указывает на большую подъемную силу за счет большей силы сопротивления. Было обнаружено , что реализация подъемного движения во время броска, [23] гибких крыльев [21] и механизма замедленного срыва усиливает стабильность и крепление вихря. [24] Наконец, чтобы компенсировать общую меньшую подъемную силу во время полета с низким числом Рейнольдса (с ламинарным потоком ), крошечные насекомые часто имеют более высокую частоту взмахов, чтобы генерировать скорости кончиков крыльев, сравнимые с более крупными насекомыми. [24]
Наибольшие ожидаемые силы сопротивления возникают во время движения назад, поскольку крыльям необходимо отделиться и вращаться. [22] Ослабление больших сил сопротивления происходит за счет нескольких механизмов. Было обнаружено, что гибкие крылья уменьшают сопротивление при броске до 50% и еще больше снижают общее сопротивление на протяжении всего хода крыла по сравнению с жесткими крыльями. [21] Щетинки на краях крыла, как видно у Encarsia formosa , вызывают пористость потока, которая увеличивает и уменьшает силы сопротивления за счет снижения подъемной силы. [25] Кроме того, расстояние между крыльями перед броском играет важную роль в общем эффекте сопротивления. По мере увеличения расстояния между крыльями общее сопротивление уменьшается. [18]
Механизм хлопка и броска также используется морским моллюском Limacina helicina , морской бабочкой. [26] [27] Некоторые насекомые, такие как минер Liriomyza sativae (муха), используют частичный хлопок и бросок, используя механизм только на внешней части крыла, чтобы увеличить подъемную силу примерно на 7% при зависании. [24]






Крыло, движущееся в жидкости, испытывает действие силы жидкости , которая соответствует правилам аэродинамики. Составляющая силы, нормальная к направлению потока относительно крыла, называется подъемной силой ( L ), а составляющая силы, противоположная направлению потока, — сопротивлением ( D ). При рассматриваемых здесь числах Рейнольдса подходящей единицей силы является 1/2(ρU 2 S), где ρ — плотность жидкости, S — площадь крыла, а U — скорость крыла. Безразмерные силы называются коэффициентами подъемной силы ( CL ) и сопротивления ( CD ) , то есть: [8]
C L и C D являются постоянными, только если поток устойчив. Особый класс объектов, таких как аэродинамические профили , может достичь устойчивого состояния, когда он прорезает жидкость под небольшим углом атаки. В этом случае невязкое обтекание профиля можно аппроксимировать потенциальным течением, удовлетворяющим граничному условию непроникновения. Теорема Кутты-Жуковского о двумерном профиле также предполагает, что поток плавно покидает острую заднюю кромку, и это определяет общую циркуляцию вокруг профиля. Соответствующий подъем задается принципом Бернулли ( теорема Блазиуса ): [8]
Потоки вокруг птиц и насекомых можно считать несжимаемыми : число Маха , или скорость относительно скорости звука в воздухе, обычно составляет 1/300, а частота крыльев составляет около 10–103 Гц. Используя основное уравнение в виде уравнения Навье-Стокса, на которое распространяется граничное условие прилипания, уравнение имеет вид: [8]
Где u (x, t) — поле течения, p — давление, ρ — плотность жидкости, ν — кинематическая вязкость, u bd — скорость на границе, u — скорость твердого тела. Выбрав масштаб длины L и масштаб скорости U, уравнение можно выразить в безразмерной форме, содержащей число Рейнольдса R e =uL/ν. Есть два очевидных различия между крылом насекомого и аэродинамическим профилем: крыло насекомого намного меньше и машет крыльями. На примере стрекозы ее хорда (c) составляет около 1 см (0,39 дюйма), длина крыла (l) около 4 см (1,6 дюйма), а частота крыльев (f) около 40 Гц. Конечная скорость (u) составляет около 1 м/с (3,3 фута/с), а соответствующее число Рейнольдса около 103. На меньшем конце типичная хальцидоидная оса имеет длину крыла около 0,5–0,7 мм (0,020–0,028 мм). в) и бьет крылом с частотой около 400 Гц. Его число Рейнольдса около 25. Диапазон чисел Рейнольдса при полете насекомых составляет примерно от 10 до 10 4 , что лежит между двумя пределами, удобными для теорий: невязкое стационарное обтекание профиля и стоксово течение, испытываемое плавающей бактерией. По этой причине этот промежуточный диапазон недостаточно изучен. С другой стороны, это, пожалуй, самый вездесущий режим из тех, что мы видим. Падающие листья и семена, рыбы и птицы сталкиваются с неустойчивыми потоками, подобными тем, которые наблюдаются вокруг насекомого. [8] Хордовое число Рейнольдса можно описать следующим образом:
и
Где - средняя длина хорды, - скорость законцовки крыла, - амплитуда хода, - частота биений, - радиус вращения, - площадь крыла, - длина крыла, включая законцовку крыла.
Помимо числа Рейнольдса, существует как минимум два других важных безразмерных параметра. Крыло имеет три шкалы скорости: скорость взмаха относительно корпуса ( u ), скорость движения тела вперед ( U 0 ) и скорость тангажа ( Ω c ). Их отношения образуют две безразмерные переменные U 0 / u и Ωc/ u , первую часто называют коэффициентом опережения, и она также связана с приведенной частотой fc / U 0 . [8]
Если крыло насекомого жесткое, например, крыло дрозофилы примерно таковым, то его движение относительно неподвижного тела можно описать тремя переменными: положением кончика в сферических координатах , (Θ(t),Φ(t) ) и угол тангажа ψ(t) вокруг оси, соединяющей основание и вершину. Для оценки аэродинамических сил на основе анализа лопастных элементов необходимо также определить угол атаки (α). Типичный угол атаки при 70% размахе крыльев колеблется от 25° до 45° у парящих насекомых (15° у колибри). Несмотря на обилие данных, доступных для многих насекомых, относительно немногие эксперименты сообщают об изменении во времени α во время инсульта. Среди них — эксперименты в аэродинамической трубе с привязанной саранчой и привязанной мухой, а также свободный полет плодовой мухи. [8]
Поскольку их относительно легко измерить, о траекториях законцовок крыльев сообщают чаще. Например, если выбрать только те последовательности полета, которые обеспечивают достаточную подъемную силу, чтобы выдержать вес, вы увидите, что законцовка крыла имеет эллиптическую форму. Непересекающиеся формы наблюдались и у других насекомых. Независимо от их точной формы, закупорочное движение указывает на то, что насекомые могут использовать аэродинамическое сопротивление в дополнение к подъемной силе, чтобы поддерживать свой вес. [8]
.jpg/1280px-Hoverfly_September_2007-7_(cropped).jpg)
Многие насекомые могут парить или оставаться в одном месте в воздухе, быстро взмахивая крыльями. Для этого требуется боковая стабилизация, а также создание подъемной силы. Подъемная сила в основном создается за счет хода вниз. Когда крылья давят на окружающий воздух, возникающая в результате сила реакции воздуха на крыльях толкает насекомое вверх. Крылья большинства насекомых устроены так, что при взмахе вверх сила, действующая на крыло, невелика. Поскольку удары вниз и обратный ход заставляют насекомое подниматься и опускаться соответственно, насекомое колеблется и остается в том же положении. [14]
Расстояние, на которое насекомое пролетает между взмахами крыльев, зависит от того, насколько быстро бьются его крылья: чем медленнее оно машет, тем длиннее интервал падения и тем дальше оно падает между каждым взмахом крыльев. Можно вычислить частоту взмахов крыльев, необходимую насекомому для поддержания заданной стабильности их амплитуды. Для упрощения расчетов следует предположить, что подъемная сила имеет конечное постоянное значение при движении крыльев вниз и равна нулю при движении крыльев вверх. За промежуток времени Δt взмаха крыльев вверх насекомое под действием силы тяжести падает на расстояние h . [14]
Затем ход вверх возвращает насекомое в исходное положение. Обычно может потребоваться изменение вертикального положения насекомого не более чем на 0,1 мм (т. е. h = 0,1 мм). Тогда максимально допустимое время свободного падения составит [14]
Поскольку движения крыльев вверх и вниз примерно равны по длительности, период Т для полного подъема и опускания крыла равен удвоенному Δr , т. е. [14]
Частота взмахов f, означающая количество взмахов крыльев в секунду, представлена уравнением: [14]
В использованных примерах используемая частота составляет 110 ударов в секунду, что является типичной частотой, встречающейся у насекомых. У бабочек гораздо более медленная частота — около 10 ударов в секунду, а это значит, что они не могут парить. Другие насекомые могут производить частоту 1000 ударов в секунду. Чтобы вернуть насекомое в исходное вертикальное положение, средняя восходящая сила при движении вниз Fav должна быть равна удвоенному весу насекомого. Обратите внимание: поскольку направленная вверх сила к телу насекомого прикладывается только половину времени, средняя направленная вверх сила, действующая на насекомое, равна просто его весу. [14]
Теперь можно вычислить мощность, необходимую для поддержания зависания, снова рассматривая насекомое массой m 0,1 г: средняя сила Fav, приложенная двумя крыльями во время движения вниз, в два раза превышает вес. Поскольку давление, оказываемое крыльями, равномерно распределяется по всей площади крыла, это означает, что можно предположить, что сила, создаваемая каждым крылом, действует через одну точку в средней части крыльев. Во время хода вниз центр крыльев проходит вертикальное расстояние d . [14] Общая работа, совершаемая насекомым во время каждого удара вниз, является произведением силы и расстояния; то есть,
Если крылья раскачиваются в ходе взмаха под углом 70°, то в случае, представленном для насекомого с длиной крыльев 1 см, d равно 0,57 см. Следовательно, работа, совершаемая за каждый взмах двумя крыльями, равна: [14]
Энергия используется, чтобы поднять насекомое против силы тяжести. Энергия E , необходимая для подъема массы насекомого на 0,1 мм при каждом движении вниз, равна: [14]
Это незначительная часть общей затраченной энергии, и очевидно, что большая часть энергии расходуется на другие процессы. Более детальный анализ задачи показывает, что работа, совершаемая крыльями, преобразуется преимущественно в кинетическую энергию воздуха, ускоряемого движением крыльев вниз. Мощность — это количество работы, совершаемой за 1 с; у насекомого, взятого в качестве примера, совершает 110 движений вниз в секунду. Следовательно, его выходная мощность P равна ударам в секунду, а это означает, что его выходная мощность P равна: [14]
При расчете мощности, используемой при зависании, в использованных примерах не учитывалась кинетическая энергия движущихся крыльев. Крылья насекомых, какими бы легкими они ни были, имеют конечную массу; следовательно, при движении они обладают кинетической энергией. Поскольку крылья находятся во вращательном движении, максимальная кинетическая энергия во время каждого взмаха крыла равна: [14]
Здесь I — момент инерции крыла, а ω max — максимальная угловая скорость при ходе крыла. Чтобы получить момент инерции крыла, предположим, что крыло можно аппроксимировать тонким стержнем, повернутым на одном конце. Тогда момент инерции крыла составит: [14]
Где l — длина крыла (1 см), а m — масса двух крыльев, которая обычно может составлять 10–3 г. Максимальная угловая скорость ω max может быть рассчитана по максимальной линейной скорости ν max в центре крыла: [14]
Во время каждого взмаха центр крыльев движется со средней линейной скоростью ν av , определяемой расстоянием d, пройденным центром крыла, деленным на продолжительность Δt хода крыла. В нашем предыдущем примере d = 0,57 см и Δt = 4,5×10 -3 с. Поэтому: [14]
Скорость крыльев равна нулю как в начале, так и в конце хода крыла, что означает, что максимальная линейная скорость выше средней скорости. Если предположить, что скорость колеблется ( синусоидально ) вдоль траектории крыла, то максимальная скорость в два раза превышает среднюю скорость. Следовательно, максимальная угловая скорость равна: [14]
И поэтому кинетическая энергия равна: [14]
Поскольку в каждом цикле движения крыла происходит два взмаха крыла (ход вверх и вниз), кинетическая энергия равна 2×43 = 86 эрг . Это примерно столько же энергии, сколько расходуется на само зависание. [14]

Насекомые получают кинетическую энергию, обеспечиваемую мышцами, когда крылья ускоряются . Когда крылья начинают замедляться к концу взмаха, эта энергия должна рассеиваться. Во время удара вниз кинетическая энергия рассеивается самими мышцами и преобразуется в тепло (иногда это тепло используется для поддержания внутренней температуры тела). Некоторые насекомые способны использовать кинетическую энергию движения крыльев вверх для облегчения полета. Суставы крыльев этих насекомых содержат подушечки из эластичного резиноподобного белка, называемого резилином . При подъеме крыла вверх резилин растягивается. Кинетическая энергия крыла преобразуется в потенциальную энергию в растянутой резилине, который сохраняет энергию подобно пружине. Когда крыло движется вниз, эта энергия высвобождается и способствует движению вниз. [14]
Используя несколько упрощающих предположений, мы можем вычислить количество энергии, запасенной в растянутом резилине. Хотя резилин согнут в сложную форму, в приведенном примере показан расчет как прямой стержень площадью А и длиной. Кроме того, будем считать, что на всем протяжении растяжения резилин подчиняется закону Гука . Это не совсем так, поскольку резилин растягивается на значительную величину, и поэтому в процессе растяжения изменяются как площадь, так и модуль Юнга . Потенциальная энергия U, запасенная в растянутом резилине, равна: [14]
Здесь E представляет собой модуль Юнга для резилина, который, согласно измерениям, составляет 1,8×10 7 дин /см 2 . Обычно у насекомого размером с пчелу объем резилина может быть эквивалентен цилиндру длиной 2×10 -2 см и площадью 4×10 -4 см 2 . В приведенном примере длина резилинового стержня при растяжении увеличивается на 50%. То есть Δℓ составляет 10 −2 см. Следовательно, в этом случае потенциальная энергия, запасенная в резилине каждого крыла, равна: [14]
Запасенная энергия в двух крыльях насекомого размером с пчелу составляет 36 эрг, что сравнимо с кинетической энергией при взмахе крыльев вверх. Эксперименты показывают, что до 80% кинетической энергии крыла может храниться в резилине. [14]

Некоторые отряды четырехкрылых насекомых, такие как Lepidoptera, развили у имаго морфологические механизмы соединения крыльев , которые делают эти таксоны функционально двукрылыми. [30] Все формы, кроме самых базальных, демонстрируют это соединение крыльев. [31]
Механизмы бывают трех разных типов: скуловой, уздечно-ретинакулярный и плексиформный: [32]
Биохимия полета насекомых была в центре внимания серьезных исследований. В то время как многие насекомые используют углеводы и липиды в качестве источника энергии для полета, многие жуки и мухи используют в качестве источника энергии аминокислоту пролин . [35] Некоторые виды также используют комбинацию источников, а мотыльки, такие как Manduca sexta, используют углеводы для предполетной разминки. [36]
Насекомые используют сенсорную обратную связь для поддержания и контроля полета. Исследования продемонстрировали роль сенсорных структур, таких как усики , [37] жужжальца [38] и крылья [39] в контроле позы полета, амплитуды и частоты взмахов крыльев.
Где-то в каменноугольном периоде , примерно 350–300 миллионов лет назад, когда существовало всего два крупных массива суши, насекомые начали летать. [40] Среди старейших окаменелостей крылатых насекомых — Delitzschala , палеодиктиоптеран из нижнего карбона; [41] Rhyniognatha старше, из раннего девона , но неясно, были ли у нее крылья или действительно было насекомое. [42] [43]
Как и почему развились крылья насекомых, до конца не понятно, во многом из-за нехватки соответствующих окаменелостей периода их развития в нижнем карбоне. Исторически существовало три основные теории происхождения полета насекомых. Во-первых, они представляют собой модификации подвижных брюшных жабр , обнаруженных у водных наяд подёнок . Филогеномный анализ показывает, что Polyneoptera , группа крылатых насекомых, в которую входят кузнечики, произошли от наземного предка, что делает маловероятным появление крыльев из жабр. [44] Дополнительное исследование прыжкового поведения личинок поденок показало, что трахейные жабры не играют никакой роли в управлении спуском насекомых, что дает дополнительные доказательства против этой эволюционной гипотезы. [45] Это оставляет две основные исторические теории: крылья развились из паранотальных долей, расширений грудной терги ; или что они возникли в результате модификаций сегментов ног, которые уже содержали мышцы. [46]
.JPG/1280px-Mayfly_nymph_dorsal_view_wing_buds_paired_gills_(rotated).JPG)
Многочисленные [47] энтомологи , в том числе Ландуа в 1871 году, Лаббок в 1873 году, Грабер в 1877 году и Осборн в 1905 году, предположили, что возможным источником крыльев насекомых могли быть подвижные брюшные жабры, обнаруженные у многих водных насекомых, например, у наяд подёнок. Согласно этой теории, жабры трахеи, которые начинали как выходы дыхательной системы и со временем превратились в двигательные функции, в конечном итоге превратились в крылья. Жабры трахеи оснащены маленькими крылышками, которые постоянно вибрируют, и имеют собственные крошечные прямые мышцы. [48]
Гипотеза паранотальной доли или тергала (дорсальной стенки тела), предложенная Фрицем Мюллером в 1875 году [49] и переработанная Г. Крэмптоном в 1916 году, [47] Ярмилой Кукаловой-Пек в 1978 году [50] и Александром П. Расницыным в 1981 году среди другие [51] предполагают, что крылья насекомого развились из паранотальных долей - преадаптации , обнаруженной в окаменелостях насекомых , которая способствовала стабилизации при прыжке или падении. В пользу этой гипотезы говорит склонность большинства насекомых, испуганных при лазании по ветвям, спасаться бегством, падая на землю. Такие доли служили бы парашютами и позволяли насекомому мягче приземляться. Теория предполагает, что эти доли постепенно увеличивались и на более поздней стадии образовали соединение с грудной клеткой. Еще позже появились мышцы, способные двигать этими грубыми крыльями. Эта модель подразумевает постепенное повышение эффективности крыльев, начиная с парашютного прыжка , затем планирования и, наконец, активного полета . Тем не менее, отсутствие существенных ископаемых свидетельств развития суставов и мышц крыльев представляет собой серьезную трудность для теории, равно как и кажущееся спонтанным развитие сочленений и жилкования, и это в значительной степени отвергается экспертами в этой области. [48]

В 1990 году Дж. У. Труман предположил, что крыло было адаптировано из эндитов и выходов , придатков на соответствующих внутренних и внешних сторонах конечностей примитивных членистоногих , что также называется плевральной гипотезой. Это было основано на исследовании Гольдшмидта в 1945 году на Drosophila melanogaster , в котором вариация под названием «стручок» (подомеры — сегменты конечностей) продемонстрировала мутацию, которая трансформировала нормальные крылья. Результат был интерпретирован как трехчленистая конструкция ног с некоторыми дополнительными придатками, но без предплюсны, на месте которой обычно находится реберная поверхность крыла. Эта мутация была переинтерпретирована как убедительное доказательство слияния дорсального выхода и эндита, а не ноги, при этом придатки гораздо лучше соответствуют этой гипотезе. Иннервация, артикуляция и мускулатура, необходимые для развития крыльев, уже присутствуют в сегментах конечностей. [46]
Другие гипотезы включают в себя предположение Винсента Вигглсворта в 1973 году о том, что крылья развились из грудных выступов, используемых в качестве радиаторов . [48]
Адриан Томас и Оке Норберг предположили в 2003 году, что крылья, возможно, изначально развились для плавания по поверхности воды, как это наблюдается у некоторых веснянок . [52] [53]
Стивен П. Яновяк и его коллеги предположили в 2009 году, что крыло возникло в результате направленного воздушного планирующего спуска — предполетного явления, обнаруженного у некоторых аптеригот , бескрылого родственного таксона крылатых насекомых. [54]
Биологи, в том числе Авероф, [55] Нива, [56] Элиас-Нето [57] и их коллеги, начали исследовать происхождение крыльев насекомых, используя эво-дево в дополнение к палеонтологическим данным. Это предполагает, что крылья серийно гомологичны как тергальным, так и плевральным структурам, что потенциально разрешает многовековые споры. [58] Якуб Прокоп и его коллеги в 2017 году обнаружили палеонтологические доказательства на палеозойских нимфальных подушечках крыльев того, что крылья действительно имели такое двойное происхождение. [59]
{{cite book}}: |work=игнорируется ( помощь )





























