Вестибулоокулярный рефлекс ( VOR ) — это рефлекс , который стабилизирует взгляд во время движения головы, при этом движение глаз происходит из-за активации вестибулярной системы , его также называют шейно-глазным рефлексом. Рефлекс стабилизирует изображение на сетчатке глаза во время движения головы. Взгляд удерживается на месте, производя движения глаз в направлении, противоположном движению головы. Например, когда голова движется вправо, глаза движутся влево, то есть изображение , которое видит человек, остается прежним, даже если голова повернулась. Поскольку небольшое движение головы присутствует все время, VOR необходим для стабилизации зрения: людям с нарушенным рефлексом трудно читать с помощью печати, потому что глаза не стабилизируются при небольшом дрожании головы, а также потому, что повреждение рефлекса может вызвать нистагм . [1]
VOR не зависит от того, что видно. Он также может быть активирован горячей или холодной стимуляцией внутреннего уха , где находится вестибулярная система, и работает даже в полной темноте или при закрытых глазах. Однако при наличии света к движению добавляется рефлекс фиксации . [2] Большинство признаков VOR присутствуют у котят, выращенных в полной темноте. [3]
У низших животных органы, координирующие равновесие и движение, не являются независимыми от движения глаз. Например, рыба двигает глазами рефлекторно, когда ее хвост двигается. У людей есть полукружные каналы , рецепторы «растяжения» шейных мышц и утрикулюс (орган гравитации). Хотя полукружные каналы вызывают большинство рефлексов, которые реагируют на ускорение, поддержание равновесия опосредовано растяжением шейных мышц и тягой гравитации на утрикулюс (отолитовый орган) внутреннего уха. [2]
VOR имеет как вращательные, так и поступательные аспекты. Когда голова вращается вокруг любой оси (горизонтальной, вертикальной или торсионной), удаленные зрительные образы стабилизируются вращением глаз вокруг той же оси, но в противоположном направлении. [4] Когда голова транслируется, например, во время ходьбы, точка визуальной фиксации поддерживается вращением направления взгляда в противоположном направлении, [5] на величину, которая зависит от расстояния. [6]
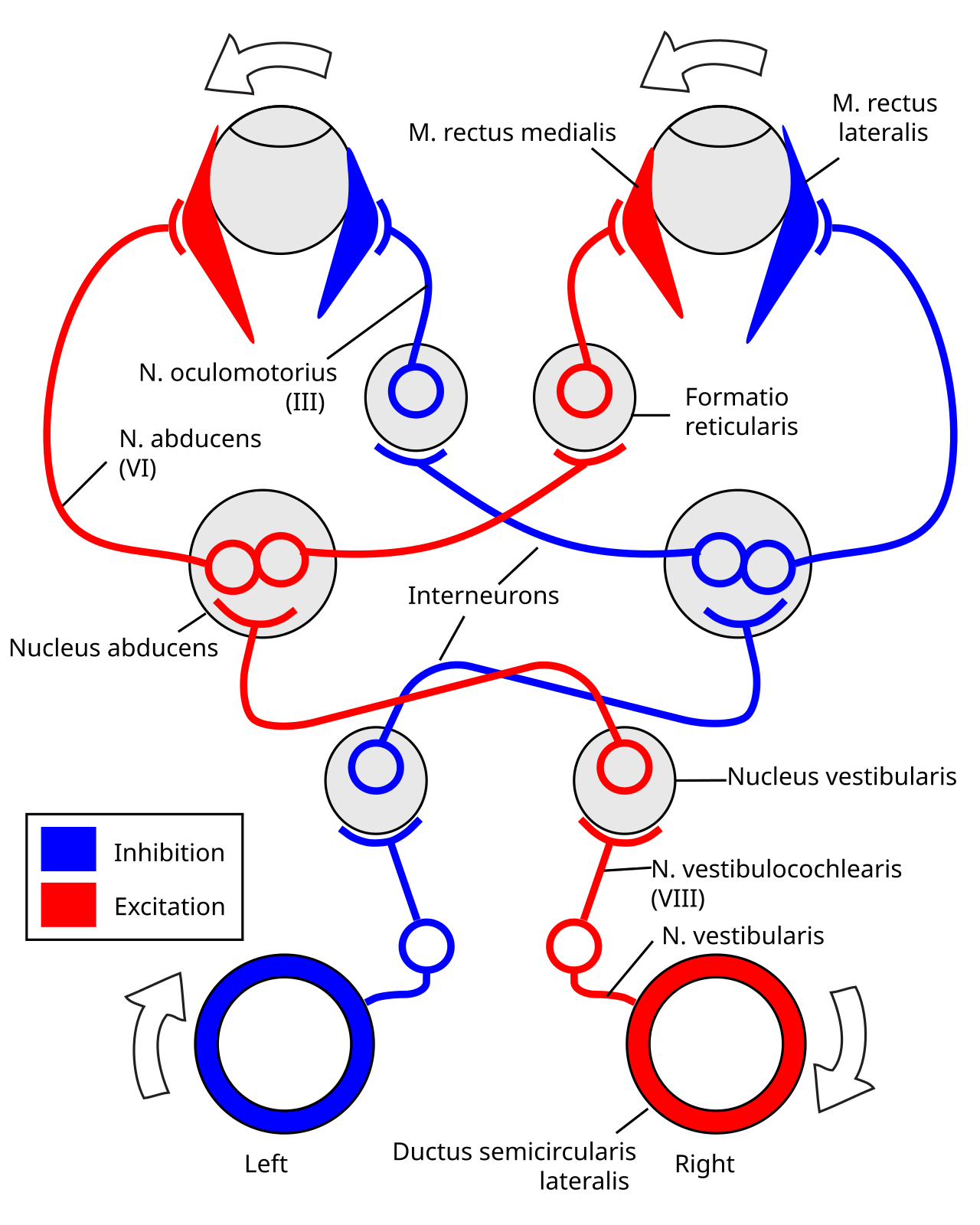
Вестибулоокулярный рефлекс управляется сигналами, исходящими из вестибулярной системы внутреннего уха. Полукружные каналы обнаруживают поворот головы и обеспечивают вращательный компонент, тогда как отолиты обнаруживают перемещение головы и управляют трансляционным компонентом. Сигнал для горизонтального вращательного компонента проходит по вестибулярному нерву через вестибулярный ганглий и заканчивается в вестибулярных ядрах в стволе мозга . Из этих ядер волокна пересекаются с отводящим ядром противоположной стороны мозга. Здесь волокна синапсируют с 2 дополнительными путями. Один путь проецируется непосредственно на латеральную прямую мышцу глаза через отводящий нерв. Другой нервный тракт проецируется от отводящего ядра через медиальный продольный пучок к глазодвигательному ядру противоположной стороны, которое содержит двигательные нейроны , которые управляют активностью глазных мышц, в частности активируя медиальную прямую мышцу глаза через глазодвигательный нерв .
Другой путь (не на рисунке) напрямую проецируется от вестибулярного ядра через восходящий тракт Дейтерса к двигательному нейрону медиальной прямой мышцы той же стороны. Кроме того, существуют тормозные вестибулярные пути к ипсилатеральному ядру отводящего нерва. Однако прямого пути от вестибулярного нейрона к двигательному нейрону медиальной прямой мышцы не существует. [7]
Аналогичные пути существуют для вертикальных и крутильных компонентов VOR.
В дополнение к этим прямым путям, которые управляют скоростью вращения глаз, существует непрямой путь, который создает сигнал положения, необходимый для предотвращения отката глаза назад к центру, когда голова перестает двигаться. Этот путь особенно важен, когда голова движется медленно, потому что здесь сигналы положения доминируют над сигналами скорости. Дэвид А. Робинсон обнаружил, что глазным мышцам требуется этот двойной привод скорости-положения, и также предположил, что он должен возникать в мозге путем математической интеграции сигнала скорости и последующей отправки результирующего сигнала положения мотонейронам. Робинсон был прав: «нейронный интегратор» для горизонтального положения глаз был обнаружен в ядре prepositus hypoglossi [8] в продолговатом мозге, а нейронный интегратор для вертикального и торсионного положения глаз был обнаружен в интерстициальном ядре Кахаля [9] в среднем мозге. Те же самые нейронные интеграторы также генерируют положение глаз для других сопряженных движений глаз, таких как саккады и плавное преследование .
Интегратор имеет утечку , с характерным временем утечки 20 с. Например, когда субъект сидит неподвижно и фокусируется на объекте, и внезапно выключается свет, глаза вернутся в нейтральное положение примерно через 40 секунд, даже если субъект пытается сохранить фокус. [10] [11]
Например, если голова повернута по часовой стрелке, как видно сверху, то возбуждающие импульсы посылаются из полукружного канала с правой стороны через вестибулярный нерв через ганглий Скарпы и заканчиваются в правых вестибулярных ядрах в стволе мозга. Из этих ядер возбуждающие волокна переходят в левое отводящее ядро. Там они проецируются и стимулируют латеральную прямую мышцу левого глаза через отводящий нерв. Кроме того, посредством медиального продольного пучка и глазодвигательных ядер они активируют медиальные прямые мышцы правого глаза. В результате оба глаза повернутся против часовой стрелки.
Кроме того, некоторые нейроны правого вестибулярного ядра напрямую стимулируют двигательные нейроны правой медиальной прямой мышцы и ингибируют правое отводящее ядро.
VOR управляется нейронным интегратором. Нейрон из каждого горизонтального полукружного канала срабатывает с частотой , где — измеренная горизонтальная угловая скорость полукружного канала. Мотонейрон, управляющий горизонтальными глазными мышцами, срабатывает с частотой , где — горизонтальный угол поворота, а — его горизонтальная угловая скорость. Эти два термина учитывают эластичность и вязкость глазной ткани. [12]
Вращательный момент инерции глаза незначителен, поскольку люди, носящие утяжеленные контактные линзы, которые увеличивают вращательный момент инерции почти в 100 раз, по-прежнему имеют тот же VOR (стр. 94 [13] ).
Вестибулоокулярный рефлекс должен быть быстрым: для ясного зрения движение головы должно компенсироваться почти немедленно; в противном случае зрение соответствует фотографии, сделанной дрожащей рукой. Сигналы посылаются из полукружных каналов с использованием всего трех нейронов, называемых трехнейронной дугой . Это приводит к движениям глаз, которые отстают от движения головы менее чем на 10 мс. [14] Вестибулоокулярный рефлекс является одним из самых быстрых рефлексов в организме человека.
Когда человек отслеживает движение чего-либо глазами и головой одновременно, VOR является контрпродуктивным для цели поддержания угла взгляда и головы на одном уровне. Исследования показывают, что в мозге существуют механизмы подавления VOR с использованием активной визуальной (ретинарной) обратной связи, получаемой при наблюдении за объектом в движении. [15] При отсутствии визуальной обратной связи, например, когда объект проходит за непрозрачным барьером, люди могут продолжать визуально отслеживать видимое положение объекта с помощью упреждающих (экстраретинальных) систем в мозге, и VOR также подавляется во время этой деятельности. VOR может быть даже когнитивно подавлен, например, при следовании за воображаемой целью с глазами и головой одновременно, хотя эффект, как правило, менее драматичен, чем при визуальной обратной связи. [16]
«Усиление» VOR определяется как изменение угла глаза, деленное на изменение угла головы во время поворота головы. В идеале усиление вращательного VOR равно 1,0. Усиление горизонтального и вертикального VOR обычно близко к 1,0, но усиление крутильного VOR (вращение вокруг линии визирования) обычно низкое. [4] Усиление трансляционного VOR должно быть скорректировано в зависимости от расстояния из-за геометрии параллакса движения. Когда голова транслируется, угловое направление ближних целей изменяется быстрее, чем угловое направление дальних целей. [6]
Если усиление VOR неправильное (отличается от 1) — например, если глазные мышцы слабые или если человек надевает новую пару очков — то движение головы приводит к движению изображения на сетчатке, что приводит к нечеткости зрения. В таких условиях двигательное обучение корректирует усиление VOR для получения более точного движения глаз. Это то, что называется адаптацией VOR.
Близорукие люди, которые обычно носят отрицательные очки, имеют более низкое усиление VOR. Дальнозоркие люди или афаки , которые обычно носят положительные очки, имеют более высокое усиление VOR. Люди, которые обычно носят контактные линзы, не показывают никаких изменений в усилении VOR. Монокулярная, неконъюгированная адаптация VOR возможна, например, после паралича экстраокулярных мышц . (стр. 27 [17] )
Фаза VOR также может адаптироваться. [18]
Окуломоторный интегратор является интегратором с утечкой, с характерным временем утечки ~20 с. Если время утечки слишком мало, происходит некоторая форма адаптации, чтобы «залатать утечку», чтобы увеличить время утечки. Предполагается, что интегратор с утечкой построен с помощью контура обратной связи с усилением немного ниже 1, а адаптация происходит путем регулировки усиления контура обратной связи. Гипотеза проверяется с помощью специально смоделированного оптокинетического барабана , который имитирует визуальный эффект наличия очень утечки окуломоторного интегратора. После 1 часа просмотра интегратор становится «анти-утечкой», что означает, что его значение растет экспоненциально даже при отсутствии входа. Движение глаза становится положительной обратной связью, что означает, что если он находится немного левее цели фиксации, он будет дрейфовать еще дальше влево, и аналогично для правого. Это также сопровождается тошнотой. [19] (стр. 84 [13] )

Потребление этанола может нарушить VOR, снижая динамическую остроту зрения. [20] В нормальных условиях купула и эндолимфа имеют одинаковую плотность (оба равны). После приема этанола этанол диффундирует в купулу, прежде чем диффундировать в эндолимфу, поскольку он находится ближе к кровеносным капиллярам. Это временно делает купулу легче. В этом состоянии, если человек ложится, касаясь правой щекой земли, то купула в левом ухе будет плавать влево, создавая иллюзорное ощущение медленного вращения головы слева направо. Чтобы компенсировать это, VOR медленно перемещает глаза влево, пока не достигнет предела, и глаза быстро тянутся вправо (нистагм). Это позиционный алкогольный нистагм , фаза I (PAN I). Необычная вестибулярная стимуляция также вызвала симптомы укачивания: иллюзии вращения тела, головокружение и тошноту. Эти симптомы исчезают через несколько секунд после принятия вертикального положения. [21]
Через некоторое время плотность купулы и эндолимфы выравнивается, устраняя эффект нистагма. После того, как этанол полностью метаболизируется, купула возвращается к нормальной плотности первой, создавая нистагм в противоположном направлении (PAN II) во время похмелья . [21]
Как и предполагалось, потребление тяжелой воды (плотность воды 1,1) имеет совершенно противоположный эффект нистагма по сравнению с потреблением этанола. Потребление смеси тяжелой воды ( ) и этанола ( ) в значительной степени отменяет эффект. [21] Макроглобулинемия , или потребление глицерина (плотность воды 1,26), имеет такие же эффекты, как и тяжелая вода. [22] [23] [24]
Этот рефлекс можно проверить с помощью теста быстрого импульса головы или теста Хальмаги-Кертойса , в котором голова быстро с силой перемещается в сторону, и контролируется, если глазам удается оставаться смотрящими в том же направлении. Когда функция правой системы равновесия снижается из-за болезни или несчастного случая, быстрое движение головы вправо больше не может быть воспринято должным образом. В результате не генерируется компенсаторное движение глаз, и пациент не может фиксировать точку в пространстве во время этого быстрого движения головы.
Тест на импульс головы можно проводить у постели больного и использовать в качестве скринингового инструмента для выявления проблем с вестибулярной системой человека. [25] Его также можно диагностически проверить, проведя видеотест на импульс головы (VHIT). В этом диагностическом тесте человек надевает высокочувствительные очки, которые обнаруживают быстрые изменения в движении глаз. Этот тест может предоставить специфическую для участка информацию о вестибулярной системе и ее функции. [26]
Другим способом проверки реакции VOR является тест на калорический рефлекс , который представляет собой попытку вызвать нистагм (компенсаторное движение глаз при отсутствии движения головы) путем заливки холодной или теплой воды в ухо. Также доступны битермические воздушные калорические орошения, при которых в ухо вводится теплый и холодный воздух. [27]
Вестибулоокулярный рефлекс можно проверить с помощью вышеупомянутого теста на калорический рефлекс ; это играет важную роль в подтверждении диагноза смерти ствола мозга. В этом процессе необходимо следовать кодексу практики, а именно кодексу Академии медицинских королевских колледжей. [28]
Резюме: Цервикоокулярный рефлекс, также известный под аббревиатурой COR, включает достижение стабилизации визуальной цели [29] и изображения на сетчатке посредством корректировки взгляда, вызванного движениями или поворотами шеи и/или головы. Процесс работает совместно с вестибулоокулярным рефлексом (VOR). [30] Он заметен у некоторых животных, которые не могут много двигать глазами, например, у сов. [31]






