
Видимый спектр — это полоса электромагнитного спектра , видимая человеческому глазу . Электромагнитное излучение в этом диапазоне длин волн называется видимым светом (или просто светом). Оптический спектр иногда считают тем же самым, что и видимый спектр, но некоторые авторы определяют этот термин более широко, включая ультрафиолетовую и инфракрасную части электромагнитного спектра, известные под общим названием оптическое излучение . [1]
Типичный человеческий глаз реагирует на длины волн от 380 до 750 нанометров . [2] С точки зрения частоты это соответствует полосе в районе 400–790 терагерц . Эти границы не являются четко определенными и могут варьироваться в зависимости от человека. [3] При оптимальных условиях эти пределы человеческого восприятия могут простираться до 310 нм (ультрафиолет) и 1100 нм (ближний инфракрасный). [4] [5] [6]
Спектр не содержит всех цветов , которые может различить зрительная система человека . Ненасыщенные цвета, такие как розовый , или фиолетовые вариации, такие как пурпурный , например, отсутствуют, поскольку они могут быть получены только из смеси нескольких длин волн. Цвета, содержащие только одну длину волны, также называются чистыми цветами или спектральными цветами . [7]
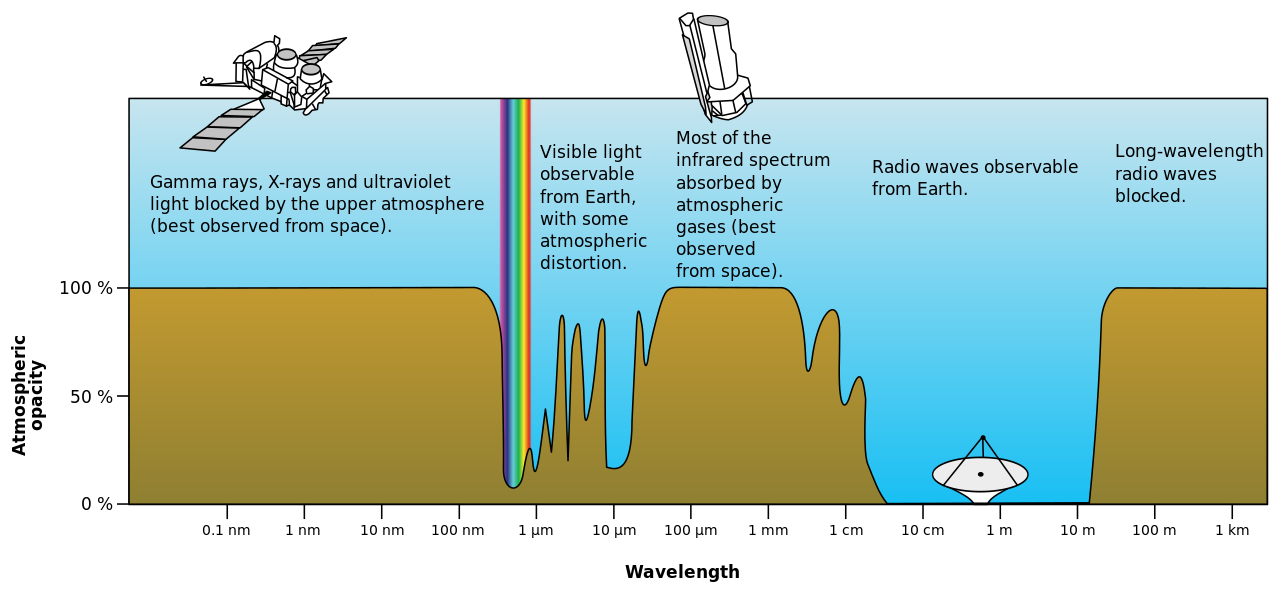
Видимые длины волн проходят в основном без ослабления через атмосферу Земли через область « оптического окна » электромагнитного спектра. Примером этого явления является случай, когда чистый воздух рассеивает синий свет больше, чем красный, и поэтому полуденное небо кажется синим (за исключением области вокруг Солнца, которая кажется белой, потому что свет рассеивается не так сильно). Оптическое окно также называют «видимым окном», потому что оно перекрывает спектр видимой реакции человека. Окно ближнего инфракрасного диапазона (NIR) находится прямо за пределами человеческого зрения, как и окно средневолнового инфракрасного диапазона (MWIR) и окно длинноволнового или дальнего инфракрасного диапазона (LWIR или FIR), хотя другие животные могут их воспринимать.
Цвета, которые могут быть получены с помощью видимого света узкой полосы длин волн ( монохроматический свет ), называются чистыми спектральными цветами . Различные цветовые диапазоны, указанные на рисунке, являются приблизительными: спектр непрерывен, без четких границ между одним цветом и другим. [8]

В XIII веке Роджер Бэкон выдвинул теорию, что радуга возникает в результате процесса, аналогичного прохождению света через стекло или кристалл. [9]
В 17 веке Исаак Ньютон открыл, что призмы могут разбирать и собирать белый свет, и описал это явление в своей книге «Оптика» . Он был первым, кто использовал слово спектр ( лат. «появление» или «привидение») в этом смысле в печати в 1671 году, описывая свои эксперименты по оптике . Ньютон заметил, что когда узкий луч солнечного света падает на поверхность стеклянной призмы под углом, часть отражается , а часть луча проходит в стекло и сквозь него, появляясь в виде разноцветных полос. Ньютон выдвинул гипотезу, что свет состоит из «корпускул» (частиц) разных цветов, причем разные цвета света движутся с разной скоростью в прозрачном веществе, красный свет движется быстрее фиолетового в стекле. В результате красный свет преломляется ( преломляется ) менее резко, чем фиолетовый, когда он проходит через призму, создавая спектр цветов.

Первоначально Ньютон разделил спектр на шесть названных цветов: красный , оранжевый , желтый , зеленый , синий и фиолетовый . Позже он добавил индиго в качестве седьмого цвета, поскольку считал, что семь — это совершенное число, полученное от древнегреческих софистов , о существовании связи между цветами, музыкальными нотами, известными объектами в Солнечной системе и днями недели. [10] Человеческий глаз относительно нечувствителен к частотам индиго, и некоторые люди, которые в остальном обладают хорошим зрением, не могут отличить индиго от синего и фиолетового. По этой причине некоторые более поздние комментаторы, включая Айзека Азимова , [11] предположили, что индиго не следует рассматривать как цвет сам по себе, а просто как оттенок синего или фиолетового. Факты указывают на то, что то, что Ньютон подразумевал под «индиго» и «синим», не соответствует современным значениям этих цветовых слов. Сравнение наблюдений Ньютона за призматическими цветами с цветным изображением видимого спектра света показывает, что «индиго» соответствует тому, что сегодня называется синим, тогда как его «синий» соответствует циану . [12] [13] [14]
В XVIII веке Иоганн Вольфганг фон Гёте писал об оптических спектрах в своей «Теории цветов» . Гёте использовал слово спектр ( Spektrum ) для обозначения призрачного оптического остаточного изображения , как и Шопенгауэр в «О зрении и цветах» . Гёте утверждал, что непрерывный спектр — это составное явление. Там, где Ньютон сузил луч света, чтобы изолировать явление, Гёте заметил, что более широкая апертура создает не спектр, а скорее красновато-жёлтые и сине-голубые края с белым между ними. Спектр появляется только тогда, когда эти края достаточно близки, чтобы перекрываться.
В начале 19 века концепция видимого спектра стала более определенной, поскольку свет за пределами видимого диапазона был открыт и охарактеризован Уильямом Гершелем ( инфракрасный ) и Иоганном Вильгельмом Риттером ( ультрафиолетовый ), Томасом Юнгом , Томасом Иоганном Зеебеком и другими. [15] Юнг был первым, кто измерил длины волн различных цветов света в 1802 году. [16]
Связь между видимым спектром и цветовым зрением была исследована Томасом Юнгом и Германом фон Гельмгольцем в начале 19 века. Их теория цветового зрения правильно предполагала, что глаз использует три различных рецептора для восприятия цвета.

Видимый спектр ограничен длинами волн, которые могут как достигать сетчатки, так и вызывать зрительную фототрансдукцию (возбуждать зрительный опсин ). Нечувствительность к УФ-свету, как правило, ограничивается передачей через хрусталик . Нечувствительность к ИК-свету ограничивается функциями спектральной чувствительности зрительных опсинов. Диапазон определяется психометрически функцией световой эффективности , которая учитывает все эти факторы. У людей существует отдельная функция для каждой из двух зрительных систем, одна для фотопического зрения , используемого при дневном свете, которое опосредовано колбочками , и одна для скотопического зрения , используемого при тусклом свете, которое опосредовано палочками . Каждая из этих функций имеет различные видимые диапазоны. Однако обсуждение видимого диапазона обычно предполагает фотопическое зрение.
Видимый диапазон большинства животных эволюционировал, чтобы соответствовать оптическому окну , которое является диапазоном света, который может проходить через атмосферу. Озоновый слой поглощает почти весь ультрафиолетовый свет (ниже 315 нм). [17] Однако это влияет только на космический свет (например, солнечный свет ), а не на земной свет (например, биолюминесценцию ).
Прежде чем достичь сетчатки , свет должен сначала пройти через роговицу и хрусталик . UVB-свет (< 315 нм) фильтруется в основном роговицей, а UVA-свет (315–400 нм) фильтруется в основном хрусталиком. [18] Хрусталик также желтеет с возрастом, сильнее всего ослабляя передачу в синей части спектра. [18] Это может вызвать ксантопсию , а также небольшое усечение коротковолнового (синего) предела видимого спектра. У людей с афакией отсутствует хрусталик, поэтому UVA-свет может достигать сетчатки и возбуждать зрительные опсины; это расширяет видимый диапазон и также может привести к цианопсии .
Каждый опсин имеет спектральную функцию чувствительности, которая определяет вероятность поглощения им фотона каждой длины волны. Функция световой эффективности приблизительно является суперпозицией вносящих вклад визуальных опсинов . Таким образом, дисперсия в положении отдельных функций спектральной чувствительности опсинов влияет на функцию световой эффективности и видимый диапазон. Например, длинноволновый (красный) предел изменяется пропорционально положению L-опсина. Положения определяются пиковой длиной волны (длина волны наибольшей чувствительности), так как пиковая длина волны L-опсина в синюю область смещается на 10 нм, длинноволновый предел видимого спектра также смещается на 10 нм. Большие отклонения пиковой длины волны L-опсина приводят к форме цветовой слепоты , называемой протаномалией , а отсутствующий L-опсин ( протанопия ) укорачивает видимый спектр примерно на 30 нм на длинноволновом пределе. Формы дальтонизма, затрагивающие М-опсин и S-опсин, не оказывают существенного влияния на функцию световой эффективности или пределы видимого спектра.
Независимо от фактической физической и биологической дисперсии, определение пределов не является стандартным и будет меняться в зависимости от отрасли. Например, некоторые отрасли могут быть озабочены практическими пределами, поэтому консервативно сообщат 420–680 нм, [19] [20] в то время как другие могут быть озабочены психометрией и достижением самого широкого спектра, щедро сообщат 380–750 или даже 380–800 нм. [21] [22] Функция световой эффективности в ближнем ИК-диапазоне не имеет жесткого отсечения, а скорее экспоненциального спада, так что значение функции (или чувствительность зрения) при 1050 нм примерно в 10 9 раз слабее, чем при 700 нм; поэтому для восприятия света 1050 нм требуется гораздо более высокая интенсивность, чем при 700 нм. [23]
В идеальных лабораторных условиях субъекты могут воспринимать инфракрасный свет длиной волны не менее 1064 нм. [23] В то время как ближний инфракрасный свет с длиной волны 1050 нм может вызывать красный цвет, что предполагает прямое поглощение L-опсином, также имеются сообщения о том, что импульсные ближние инфракрасные лазеры могут вызывать зеленый цвет, что предполагает, что двухфотонное поглощение может обеспечивать расширенную чувствительность к ближнему инфракрасному свету. [23]
Аналогично, молодые люди могут воспринимать ультрафиолетовые волны длиной до 310–313 нм, [24] [25] [26], но обнаружение света ниже 380 нм может быть связано с флуоресценцией глазных сред, а не с прямым поглощением УФ-света опсинами. Поскольку свет UVA поглощается глазными средами (хрусталиком и роговицей), он может флуоресцировать и высвобождаться с более низкой энергией (более длинная волна), которая затем может быть поглощена опсинами. Например, когда хрусталик поглощает свет длиной 350 нм, спектр флуоресцентного излучения центрируется на 440 нм. [27]
Помимо фотопической и скотопической систем, у людей есть и другие системы для обнаружения света, которые не вносят вклад в первичную зрительную систему . Например, меланопсин имеет диапазон поглощения 420–540 нм и регулирует циркадный ритм и другие рефлекторные процессы. [28] Поскольку система меланопсин не формирует изображения, она, строго говоря, не считается зрением и не вносит вклад в видимый диапазон.
Видимый спектр определяется как видимый для человека, но дисперсия между видами велика. Не только опсины колбочек могут быть спектрально смещены для изменения видимого диапазона, но позвоночные с 4 колбочками (тетрахроматические) или 2 колбочками (дихроматические) относительно человеческих 3 (трихроматические) также будут иметь более широкий или более узкий видимый спектр, чем люди, соответственно.
У позвоночных обычно имеется от 1 до 4 различных классов опсинов: [17]
Тестирование зрительной системы животных на основе поведения затруднительно, поэтому диапазон видимого света животных обычно оценивается путем сравнения пиковых длин волн опсинов с таковыми у типичных людей (S-опсин при 420 нм и L-опсин при 560 нм).
Большинство млекопитающих сохранили только два класса опсинов (LWS и VS), вероятно, из-за ночного «бутылочного горлышка» . Однако приматы Старого Света (включая людей) с тех пор развили две версии в классе LWS, чтобы восстановить трихроматию. [17] В отличие от большинства млекопитающих, UVS-опсины грызунов остались на более коротких длинах волн. Наряду с отсутствием УФ-фильтров в хрусталике, у мышей есть UVS-опсин, который может обнаруживать вплоть до 340 нм. Хотя попадание УФ-света на сетчатку может привести к ее повреждению, короткая продолжительность жизни мышей по сравнению с другими млекопитающими может минимизировать этот недостаток по сравнению с преимуществом УФ-зрения. [29] У собак есть два колбочковых опсина на 429 нм и 555 нм, поэтому они видят почти весь видимый спектр человека, несмотря на то, что они дихроматичны. [30] У лошадей есть два колбочковых опсина на 428 нм и 539 нм, что дает немного более усеченное красное зрение. [31]
Большинство других позвоночных (птицы, ящерицы, рыбы и т. д.) сохранили свою тетрахроматию , включая UVS-опсины, которые простираются дальше в ультрафиолет, чем человеческий VS-опсин. [17] Чувствительность птичьих UVS-опсинов сильно варьируется: от 355 до 425 нм, а LWS-опсинов — от 560 до 570 нм. [32] Это приводит к тому, что некоторые птицы имеют видимый спектр наравне с людьми, а другие птицы имеют значительно расширенную чувствительность к УФ-свету. Иногда сообщается, что LWS-опсин птиц имеет пиковую длину волны выше 600 нм, но это эффективная пиковая длина волны, которая включает фильтр капель птичьего жира . [32] Пиковая длина волны LWS-опсина сама по себе является лучшим предиктором предела длинных волн. Возможным преимуществом птичьего УФ-зрения являются зависящие от пола отметины на их оперении , которые видны только в ультрафиолетовом диапазоне. [33] [34]
Костистые рыбы (костные рыбы) в целом тетрахроматичны. Чувствительность опсинов UVS рыб варьируется от 347 до 383 нм, а опсинов LWS от 500 до 570 нм. [35] Однако некоторые рыбы, использующие альтернативные хромофоры, могут расширить чувствительность опсинов LWS до 625 нм. [35] Распространенное мнение о том, что обыкновенная золотая рыбка — единственное животное, способное видеть как инфракрасный, так и ультрафиолетовый свет [36] , неверно, поскольку золотая рыбка не может видеть инфракрасный свет. [37]
Зрительные системы беспозвоночных сильно отличаются от позвоночных, поэтому прямые сравнения затруднены. Однако чувствительность к ультрафиолету была зарегистрирована у большинства видов насекомых. [38] Пчелы и многие другие насекомые могут обнаруживать ультрафиолетовый свет, который помогает им находить нектар в цветах. Виды растений, которые зависят от опыления насекомыми, могут быть обязаны репродуктивному успеху своим появлением в ультрафиолетовом свете, а не тем, насколько красочными они кажутся людям. Предел длинных волн пчел составляет около 590 нм. [39] Креветки-богомолы демонстрируют до 14 опсинов, что обеспечивает видимый диапазон от менее 300 нм до более 700 нм. [17]
Некоторые змеи могут «видеть» [40] лучистое тепло на длинах волн от 5 до 30 мкм с такой степенью точности, что слепая гремучая змея может нацелиться на уязвимые части тела добычи, на которую она нападает, [41] а другие змеи с этим органом могут обнаруживать теплые тела на расстоянии метра. [42] Он также может использоваться для терморегуляции и обнаружения хищников . [43] [44]
Спектроскопия — это изучение объектов на основе спектра цвета, который они испускают, поглощают или отражают. Спектроскопия видимого света является важным инструментом в астрономии (как и спектроскопия на других длинах волн), где ученые используют ее для анализа свойств удаленных объектов. Химические элементы и малые молекулы могут быть обнаружены в астрономических объектах путем наблюдения линий излучения и линий поглощения . Например, гелий был впервые обнаружен путем анализа спектра Солнца . Сдвиг частоты спектральных линий используется для измерения доплеровского сдвига ( красного или синего смещения ) удаленных объектов для определения их скоростей к наблюдателю или от него. Астрономическая спектроскопия использует высокодисперсные дифракционные решетки для наблюдения спектров с очень высоким спектральным разрешением.
призма Роджера Бэкона.
Излучение ниже 320 нм [ультрафиолет (УФ)А] в значительной степени экранируется озоновым слоем в верхней атмосфере Земли и поэтому недоступно для зрительной системы,
Была измерена фовеолярная чувствительность к нескольким длинам волн лазера ближнего инфракрасного диапазона. Было обнаружено, что глаз может реагировать на излучение с длиной волны не менее 1064 нм. Непрерывный источник лазера 1064 нм казался красным, но импульсный источник лазера 1060 нм казался зеленым, что предполагает наличие генерации второй гармоники в сетчатке.
Пределы общего диапазона чувствительности глаза простираются примерно от 310 до 1050 нанометров
Обычно человеческий глаз реагирует на световые лучи от 390 до 760 нм. Этот диапазон может быть расширен до 310–1050 нм в искусственных условиях.