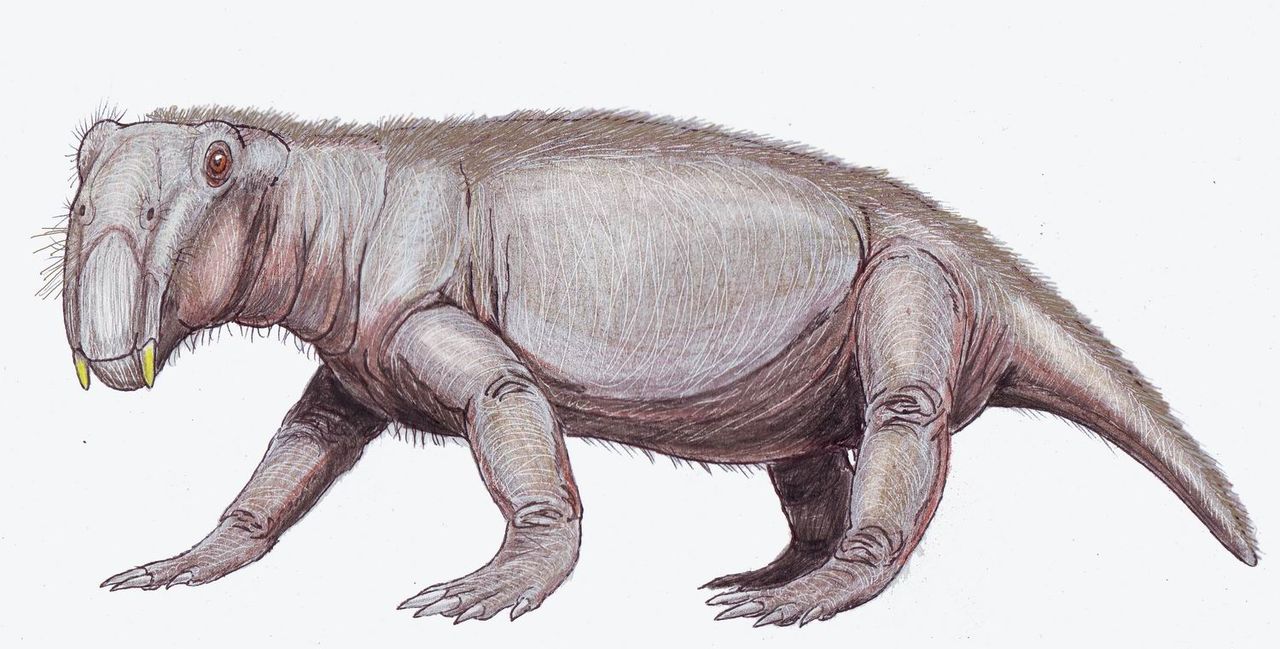
Как дицинодонт, Lystrosaurus имел только два зуба (пару похожих на бивни клыков ) и, как полагают, имел роговой клюв, который использовался для откусывания кусков растительности. Lystrosaurus был крепкого телосложения, травоядным животным. Строение его плеч и тазобедренных суставов предполагает, что Lystrosaurus передвигался полуразваленной походкой . Передние конечности были даже более крепкими, чем задние, и считается, что животное было мощным землекопом, который гнездился в норах.
Lystrosaurus пережил пермско-триасовое вымирание 252 миллиона лет назад. В раннем триасе они были самыми распространенными наземными позвоночными, составляя до 95% от общего числа особей в некоторых ископаемых слоях. [2] Исследователи выдвигали различные гипотезы о том, почему Lystrosaurus пережил вымирание и процветал в раннем триасе.
История открытия
Карта Пангеи , на которой желтыми дисками показаны места нахождения останков Lystrosaurus . Искаженные границы современных континентов показаны серыми линиями. (Распределение Lystrosaurus и трех других пермских и триасовых ископаемых групп, используемых в качестве биогеографических доказательств континентального дрейфа и определенных сухопутных мостов .)
Доктор Элиас Рут Бидл, миссионер из Филадельфии и страстный коллекционер окаменелостей, обнаружил первый череп Lystrosaurus . Бидл написал выдающемуся палеонтологу Отниелу Чарльзу Маршу , но не получил ответа. Соперник Марша, Эдвард Дринкер Коуп , был очень заинтересован в находке и описал и назвал Lystrosaurus в Трудах Американского философского общества в 1870 году. [3] Его название происходит от древнегреческих слов listron , «лопата», и sauros , «ящерица». [4] Марш с опозданием купил череп в мае 1871 года, хотя его интерес к уже описанному образцу был неясен; возможно, он хотел тщательно изучить описание и иллюстрацию Коупа. [3]
Тектоника плит
Открытие окаменелостей Lystrosaurus в Коулсэк-Блафф в Трансантарктических горах Эдвином Х. Колбертом и его командой в 1969–1970 годах помогло поддержать гипотезу тектоники плит и укрепить теорию, поскольку Lystrosaurus уже был найден в нижнем триасе южной Африки, а также в Индии и Китае. [5] [6]
Распространение и виды
Окаменелости Lystrosaurus были найдены во многих наземных костных отложениях поздней перми и раннего триаса , в основном в Африке и в меньшей степени в частях современной Индии, Китая, Монголии, европейской части России и Антарктиды (которая в то время не находилась над Южным полюсом). [7]
Виды, встречающиеся в Африке
Скелет L. murrayi
Большинство окаменелостей Lystrosaurus были найдены в формациях Бальфур и Катберг бассейна Кару в Южной Африке ; эти образцы предлагают наилучшие перспективы для идентификации видов, поскольку они наиболее многочисленны и изучаются в течение самого длительного времени. Как это часто бывает с окаменелостями, в палеонтологическом сообществе ведутся дебаты относительно того, сколько именно видов было найдено в бассейне Кару. Исследования с 1930-х по 1970-е годы предполагали большое количество (23 в одном случае). [8] Однако к 1980-м и 1990-м годам в Кару было признано только 6 видов : L. curvatus , L. platyceps , L. oviceps , L. maccaigi , L. murrayi и L. declivis . Исследование, проведенное в 2011 году, сократило это число до четырех, рассматривая ископаемые останки, ранее обозначенные как L. platyceps и L. oviceps, как представителей L. curvatus . [9]
L. maccaigi является самым крупным и, по-видимому, наиболее специализированным видом, в то время как L. curvatus был наименее специализированным. Ископаемое, похожее на Lystrosaurus , Kwazulusaurus shakai , также было найдено в Южной Африке. Хотя K. shakai не относится к тому же роду , он очень похож на L. curvatus . Поэтому некоторые палеонтологи предположили, что K. shakai , возможно, был предком или тесно связан с предками L. curvatus , в то время как L. maccaigi произошел от другой линии. [8] L. maccaigi встречается только в отложениях пермского периода и, по-видимому, не пережил пермско-триасовое вымирание . Его специализированные черты и внезапное появление в летописи окаменелостей без очевидного предка могут указывать на то, что он иммигрировал в Кару из области, в которой позднепермские отложения не были обнаружены. [8] L. curvatus обнаружен в относительно узкой полосе отложений, образовавшихся незадолго до и после вымирания, и может использоваться в качестве приблизительного маркера границы между пермским и триасовым периодами. Череп, идентифицированный как L. curvatus, был найден в позднепермских отложениях в Замбии . В течение многих лет считалось, что в Кару нет пермских образцов L. curvatus , что привело к предположениям, что L. curvatus иммигрировал из Замбии в Кару. Однако повторное исследование пермских образцов в Кару идентифицировало некоторых как L. curvatus , и нет необходимости предполагать иммиграцию. [8]
L. murrayi и L. declivis встречаются только в триасовых отложениях. [8]
Другие виды
Останки Lystrosaurus georgi были найдены в самых ранних триасовых отложениях Московского бассейна в России. Вероятно, он был тесно связан с африканским Lystrosaurus curvatus , [7] который считается одним из наименее специализированных видов и был найден в очень позднепермских и очень раннетриасовых отложениях. [8]
L. murrayi , в дополнение к двум неописанным видам, в настоящее время отнесенным к L. curvatus и L. declivis , известен из раннетриасовой формации Панчет долины Дамодар и формации Камти бассейна Пранхита-Годавари в Индии . [10] Семь видов Lystrosaurus были описаны из раннетриасовых формаций Цзюцайюань, Годикэн и Утунгоу гор Богда в Синьцзяне , Китай , хотя возможно, что только два ( L. youngi и L. hedini ) являются достоверными; необычно, что ни один китайский образец Lystrosaurus не известен ниже границы перми и триаса в этом регионе. [11] [ 12] L. curvatus , L. murrayi и L. maccaigi известны из формации Фремоув в Трансантарктических горах Антарктиды . [13]
Описание
Размер Lystrosaurus murrayi относительно человека.
Lystrosaurus был дицинодонтом терапсидом , длиной от 0,6 до 2,5 м (от 2 до 8 футов), в среднем около 0,9 м (3 фута) в зависимости от вида. [14]
В отличие от других терапсид, дицинодонты имели очень короткие морды и не имели зубов, за исключением верхних клыков , похожих на бивни . Обычно считается, что у дицинодонтов были роговые клювы , как у черепах, для срезания кусков растительности, которые затем измельчались на роговом вторичном небе, когда рот был закрыт. Челюстной сустав был слабым и двигался вперед и назад с режущим действием, вместо более распространенных боковых или вверх-вниз движений. Считается, что челюстные мышцы были прикреплены необычно далеко вперед на черепе и занимали много места на верхней и задней части черепа. В результате глаза были посажены высоко и далеко вперед на черепе, а лицо было коротким. [15]
Особенности скелета указывают на то, что Lystrosaurus передвигался полуразваливающейся походкой . Нижний задний угол лопатки ( плечевой кости) был сильно окостеневшим (состоял из прочной кости), что говорит о том, что движение лопатки способствовало длине шага передних конечностей и уменьшало боковое сгибание тела. [7] Пять крестцовых позвонков были массивными, но не срослись друг с другом и с тазом , что делало спину более жесткой и уменьшало боковое сгибание во время ходьбы животного. Считается, что терапсиды с менее чем пятью крестцовыми позвонками имели раскинутые конечности, как у современных ящериц. [7] У динозавров и млекопитающих , у которых конечности прямостоящие, крестцовые позвонки срослись друг с другом и с тазом . [16] Считается, что опора над каждой вертлужной впадиной (тазобедренной впадиной) предотвращала вывих бедренной кости (бедренной кости), когда Lystrosaurus ходил полуразвалившись. [7] Передние конечности Lystrosaurus были массивными, [7] и считается, что Lystrosaurus был мощным роющим животным.
Мумифицированные образцы, найденные в бассейне Кару и описанные в 2022 году, показали, что у Lystrosaurus была ямчатая, кожистая и безволосая кожа. [17] [18]
Палеоэкология
Доминирование раннего триаса
Lystrosaurus georgi
Lystrosaurus примечателен тем, что доминировал на юге Пангеи в течение миллионов лет в раннем триасе . По крайней мере один неопознанный вид этого рода пережил массовое вымирание в конце пермского периода и, в отсутствие хищников и травоядных конкурентов, продолжал процветать и повторно распался на ряд видов внутри рода, [19] став наиболее распространенной группой наземных позвоночных в раннем триасе; некоторое время 95% наземных позвоночных были Lystrosaurus . [19] [20] Это единственный случай, когда один вид или род наземных животных доминировал на Земле в такой степени. [21] Несколько других родов пермских терапсид также пережили массовое вымирание и появляются в триасовых породах — тероцефалы Tetracynodon , Moschorhinus , Ictidosuchoides и Promoschorhynchus — но, по-видимому, не были широко распространены в триасе; [8] [22] Полное экологическое восстановление заняло 30 миллионов лет, охватывая ранний и средний триас . [23]
Было предпринято несколько попыток объяснить, почему Lystrosaurus пережил пермско-триасовое вымирание , «мать всех массовых вымираний» [24] , и почему он доминировал в раннетриасовой фауне в такой беспрецедентной степени:
Lystrosaurus murrayi
Следы роста на окаменелых бивнях указывают на то, что Lystrosaurus , живший в Антарктиде ~250 млн лет назад, мог впадать в длительное состояние оцепенения , аналогичное спячке . Это может быть самым древним свидетельством состояния, похожего на спячку, у позвоночных животных и указывает на то, что оцепенение возникло у позвоночных до того, как эволюционировали млекопитающие и динозавры. [25] [26] [27]
Одна из последних теорий заключается в том, что вымирание привело к снижению содержания кислорода в атмосфере и повышению содержания углекислого газа , так что многие наземные виды вымерли, потому что им было слишком трудно дышать. [20] Поэтому было высказано предположение, что Lystrosaurus выжил и стал доминирующим, потому что его роющий образ жизни позволил ему справиться с атмосферой «застоявшегося воздуха», и что определенные особенности его анатомии были частью этой адаптации: бочкообразная грудь, вмещающая большие легкие, короткие внутренние ноздри , которые способствовали быстрому дыханию, и высокие невральные отростки (выступы на дорсальной стороне позвонков ) , которые давали больше рычагов мышцам, которые расширяли и сокращали его грудь. Однако во всех этих пунктах есть слабые стороны: грудь Lystrosaurus была не значительно больше по сравнению с его размером, чем у других вымерших дицинодонтов ; Хотя триасовые дицинодонты, по-видимому, имели более длинные невральные отростки, чем их пермские аналоги, эта особенность может быть связана с позой, передвижением или даже размером тела, а не с эффективностью дыхания; L. murrayi и L. declivis гораздо более многочисленны, чем другие раннетриасовые роющие животные, такие как Procolophon или Thrinaxodon . [8]
Схема скелета листрозавра
Предположение о том, что Lystrosaurus выжил и доминировал благодаря своему полуводному образу жизни, имеет схожий недостаток: хотя темноспондилы стали более распространенными в триасовых отложениях Карру, их было гораздо меньше, чем L. murrayi и L. declivis . [8]
Наиболее специализированные и крупные животные подвергаются большему риску при массовых вымираниях; это может объяснить, почему неспециализированный L. curvatus выжил, в то время как более крупный и специализированный L. maccaigi вымер вместе со всеми другими крупными пермскими травоядными и плотоядными. [8] Хотя Lystrosaurus в целом выглядит приспособленным к питанию растениями, похожими на Dicroidium , который доминировал в раннем триасе, более крупные размеры L. maccaigi , возможно, заставили его полагаться на более крупных представителей флоры Glossopteris , которые не пережили вымирание в конце пермского периода. [8]
Только тероцефал Moschorhinus длиной 1,5 м (5 футов) и крупный архозаврообразный Proterosuchus , по-видимому, были достаточно крупными, чтобы охотиться на триасовый вид Lystrosaurus , и эта нехватка хищников могла быть причиной всплеска популяции Lystrosaurus в раннем триасе. [8]
По словам Бентона, «Возможно, выживание Lystrosaurus было просто делом удачи» [19] .
^ Клувер, Майкл Альберт (1978). Ископаемые рептилии южноафриканского Кару . Южноафриканский музей. ISBN 9780908407583.
^ Дамиани, Р. Дж.; Невелинг, Дж.; Модесто, С. П. и Йейтс, А. М. (2004). «Барендскрааль, разнообразное местонахождение амниот из зоны скопления Lystrosaurus , ранний триас Южной Африки». Palaeontologia Africana . 39 : 53–62.
^ ab Wallace, David Rains (2000). Месть охотников за костями: динозавры, жадность и величайшая научная вражда позолоченного века . Houghton Mifflin Harcourt. стр. 44–45. ISBN978-0-618-08240-7.
^ Любик, Наоми (февраль 2005 г.). «Исследование Антарктики». Geotimes. Американский институт геонаук – через agiweb.org.
^ Тревик, Стив (2016). «Тектоника плит в биогеографии». Международная энциклопедия географии: люди, Земля, окружающая среда и технологии . John Wiley & Sons, Ltd. стр. 1–9. doi :10.1002/9781118786352.wbieg0638. ISBN9781118786352.
^ abcdef Сурков, М.В.; Каландадзе, Н.Н. и Бентон, MJ (июнь 2005 г.). «Lystrosaurus georgi, дицинодонт из нижнего триаса России» (PDF) . Журнал палеонтологии позвоночных . 25 (2): 402–413. doi :10.1671/0272-4634(2005)025[0402:LGADFT]2.0.CO;2. ISSN 0272-4634. S2CID 59028100. Архивировано из оригинала (PDF) 19 декабря 2008 года . Проверено 7 июля 2008 г.
^ abcdefghijkl Бота, Дж. и Смит, RMH (2005). « Видовой состав Lystrosaurus на границе пермо-триаса в бассейне Кару в Южной Африке». Летайя . 40 (2): 125–137. дои : 10.1111/j.1502-3931.2007.00011.x. Полная версия доступна на сайте Botha; Smith (2007). Состав видов Lystrosaurus на границе пермо-триаса в бассейне Кару в Южной Африке (PDF) (Отчет). Архивировано из оригинала (PDF) 10 сентября 2008 года . Получено 2 июля 2008 года .
^ Грин, FE; Форстер, Калифорния; Клювер, Массачусетс, и Георгий, Дж. А. (2006). Палеобиология амниот: перспективы эволюции млекопитающих, птиц и рептилий . Издательство Чикагского университета. стр. 432–503.
^ Bandyopadhyay, Saswati; Ray, Sanghamitra (1 марта 2020 г.). «Гондванская фауна позвоночных Индии: их разнообразие и межконтинентальные связи». Episodes Journal of International Geoscience . 43 (1): 438–460. doi : 10.18814/epiiugs/2020/020028 .
^ Angielczyk, Kenneth D.; Liu, Jun; Sidor, Christian A.; Yang, Wan (1 ноября 2022 г.). «Стратиграфические и географические проявления пермо-триасовых тетрапод в горах Богда, северо-запад Китая — последствия новой циклостратиграфической структуры и байесовской возрастной модели». Журнал африканских наук о Земле . 195 : 104655. doi : 10.1016/j.jafrearsci.2022.104655 . ISSN 1464-343X.
^ Хань, Фэнлу; Чжао, Ци; Лю, Цзюнь (18 марта 2021 г.). «Предварительный гистологический анализ костей Lystrosaurus (Therapsida: Dicynodontia) из нижнего триаса Северного Китая и его значение для образа жизни и окружающей среды после вымирания в конце пермского периода». PLoS One . 16 (3): e0248681. doi : 10.1371/journal.pone.0248681 . ISSN 1932-6203. PMC 7971864. PMID 33735263 .
^ "Lystrosaurus". Encyclopaedia Britannica . Fossils & Facts . Получено 18 марта 2019 г.
^ Коуэн, Р. (2000). История жизни (3-е изд.). Blackwell Scientific. стр. 167–168. ISBN978-0-632-04444-3.
^ Бентон, Майкл Дж. (2004). «Происхождение и связи динозавров». В Weishampel, Дэвид Б.; Додсон, Питер и Осмольска, Гальска (ред.). Динозавры (2-е изд.). Беркли, Калифорния: Издательство Калифорнийского университета. стр. 7–19. ISBN978-0-520-24209-8.
^ Смит, Р.М. Х.; Бота, Дж.; Виджилетти, ПА (2022). «Тафономия засушливых тетрапод в раннетриасовом бассейне Кару, Южная Африка». Палеогеография, палеоклиматология, палеоэкология . 604. 111207. Bibcode : 2022PPP...60411207S. doi : 10.1016/j.palaeo.2022.111207. S2CID 251781291.
^ «Эти ископаемые мумии раскрывают жестокий мир задолго до того, как появился тираннозавр». gizmodo.com . 20 сентября 2022 г.
^ abc Бентон, М.Дж. (2006). Когда жизнь почти умерла: величайшее массовое вымирание всех времен . Лондон, Великобритания: Thames & Hudson. ISBN978-0-500-28573-2.
^ ab Cokinos, Christopher (12 октября 2007 г.) [Май-июнь 2007 г.]. "Утешения исчезновения". Orion . Архивировано из оригинала 12 октября 2007 г.
^ Huttenlocker, AK; Sidor, CA; Smith, RMH (2011). «Новый образец Promoschorhynchus (Therapsida: Therocephalia: Akidnognathidae) из нижнего триаса Южной Африки и его значение для выживания териодонтов через пермо-триасовую границу». Journal of Vertebrate Paleontology . 31 (2): 405–421. Bibcode :2011JVPal..31..405H. doi :10.1080/02724634.2011.546720. S2CID 129242450.
^ Sahney, S. & Benton, MJ (2008). «Восстановление после самого глубокого массового вымирания всех времен». Труды Королевского общества B: Биологические науки . 275 (1636): 759–65. doi :10.1098/rspb.2007.1370. PMC 2596898. PMID 18198148 .
^ Эрвин, Д. Х. (1993). Великий палеозойский кризис: жизнь и смерть в пермском периоде . Columbia University Press. ISBN978-0-231-07467-4.
^ «Ископаемые свидетельства состояния, похожего на спячку, у антарктического животного возрастом 250 миллионов лет». phys.org . Август 2020 г. Получено 7 сентября 2020 г.
^ "Ископаемые останки предполагают, что животные впадают в спячку уже 250 миллионов лет". UPI . Получено 7 сентября 2020 г. .
^ Уитни, Меган Р.; Сидор, Кристиан А. (27 августа 2020 г.). «Доказательства оцепенения в бивнях Lystrosaurus из раннего триаса Антарктиды». Communications Biology . 3 (1): 471. doi :10.1038/s42003-020-01207-6. ISSN 2399-3642. PMC 7453012. PMID 32855434 . Текст и изображения доступны по лицензии Creative Commons Attribution 4.0 International.
Внешние ссылки
На Викискладе есть медиафайлы по теме Lystrosaurus .
"Dicynodontia". Palaeos.com . 400.725. Архивировано из оригинала 22 февраля 2006 г.
Рэнс, Хью. "Млекопитающие рептилии Пангеи". geowords.com . Настоящее — ключ к прошлому. Архивировано из оригинала 27 апреля 2006 г.