Сукцинатдегидрогеназа ( SDH ) или сукцинат-кофермент Q-редуктаза ( SQR ) или дыхательный комплекс II — это ферментный комплекс, обнаруженный во многих бактериальных клетках и во внутренней митохондриальной мембране эукариот . Это единственный фермент, который участвует как в цикле лимонной кислоты, так и в окислительном фосфорилировании . [1] Гистохимический анализ , показывающий высокий уровень сукцинатдегидрогеназы в мышцах, демонстрирует высокое содержание митохондрий и высокий окислительный потенциал. [2]
На этапе 6 цикла лимонной кислоты SQR катализирует окисление сукцината до фумарата с восстановлением убихинона до убихинола . Это происходит во внутренней митохондриальной мембране путем объединения двух реакций.
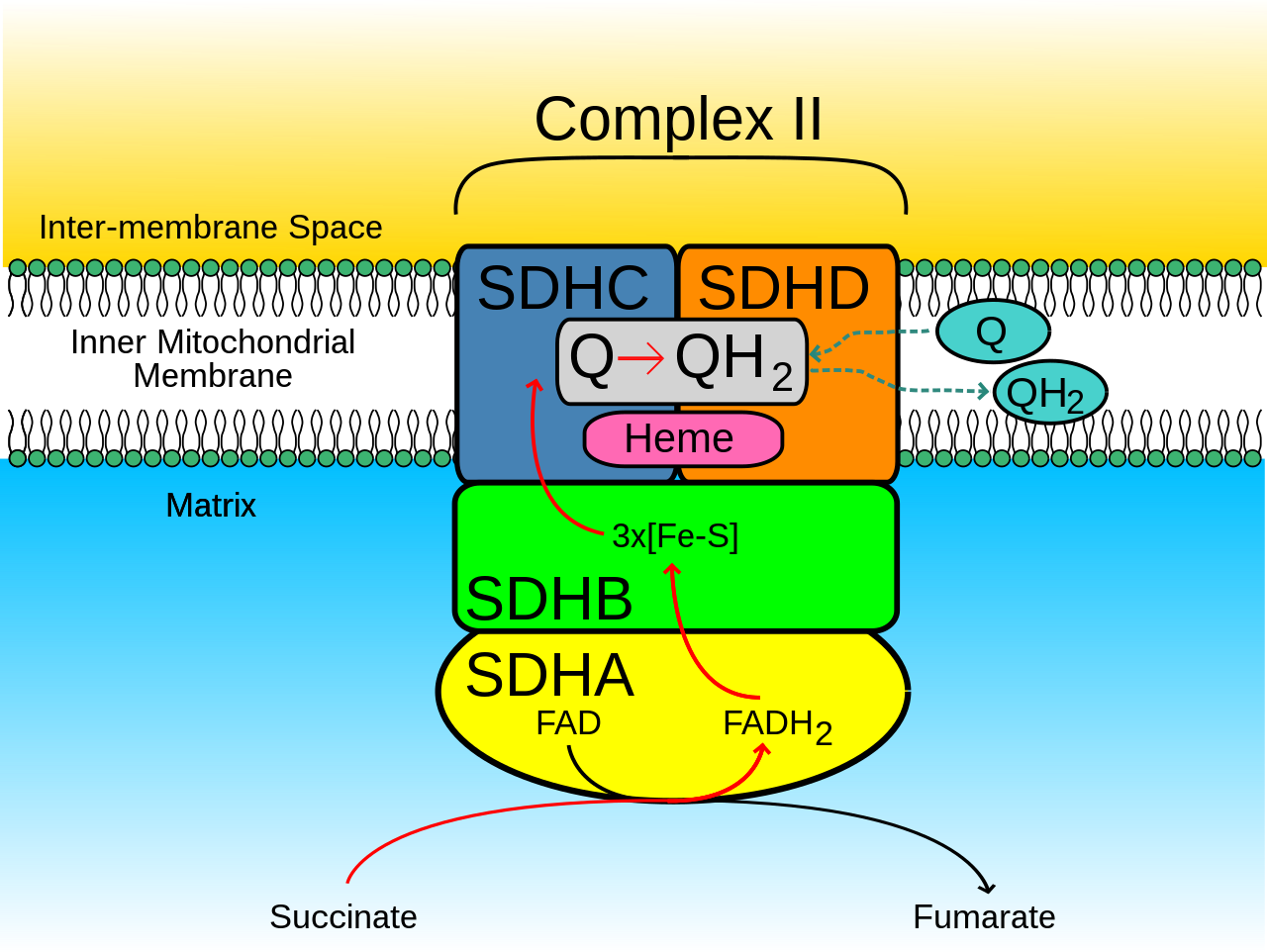
Митохондриальные и многие бактериальные SQR состоят из четырех структурно различных субъединиц : двух гидрофильных и двух гидрофобных . Первые две субъединицы, флавопротеин (SDHA) и железо-серный белок (SDHB), образуют гидрофильную головку, где происходит ферментативная активность комплекса. SDHA содержит ковалентно связанный кофактор флавинадениндинуклеотида (FAD) и сайт связывания сукцината , а SDHB содержит три железо-серных кластера: [2Fe-2S], [4Fe-4S] и [3Fe-4S]. Вторые две субъединицы являются гидрофобными субъединицами мембранного якоря, SDHC и SDHD. Человеческие митохондрии содержат две различные изоформы SDHA (субъединицы Fp типа I и типа II), эти изоформы также встречаются у Ascaris suum и Caenorhabditis elegans . [3] Субъединицы образуют связанный с мембраной комплекс цитохрома b с шестью трансмембранными спиралями , содержащими одну группу гема b и сайт связывания убихинона . Две молекулы фосфолипида , одна кардиолипиновая и одна фосфатидилэтаноламиновая , также обнаружены в субъединицах SDHC и SDHD (не показаны на изображении). Они служат для занятия гидрофобного пространства под гемом b. Эти субъединицы показаны на приложенном изображении. SDHA имеет зеленый цвет, SDHB имеет цвет сине-зеленой зелени, SDHC имеет цвет фуксии, а SDHD имеет желтый цвет. Вокруг SDHC и SDHD находится фосфолипидная мембрана с межмембранным пространством в верхней части изображения. [4]
На SDH млекопитающих можно распознать два отличительных сайта связывания убихинона – матрикс-проксимальный Q P и матрикс-дистальный Q D . Сайт связывания убихинона Qp, который показывает более высокое сродство к убихинону, расположен в зазоре, состоящем из SDHB, SDHC и SDHD. Убихинон стабилизирован боковыми цепями His207 субъединицы B, Ser27 и Arg31 субъединицы C и Tyr83 субъединицы D. Хинонное кольцо окружено Ile28 субъединицы C и Pro160 субъединицы B. Эти остатки вместе с Il209, Trp163 и Trp164 субъединицы B и Ser27 (атом C) субъединицы C образуют гидрофобную среду кармана связывания хинона Qp. [6] Напротив, участок связывания убихинона QD , который находится ближе к межмембранному пространству, состоит только из SDHD и имеет более низкое сродство к убихинону. [7]
SDHA обеспечивает сайт связывания для окисления сукцината . Боковые цепи Thr254, His354 и Arg399 субъединицы A стабилизируют молекулу , в то время как FAD окисляется и переносит электроны к первому из железо-серных кластеров , [2Fe-2S]. [8] Это можно увидеть на изображении 5.
Связывающий участок сукцината и связывающий участок убихинона соединены цепочкой окислительно-восстановительных центров, включающей FAD и кластеры железа и серы . Эта цепочка простирается более чем на 40 Å через мономер фермента . Все расстояния от края до края между центрами меньше предполагаемого предела в 14 Å для физиологического переноса электронов . [4] Этот перенос электронов показан на рисунке 8.
Все субъединицы митохондриальной SDH человека кодируются в ядре. После трансляции субъединица SDHA транслоцируется как апопротеин в митохондриальный матрикс. Впоследствии одним из первых шагов является ковалентное присоединение кофактора FAD ( ковалентное флавинилирование). Этот процесс усиливается фактором сборки сукцинатдегидрогеназы 2 ( SDHAF2 ; [9] также называемым SDH5 у дрожжей и SDHE у бактерий) и некоторыми промежуточными продуктами цикла Кребса. Фумарат сильнее всего стимулирует ковалентное флавинилирование SDHA. [10] Благодаря исследованиям бактериальной системы было показано, что механизм присоединения FAD включает промежуточный хинон:метид. [11] В митохондриальной, но не бактериальной, сборке SDHA взаимодействует со вторым фактором сборки, называемым фактором сборки сукцинатдегидрогеназы 4 (SDHAF4; называемым SDH8 у дрожжей), прежде чем он будет вставлен в конечный комплекс. [7]
Простетические группы Fe-S субъединицы SDHB предварительно формируются в митохондриальной матрице белковым комплексом ISU. Также считается, что комплекс способен вставлять кластеры железа и серы в SDHB во время его созревания. Исследования показывают, что вставка кластера Fe-S предшествует образованию димера SDHA-SDHB. Такое включение требует восстановления остатков цистеина в активном центре SDHB. Как восстановленные остатки цистеина, так и уже включенные кластеры Fe-S весьма восприимчивы к повреждению ROS . Еще два фактора сборки SDH, SDHAF1 (SDH6) и SDHAF3 (SDH7 в дрожжах), по-видимому, участвуют в созревании SDHB, защищая субъединицу или димер SDHA-SDHB от повреждения кластера Fe-S, вызванного ROS. [7]
Сборка гидрофобного якоря, состоящего из субъединиц SDHC и SDHD, остается неясной. Особенно в случае вставки гема b и даже его функции. Простетическая группа гема b, по-видимому, не является частью пути переноса электронов в комплексе II. [5] Кофактор скорее поддерживает стабильность якоря.


Многое известно о механизме окисления сукцината , который включает перенос протона и гидрида. Сочетание мутагенеза и структурного анализа идентифицирует Arg-286 субъединицы SDHA ( нумерация E. coli ) как протонный челнок. Кристаллические структуры ферментов из нескольких организмов показывают, что это хорошо подготовлено для этапа переноса протона. После этого существует два возможных механизма элиминации: E2 или E1cb. При элиминации E2 механизм согласован. Основной остаток или кофактор депротонирует альфа-углерод , а FAD принимает гидрид из бета-углерода , окисляя связанный сукцинат до фумарата — см. изображение 6. В E1cb образуется енолятный промежуточный продукт, показанный на изображении 7, прежде чем FAD принимает гидрид . Необходимы дальнейшие исследования, чтобы определить, какой механизм элиминации сукцинат проходит в сукцинатдегидрогеназе. Окисленный фумарат , теперь слабо связанный с активным центром , может свободно выходить из белка .
После того, как электроны получены из окисления сукцината через FAD , они туннелируют вдоль реле [Fe-S], пока не достигнут кластера [3Fe-4S]. Эти электроны затем переносятся в ожидающую молекулу убихинона в активном центре . Система туннелирования электронов железо - сера показана на рисунке 9.


Карбонильный кислород O1 убихинона ориентирован в активном центре (изображение 4) посредством водородных связей с Tyr83 субъединицы D. Наличие электронов в железо-серном кластере [3Fe-4S] вызывает перемещение убихинона во вторую ориентацию. Это облегчает второе водородное взаимодействие между карбонильной группой O4 убихинона и Ser27 субъединицы C. После первого этапа восстановления одного электрона образуется радикальная разновидность семихинона . Второй электрон поступает из кластера [3Fe-4S], обеспечивая полное восстановление убихинона до убихинола . Этот механизм восстановления убихинона показан на изображении 8.
Хотя функциональность гема в сукцинатдегидрогеназе все еще изучается, некоторые исследования [ кем? ] утверждают, что первый электрон, доставленный к убихинону через [3Fe-4S], может туннелировать туда и обратно между гемом и промежуточным продуктом убихинона . Таким образом, кофактор гема действует как поглотитель электронов . Его роль заключается в предотвращении взаимодействия промежуточного продукта с молекулярным кислородом с образованием активных форм кислорода (ROS). Группа гема , относительно убихинона , показана на рисунке 4.
Также было высказано предположение, что может существовать механизм пропускания, предотвращающий туннелирование электронов непосредственно в гем из кластера [3Fe-4S]. Потенциальным кандидатом является остаток His207, который находится непосредственно между кластером и гемом . His207 субъединицы B находится в непосредственной близости от кластера [3Fe-4S], связанного убихинона и гемом ; и может модулировать поток электронов между этими окислительно-восстановительными центрами. [12]
Для полного восстановления хинона в SQR необходимы два электрона и два протона . Утверждалось, что молекула воды (HOH39) поступает в активный центр и координируется His207 субъединицы B, Arg31 субъединицы C и Asp82 субъединицы D. Вид семихинона протонируется протонами, поставляемыми из HOH39, завершая восстановление убихинона до убихинола . His207 и Asp82, скорее всего, облегчают этот процесс. Другие исследования утверждают, что Tyr83 субъединицы D координируется с близлежащим гистидином , а также с карбонильным кислородом O1 убихинона . Остаток гистидина снижает pKa тирозина , делая его более подходящим для передачи своего протона восстановленному промежуточному продукту убихинона .
Существует два различных класса ингибиторов (SDHI) комплекса II: те, которые связываются в сукцинатном кармане, и те, которые связываются в убихиноновом кармане. Ингибиторы типа убихинона включают карбоксин и теноилтрифторацетон . Ингибиторы сукцинатных аналогов включают синтетическое соединение малонат , а также промежуточные продукты цикла трикарбоновых кислот, малат и оксалоацетат . Действительно, оксалоацетат является одним из самых мощных ингибиторов комплекса II. Почему распространенный промежуточный продукт цикла трикарбоновых кислот будет ингибировать комплекс II, не совсем понятно, хотя он может играть защитную роль в минимизации опосредованного обратным переносом электронов производства супероксида комплексом I. [13] Апенин 5a является высокоэффективным ингибитором комплекса II, имитирующим связывание убихинона.
Ингибиторы типа убихинона использовались в качестве фунгицидов в сельском хозяйстве с 1960-х годов. Карбоксин в основном использовался для борьбы с болезнями, вызываемыми базидиомицетами, такими как стеблевая ржавчина и заболевания Rhizoctonia . В 1980-х годах было обнаружено, что простые бензанилиды обладают сравнимой с карбоксином активностью, и ряд из них поступили в продажу, включая беноданил , флутоланил и мепронил . [14] Совсем недавно были разработаны другие соединения с более широким спектром действия против ряда фитопатогенов, включая боскалид , флуопирам , флуксапироксад , пидифлуметофен и седаксан . [15] [14] Некоторые важные для сельского хозяйства грибы нечувствительны к представителям нового поколения ингибиторов типа убихинона. [16]
FRAC имеет рабочую группу [17] по SDHI и рекомендует методы управления резистентностью . [18]
Фундаментальная роль сукцинат-кофермента Q-редуктазы как в окислительном фосфорилировании , так и в цикле лимонной кислоты делает ее жизненно важной для всех эукариотических организмов. Потеря функции SDH из-за мутаций или токсинов может вызвать широкий спектр заболеваний.
Когда SDH дисфункционален в цикле лимонной кислоты, это может привести к накоплению онкометаболита сукцината, что может привести к опухолеобразованию. Хорошо известно, что это происходит в хромаффинных клетках , вызывая нейроэндокринные опухоли, такие как параганглиома , почечная карцинома и гастроинтестинальная стромальная опухоль (GIST). [19] Данные о пенетрантности мутаций SDH, вызывающих опухолеобразование, отсутствуют, и международные руководящие принципы предполагают тщательный скрининг любых носителей. [20] Пенетрантность параганглиомы при мутациях потери функции SDH неполна и варьируется в зависимости от субъединицы. Мутации SDHB имеют пенетрантность от 8% до 37%, мутации SDHD имеют пенетрантность от 38% до 64% с некоторыми эффектами материнского импринтинга, а пенетрантность как для мутаций SDHA, так и для мутаций SDHC плохо изучена, но, вероятно, составляет от 1% до 30%. [21] [22] У млекопитающих SDH функционирует не только в генерации энергии, но и играет роль в чувствительности к кислороду . Накопление сукцината из-за дефектного SDH может вызвать псевдогипоксию и ангиогенез, оба из которых способствуют отчетливо васкулярному и характерному виду параганглиомы «соль и перец» на снимках. [23]
Биаллельные мутации потери функции SDHA, SDHB, SDHD и SDHAF1 или моноаллельные мутации потери функции SDHA могут вызвать дефицит митохондриального комплекса II . Это нарушение окислительного фосфорилирования может привести к синдрому Лея , митохондриальной энцефалопатии , атрофии зрительного нерва , миопатии и спектру заболеваний. Эти проявления могут варьироваться от смерти в течение первого года жизни или внутриутробно до легких симптомов, начинающихся во взрослом возрасте. [24]
Сниженные уровни СДГ наблюдаются посмертно в мозге пациентов с болезнью Гентингтона , а дефекты энергетического метаболизма были выявлены как у пациентов с предсимптомной, так и у пациентов с симптоматической болезнью Гентингтона. [25]
{kind=link}