
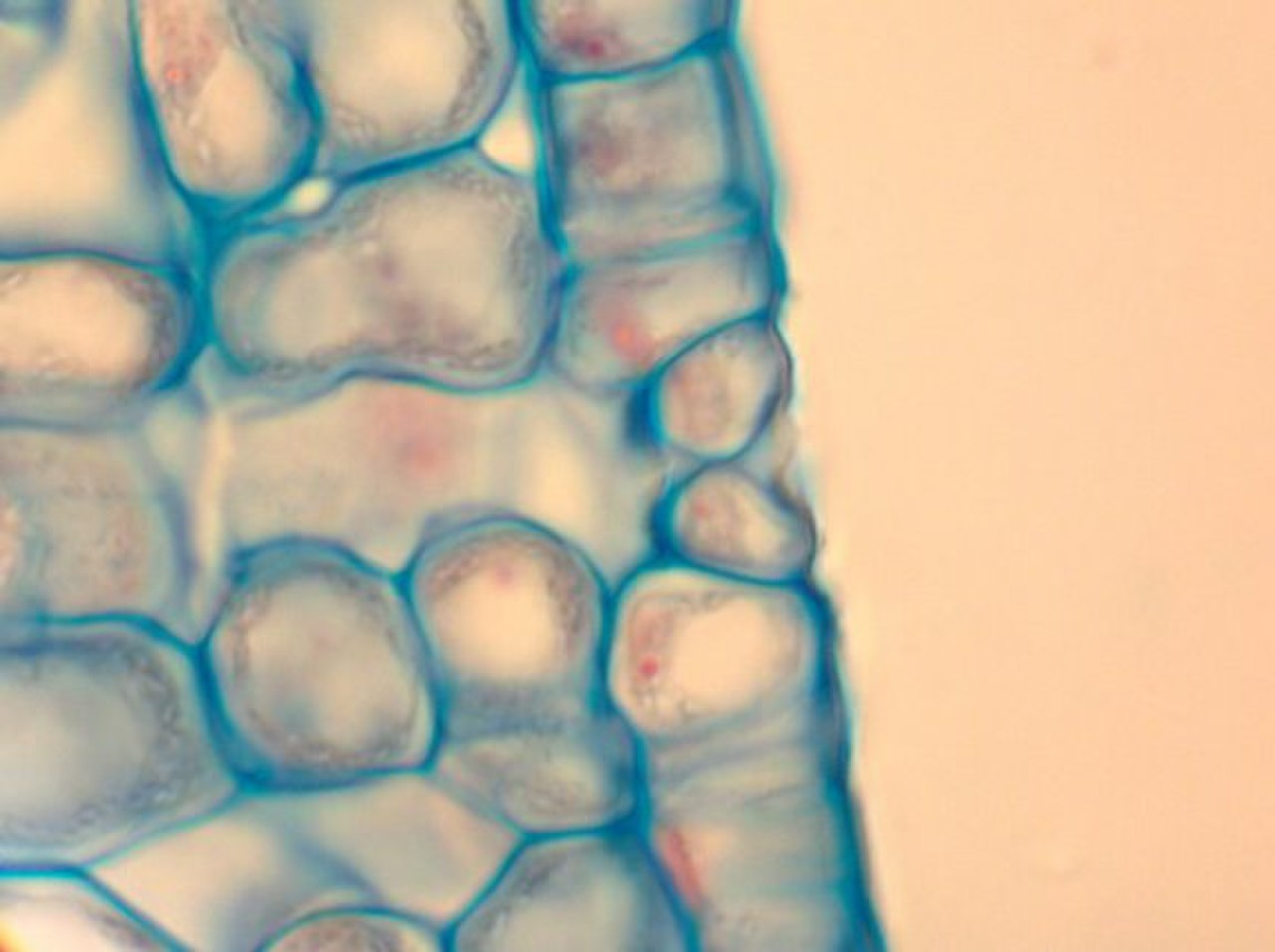

В ботанике устьица ( мн.: устьица , от греческого στόμα , «рот»), также называемая устьицей ( мн .: устьица ) , представляет собой пору, обнаруженную в эпидермисе листьев, стеблей и других органов, которая контролирует скорость газообмена между внутренними воздушными пространствами листа и атмосферой. Пора ограничена парой специализированных клеток паренхимы , известных как замыкающие клетки , которые регулируют размер устьичного отверстия.
Этот термин обычно используется в совокупности для обозначения всего устьичного комплекса, состоящего из парных замыкающих клеток и самой поры, называемой устьичным отверстием. [1] Воздух, содержащий кислород , который используется при дыхании , и углекислый газ , который используется при фотосинтезе , проходит через устьица путем газовой диффузии . Водяной пар диффундирует через устьица в атмосферу как часть процесса, называемого транспирацией .
Устьица присутствуют в спорофитном поколении подавляющего большинства наземных растений , за исключением печеночников , а также некоторых мхов и роголистников . У сосудистых растений количество, размер и расположение устьиц широко варьируют. Двудольные обычно имеют больше устьиц на нижней поверхности листьев, чем на верхней. Однодольные растения , такие как лук , овес и кукуруза , могут иметь примерно одинаковое количество устьиц на обеих поверхностях листьев. [2] : 5 У растений с плавающими листьями устьица могут быть обнаружены только на верхнем эпидермисе, а у погруженных листьев устьица могут полностью отсутствовать. У большинства видов деревьев устьица есть только на нижней поверхности листа. [3] Листья с устьицами как на верхней, так и на нижней поверхности листа называются амфистоматозными листьями; листья с устьицами только на нижней поверхности — гипостоматозные , а листья с устьицами только на верхней поверхности — эпистоматозные или гиперстоматозные . [3] Размер варьируется в зависимости от вида: длина от конца до конца варьируется от 10 до 80 мкм , а ширина - от нескольких до 50 мкм. [4]

Углекислый газ , ключевой реагент фотосинтеза , присутствует в атмосфере в концентрации около 400 частей на миллион. Большинству растений требуется, чтобы устьица были открыты в дневное время. Воздушные пространства листа насыщены водяным паром , который выходит из листа через устьица в процессе транспирации . Следовательно, растения не могут получать углекислый газ, не теряя при этом водяной пар. [5]
Обычно углекислый газ связывается с рибулозо-1,5-бисфосфатом (RuBP) с помощью фермента RuBisCO в клетках мезофилла , находящихся непосредственно в воздушном пространстве внутри листа. Это усугубляет проблему транспирации по двум причинам: во-первых, RuBisCo имеет относительно низкое сродство к углекислому газу, а во-вторых, он связывает кислород с RuBP, тратя энергию и углерод впустую в процессе, называемом фотодыханием . По обеим этим причинам Рубиско необходима высокая концентрация углекислого газа, что означает широкие устьичные отверстия и, как следствие, большие потери воды.
Более узкие устьичные отверстия можно использовать в сочетании с промежуточной молекулой с высоким сродством к диоксиду углерода, фосфоенолпируваткарбоксилазой (PEPcase). Однако извлечение продуктов фиксации углерода из PEPCase — энергозатратный процесс. В результате альтернатива PEPCase предпочтительна только там, где количество воды ограничено, но много света, или где высокие температуры увеличивают растворимость кислорода по сравнению с растворимостью углекислого газа, усугубляя проблему оксигенации RuBisCo.
.svg/1280px-Differences_in_Stomata_Opening_Throughout_the_Day_for_C3_plants_and_CAM_plants_(1).svg.png)
Группа преимущественно пустынных растений, называемых растениями «САМ» ( кислотный метаболизм толстянковых , по названию семейства Crassulaceae, включающего виды, у которых впервые был обнаружен процесс САМ), открывает свои устьица ночью (когда вода испаряется из листьев медленнее в течение заданного периода времени). степень открытия устьиц), используют ПЭПкейс для фиксации углекислого газа и хранения продуктов в крупных вакуолях. На следующий день они закрывают устьица и выделяют углекислый газ, зафиксированный накануне вечером, в присутствии РуБисКО. Это насыщает RuBisCO углекислым газом, обеспечивая минимальное фотодыхание. Однако этот подход сильно ограничен способностью хранить фиксированный углерод в вакуолях, поэтому он предпочтителен только тогда, когда вода сильно ограничена.

Однако у большинства растений нет САМ, и поэтому они должны открывать и закрывать устьица в дневное время в ответ на изменяющиеся условия, такие как интенсивность света, влажность и концентрация углекислого газа. Когда условия способствуют открытию устьиц (например, высокая интенсивность света и высокая влажность), протонный насос вытесняет протоны (H + ) из замыкающих клеток. Это означает, что электрический потенциал клеток становится все более отрицательным. Отрицательный потенциал открывает калиевые потенциалзависимые каналы, и таким образом происходит поглощение ионов калия (K + ). Чтобы поддерживать это внутреннее отрицательное напряжение и не прекращать поступление ионов калия, отрицательные ионы уравновешивают приток калия. В одних случаях поступают ионы хлорида, а у других растений в замыкающих клетках образуется малат органического иона . Это увеличение концентрации растворенных веществ снижает водный потенциал внутри клетки, что приводит к диффузии воды в клетку посредством осмоса . Это увеличивает объем клетки и тургорное давление . Затем, благодаря кольцам целлюлозных микрофибрилл , которые предотвращают набухание замыкающих клеток по ширине и, таким образом, позволяют дополнительному тургорному давлению удлинять замыкающие клетки, концы которых прочно удерживаются на месте окружающими эпидермальными клетками, две замыкающие клетки удлиняются. отклоняясь друг от друга, создавая открытую пору, через которую может диффундировать газ. [6]
Когда корни начинают ощущать нехватку воды в почве, выделяется абсцизовая кислота (АБК). [7] АБК связывается с рецепторными белками в плазматической мембране замыкающих клеток и цитозоле, что сначала повышает pH цитозоля клеток и вызывает увеличение концентрации свободного Са 2+ в цитозоле за счет притока извне клетки. и высвобождение Ca 2+ из внутренних хранилищ, таких как эндоплазматический ретикулум и вакуоли. [8] Это приводит к выходу хлоридов (Cl - ) и органических ионов из клеток. Во-вторых, это останавливает дальнейшее поступление K + в клетки и, как следствие, потерю K + . Потеря этих растворенных веществ вызывает увеличение водного потенциала , что приводит к диффузии воды обратно из клетки путем осмоса . Это приводит к плазмолизу клетки , что приводит к закрытию устьичных пор.
Замыкающие клетки содержат больше хлоропластов, чем другие эпидермальные клетки, из которых произошли замыкающие клетки. Их функция спорна. [9] [10]
Степень устьичной резистентности можно определить путем измерения газообмена листа. Скорость транспирации зависит от диффузионного сопротивления, обеспечиваемого устьичными порами, а также от градиента влажности между внутренними воздушными пространствами листа и наружным воздухом. Таким образом , устьичное сопротивление (или его обратная устьичная проводимость ) можно рассчитать по скорости транспирации и градиенту влажности. Это позволяет ученым исследовать, как устьица реагируют на изменения условий окружающей среды, таких как интенсивность света и концентрации таких газов, как водяной пар, углекислый газ и озон . [11] Испарение ( E ) можно рассчитать как [12]
где e i и e a — парциальное давление воды в листе и окружающем воздухе соответственно, P — атмосферное давление, r — устьичное сопротивление. Обратная величина r — это проводимость водяного пара ( g ), поэтому уравнение можно переписать в [12]
и решено для g : [12]
Фотосинтетическая ассимиляция CO 2 ( A ) может быть рассчитана по формуле
где Ca и C i — атмосферное и субустьичное парциальное давление CO 2 соответственно [ необходимы пояснения ] . Скорость испарения с листа можно определить с помощью системы фотосинтеза . Эти научные инструменты измеряют количество водяного пара, выходящего из листа, и давление пара окружающего воздуха. Фотосинтетические системы могут рассчитывать эффективность использования воды ( A / E ), g , собственную эффективность использования воды ( A / g ) и C i . Эти научные инструменты обычно используются физиологами растений для измерения поглощения CO 2 и, таким образом, измерения скорости фотосинтеза. [13] [14]
В летописи окаменелостей мало свидетельств эволюции устьиц, но у наземных растений они появились к середине силурийского периода. [15] Возможно, они произошли в результате модификации концепций водорослеподобных предков растений. [16] Однако эволюция устьиц, должно быть, произошла одновременно с развитием восковой кутикулы – эти две черты вместе составляли главное преимущество для ранних наземных растений. [ нужна цитата ]
Существует три основных типа эпидермальных клеток, которые в конечном итоге происходят из самого внешнего (L1) слоя ткани апикальной меристемы побега , называемого протодермальными клетками: трихомы , клетки мостовой и замыкающие клетки, все из которых расположены неслучайным образом.
В протодермальных клетках происходит асимметричное деление клеток, в результате чего образуется одна большая клетка, которой суждено стать опорной клеткой, и меньшая клетка, называемая меристемоидом, которая в конечном итоге дифференцируется в замыкающие клетки, окружающие стому. Затем этот меристемоид асимметрично делится один-три раза, прежде чем дифференцироваться в замыкающую материнскую клетку. Затем замыкающая материнская клетка делает одно симметричное деление, образующее пару замыкающих клеток. [17] В некоторых клетках деление ингибируется, поэтому между устьицами всегда есть хотя бы одна клетка. [18]
Формирование устьичного паттерна контролируется взаимодействием многих компонентов сигнальной трансдукции , таких как EPF (фактор эпидермального паттерна), ERL (ERecta Like) и YODA (предполагаемая киназа киназы MAP-киназы ). [18] Мутации любого из генов, кодирующих эти факторы, могут изменить развитие устьиц в эпидермисе. [18] Например, мутация в одном гене приводит к тому, что устьица сгруппированы вместе, поэтому это называется «слишком много ртов» ( TMM ). [17] Принимая во внимание, что нарушение гена SPCH (SPeecCHless) в целом предотвращает развитие устьиц. [18] Подавление устьичной продукции может происходить за счет активации EPF1, который активирует TMM/ERL, которые вместе активируют YODA. YODA ингибирует SPCH, вызывая снижение активности SPCH, предотвращая асимметричное деление клеток, которое инициирует образование устьиц. [18] [19] Развитие устьиц также координируется клеточным пептидным сигналом, называемым стомагеном, который сигнализирует об активации SPCH, что приводит к увеличению количества устьиц. [20]
Экологические и гормональные факторы могут влиять на развитие устьиц. Свет ускоряет развитие устьиц растений; в то время как растения, выращенные в темноте, имеют меньшее количество устьиц. Ауксин подавляет развитие устьиц, влияя на их развитие на уровне рецепторов, таких как рецепторы ERL и TMM. Однако низкая концентрация ауксина обеспечивает равное деление замыкающей материнской клетки и увеличивает вероятность образования замыкающих клеток. [21]
У большинства покрытосеменных деревьев устьица есть только на нижней поверхности листьев. У тополей и ив они есть на обеих поверхностях. Когда на обеих поверхностях листьев появляются устьица, устьица на нижней поверхности, как правило, крупнее и многочисленнее, но могут существовать большие различия в размере и частоте появления в зависимости от вида и генотипа. Листья белого ясеня и белой березы имели меньше устьиц, но были крупнее по размеру. С другой стороны, клен сахарный и клен серебристый имели более мелкие устьица. [22]
Существуют различные классификации типов стом. Один из них, получивший широкое распространение, основан на типах, которые Жюльен Жозеф Веск представил в 1889 году, получил дальнейшее развитие Меткалфа и Чока [23] и позднее был дополнен другими авторами. Он основан на размере, форме и расположении вспомогательных клеток, окружающих две замыкающие клетки. [24] У двудольных различают :
У однодольных растений встречается несколько различных типов устьиц, таких как:
У папоротников выделяют четыре разных типа:
Устьичные крипты — это вдавленные участки эпидермиса листа, которые образуют камерообразную структуру, содержащую одно или несколько устьиц, а иногда и трихомы или скопления воска . Устьичные крипты могут быть адаптацией к засухе и засушливым климатическим условиям, когда устьичные крипты очень выражены. Однако сухой климат — не единственные места, где их можно встретить. Следующие растения являются примерами видов с устьичными криптами или предкамерами: Nerium oleander , хвойные деревья, Hakea [26] и Drimys Winteri , вид растений, встречающихся в облачном лесу . [27]
Устьица — это очевидные отверстия в листе, через которые, как предполагалось некоторое время, болезнетворные микроорганизмы могут беспрепятственно проникать в лист. Однако недавно было показано, что устьица действительно ощущают присутствие некоторых, если не всех, патогенов. Однако при нанесении вирулентных бактерий на листья растения Arabidopsis в ходе эксперимента бактерии выделили химический коронатин , который заставил устьица снова открыться в течение нескольких часов. [28]
Фотосинтез , транспорт воды в растениях ( ксилема ) и газообмен регулируются функцией устьиц, которая важна для функционирования растений. [29]
Устьица реагируют на свет: синий свет почти в 10 раз эффективнее красного света вызывает устьичную реакцию. Исследования показывают, что это связано с тем, что световая реакция устьиц на синий свет не зависит от других компонентов листьев, таких как хлорофилл . Протопласты замыкающих клеток набухают под синим светом при наличии достаточного количества калия . [30] Многочисленные исследования подтвердили, что увеличение концентрации калия может увеличить открытие устьиц по утрам, до начала процесса фотосинтеза, но позже в течение дня сахароза играет большую роль в регулировании открытия устьиц. [31] Зеаксантин в замыкающих клетках действует как фоторецептор синего света, который опосредует открытие устьиц. [32] Эффект синего света на замыкающие клетки нейтрализуется зеленым светом, который изомеризует зеаксантин. [32]
Плотность и апертура устьиц (длина устьиц) варьируются в зависимости от ряда факторов окружающей среды, таких как концентрация CO 2 в атмосфере , интенсивность света, температура воздуха и фотопериод (продолжительность светового дня).[33] [34]
Снижение плотности устьиц – один из способов реакции растений на увеличение концентрации CO 2 в атмосфере ([CO 2 ] атм ). [35] Хотя изменения в реакции [CO 2 ] на атмосферу наименее понятны с механистической точки зрения, эта устьичная реакция начала выходить на плато, и вскоре ожидается, что она повлияет на процессы транспирации и фотосинтеза в растениях. [29] [36]
Засуха препятствует открытию устьиц, но исследования соевых бобов показывают, что умеренная засуха не оказывает существенного влияния на закрытие устьиц листьев. Существуют разные механизмы закрытия устьиц. Низкая влажность вызывает стресс замыкающих клеток, вызывая потерю тургора , называемую гидропассивным закрытием. Гидроактивное закрытие контрастирует с тем, что весь лист подвергается стрессу, вызванному засухой, который, скорее всего, вызван абсцизовой кислотой . [37]
Ожидается, что [CO 2 ] атм достигнет 500–1000 ppm к 2100 году. [29] 96% из последних 400 000 лет испытывали концентрацию CO 2 ниже 280 ppm . Судя по этой цифре, весьма вероятно, что генотипы современных растений разошлись с их доиндустриальными родственниками. [29]
Ген HIC (высокий уровень углекислого газа) кодирует негативный регулятор развития устьиц у растений. [38] Исследование гена HIC с использованием Arabidopsis thaliana не выявило увеличения развития устьиц в доминантном аллеле , но в рецессивном аллеле «дикого типа» было обнаружено значительное увеличение, как в ответ на повышение уровня CO 2 в атмосфере. [38] Эти исследования предполагают, что реакция растений на изменение уровня CO 2 в значительной степени контролируется генетикой.
Влияние удобрений CO 2 было сильно переоценено в ходе экспериментов по обогащению углекислого газа в свободном воздухе (FACE), результаты которых показали, что повышение уровня CO 2 в атмосфере усиливает фотосинтез, снижает транспирацию и повышает эффективность использования воды (WUE). [35] Увеличение биомассы является одним из эффектов моделирования экспериментов, предсказывающих увеличение урожайности сельскохозяйственных культур на 5–20% при 550 ppm CO 2 . [39] Было показано, что скорость фотосинтеза листьев увеличивается на 30–50% у растений C3 и на 10–25% у растений C4 при удвоении уровня CO 2 . [39] Существование механизма обратной связи приводит к фенотипической пластичности в ответ на [CO 2 ] атм , которая могла быть адаптивной чертой в эволюции дыхания и функций растений. [29] [34]
Прогнозирование того, как устьица будут вести себя во время адаптации, полезно для понимания продуктивности растительных систем как в природных, так и в сельскохозяйственных системах . [33] Селекционеры и фермеры начинают работать вместе, используя эволюционную и совместную селекцию растений, чтобы найти наиболее подходящие виды, такие как сорта сельскохозяйственных культур, устойчивые к жаре и засухе, которые могут естественным образом адаптироваться к изменениям перед лицом проблем продовольственной безопасности. [35]
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )


