Ceratosaurus / ˌ s ɛr ə t oʊ ˈ s ɔːr ə s / (от греч. κέρας/κέρατος, keras/keratos означает «рог» и σαῦρος sauros означает «ящерица») был плотоядным тероподовым динозавром , который жил в позднеюрский период ( кимериджский — титонский века ). Род был впервые описан в 1884 году американским палеонтологом Отниелом Чарльзом Маршем на основе почти полного скелета, обнаруженного в Гарден-Парке, штат Колорадо , в породах, принадлежащих формации Моррисон . Типовой вид — Ceratosaurus nasicornis .
Образец из Гарден-Парка остается наиболее полным скелетом, известным из рода, и с тех пор было описано лишь несколько дополнительных образцов. Два дополнительных вида, Ceratosaurus dentisulcatus и Ceratosaurus magnicornis , были описаны в 2000 году по двум фрагментарным скелетам из карьера Кливленд-Ллойд в Юте и из окрестностей Фруты, Колорадо . Однако обоснованность этих дополнительных видов была поставлена под сомнение, и все три скелета, возможно, представляют собой разные стадии роста одного и того же вида. В 1999 году было сообщено об открытии первого молодого образца. В 2000 году частичный образец был раскопан и описан из формации Лориньян в Португалии , что предоставило доказательства присутствия рода за пределами Северной Америки. Фрагментарные останки также были зарегистрированы в Танзании , Уругвае и Швейцарии , хотя их отнесение к Ceratosaurus в настоящее время не принимается большинством палеонтологов .
Ceratosaurus был тероподом среднего размера. Первоначальный образец оценивается в 5,3 м (17 футов) или 5,69 м (18,7 фута) в длину, в то время как образец, описанный как C. dentisulcatus, был больше, около 7 м (23 фута) в длину. Ceratosaurus характеризовался глубокими челюстями, которые поддерживали пропорционально очень длинные, похожие на лезвия зубы, выступающим, похожим на гребень рогом на средней линии морды и парой рогов над глазами. Передние конечности были очень короткими, но оставались полностью функциональными. Рука имела четыре пальца с когтями на первых трех. Хвост был глубоким сверху донизу. Ряд небольших остеодерм (кожных костей) присутствовал посередине шеи, спины и хвоста. Дополнительные остеодермы присутствовали в неизвестных местах на теле животного.
Ceratosaurus дал свое название Ceratosauria , кладе динозавров-теропод, которая рано отделилась от эволюционной линии, ведущей к современным птицам . Некоторые палеонтологи предположили, что внутри Ceratosauria он наиболее тесно связан с Genyodectes из Аргентины, который разделяет сильно удлиненные зубы. Геологически более старый род Proceratosaurus из Англии , хотя первоначально был описан как предполагаемый предшественник Ceratosaurus , позже был обнаружен как ранний тираннозавроид. Ceratosaurus делил свою среду обитания с другими крупными родами теропод, включая Torvosaurus и Allosaurus , и было высказано предположение, что эти тероподы занимали разные экологические ниши, чтобы уменьшить конкуренцию. Ceratosaurus мог охотиться на травоядных динозавров, хотя некоторые палеонтологи предполагали, что он охотился на водную добычу, такую как рыба. Носовой рог, вероятно, не использовался в качестве оружия, как первоначально предполагал Марш, а, скорее всего, использовался исключительно для демонстрации.
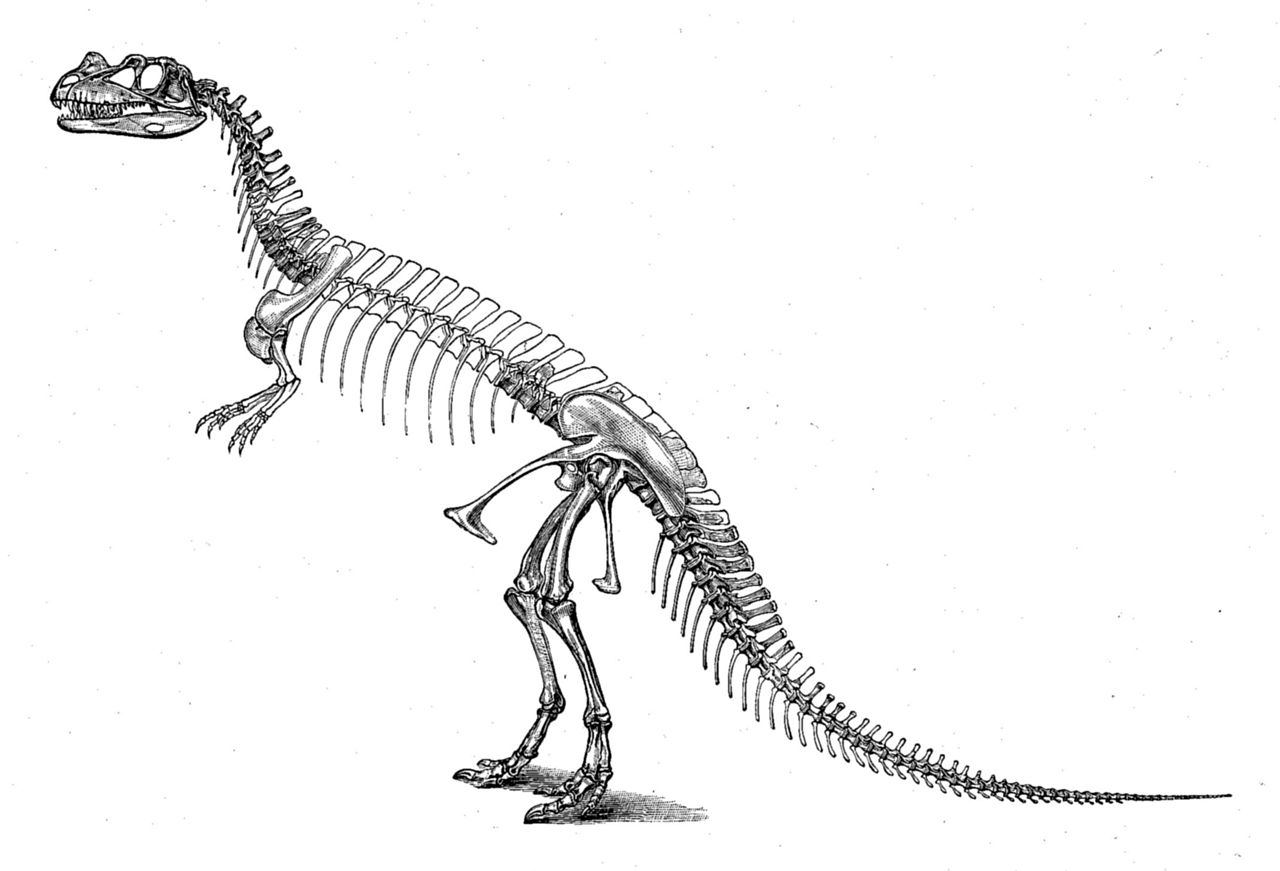
Первый образец, голотип USNM 4735, был обнаружен и выкопан фермером Маршаллом Паркером Фелчем в 1883 и 1884 годах. [1] Найденный в сочленении, с костями, все еще соединенными друг с другом, он был почти полным, включая череп. Значительные недостающие части включают неизвестное количество позвонков, все, кроме последних ребер туловища, плечевые кости (верхние кости руки), дистальные кости пальцев обеих рук, большую часть правой руки, большую часть левой ноги и большую часть ступней. [2] : 77 Образец был найден заключенным в твердый песчаник, что привело к тому, что череп и позвоночник были сильно деформированы во время окаменения . [2] : 2, 114 Место обнаружения, расположенное в районе Гарден-Парк к северу от Каньон-Сити, штат Колорадо , и известное как карьер Фелча 1, считается одним из самых богатых ископаемых мест формации Моррисон . Многочисленные окаменелости динозавров были извлечены из этого карьера еще до открытия цератозавра , среди которых наиболее примечательным был почти полный образец аллозавра (USNM 4734) в 1883 и 1884 годах. [2] : 7, 114
После раскопок образец был отправлен в Музей естественной истории Пибоди в Нью-Хейвене , где его изучал Марш, который в 1884 году описал его как новый род и вид Ceratosaurus nasicornis . [3] [2] : 114 Название Ceratosaurus можно перевести как «рогатый ящер» (от греческих слов κερας/κερατος , keras/keratos — «рог» и σαυρος / sauros — «ящерица») [4] и nasicornis с «носовым рогом» (от латинских слов nasus — «нос» и cornu — «рог»). [5] Учитывая полноту образца, недавно описанный род был на тот момент самым известным тероподом, обнаруженным в Америке. В 1898 и 1899 годах образец был передан в Национальный музей естественной истории в Вашингтоне, округ Колумбия , вместе со многими другими окаменелостями, первоначально описанными Маршем. Только часть этого материала была полностью подготовлена, когда он прибыл в Вашингтон. Последующая подготовка продолжалась с 1911 года до конца 1918 года. Упаковка и отправка из Нью-Хейвена в Вашингтон нанесли некоторые повреждения образцу цератозавра . [2] : 2, 114 В 1920 году Чарльз Гилмор опубликовал обширное переописание этого и других образцов теропод, полученных из Нью-Хейвена, включая почти полный образец аллозавра , извлеченный из того же карьера. [2] : 2

В статье 1892 года Марш опубликовал первую реконструкцию скелета цератозавра , которая изображает животное длиной 22 фута (6,7 м) и высотой 12 футов (3,7 м). [6] Как отметил Гилмор в 1920 году, туловище было изображено слишком длинным в этой реконструкции, включая по крайней мере шесть спинных позвонков больше, чем нужно. Эта ошибка была повторена в нескольких последующих публикациях, включая первую реконструкцию жизни, которая была нарисована в 1899 году Фрэнком Бондом под руководством Чарльза Р. Найта , но не опубликована до 1920 года. Более точная реконструкция жизни, опубликованная в 1901 году, была сделана Джозефом М. Глисоном , снова под руководством Найта. Голотип был смонтирован Гилмором в 1910 и 1911 годах. С тех пор он выставлялся в Национальном музее естественной истории. Большинство ранних реконструкций показывают цератозавра в вертикальном положении, с волочащимся по земле хвостом. [2] : 115–116 Напротив, крепление голотипа Гилмором значительно опередило свое время. [7] : 276 Вдохновленный верхними бедренными костями, которые были найдены под углом к голени, он изобразил крепление как бегущее животное с горизонтальной осанкой и хвостом, который не касался земли. Из-за сильного уплощения окаменелостей Гилмор установил образец не как отдельно стоящий скелет, а как барельеф внутри искусственной стены. [2] : 114 Поскольку кости были частично вмонтированы в пластину, научный доступ был ограничен. В ходе реконструкции экспозиции динозавров музея в период с 2014 по 2019 год образец был разобран и освобожден от закрывающей пластины. [8] [9] В новой выставке, открытие которой было запланировано на 2019 год, планировалось заменить монтировку отдельно стоящей моделью, а оригинальные кости должны были храниться в музейной коллекции, чтобы обеспечить полный доступ к ним для ученых. [9]
.jpg/1280px-Osteology_of_the_carnivorous_Dinosauria_in_the_United_States_National_museum_BHL40623218_edited_(cropped).jpg)
После открытия голотипа C. nasicornis , значительная находка Ceratosaurus была сделана только в начале 1960-х годов, когда палеонтолог Джеймс Мэдсен и его команда раскопали фрагментарный, разрозненный скелет, включая череп (UMNH VP 5278) в карьере динозавров Кливленд-Ллойд в штате Юта. Эта находка представляет собой один из крупнейших известных образцов Ceratosaurus . [10] : 21 Второй, сочлененный образец, включая череп (MWC 1), был обнаружен в 1976 году Тором Эриксоном, сыном палеонтолога Лэнса Эриксона, недалеко от Фруты, штат Колорадо . [4] Довольно полный образец, у него отсутствуют нижние челюсти, предплечья и брюшные полости . Череп, хотя и достаточно полный, был найден разрозненным и сильно сплющенным с боков. Хотя это была крупная особь, она еще не достигла размеров взрослой особи, на что указывают несросшиеся швы между костями черепа. [10] : 2–3 Научно точные трехмерные реконструкции черепа для использования в музейных экспонатах были созданы с использованием сложного процесса, включающего формовку и литье отдельных оригинальных костей, исправление деформаций, реконструкцию недостающих частей, сборку слепков костей в их правильном положении и покраску в соответствии с оригинальным цветом костей. [11]
Оба образца из Фруты и Кливленд-Ллойда были описаны Мэдсеном и Сэмюэлем Полом Уэллсом в монографии 2000 года, причем образец из Юты был отнесен к новому виду C. dentisulcatus , а образец из Колорадо — к новому виду C. magnicornis . [10] Название dentisulcatus относится к параллельным бороздкам , присутствующим на внутренних сторонах предчелюстных зубов и первых трех зубах нижней челюсти у этого образца. Magnicornis указывает на более крупный носовой рог. [10] : 2, 21 Однако валидность обоих видов была поставлена под сомнение в последующих публикациях. Брукс Бритт и коллеги в 2000 году заявили, что голотип C. nasicornis на самом деле был молодой особью, а два более крупных вида представляли взрослое состояние одного вида. [12] Оливер Раухут в 2003 году, а также Мэтью Каррано и Скотт Сэмпсон в 2008 году рассмотрели анатомические различия, на которые ссылались Мэдсен и Уэллс, в поддержку того, что эти дополнительные виды представляют онтогенетические (возрастные) или индивидуальные изменения. [13] [14] : 192
Еще один образец (BYUVP 12893) был обнаружен в 1992 году в карьере Агат-Бейсин к юго-востоку от Мура, штат Юта , но все еще ожидает описания. Образец, считающийся самым крупным из известных представителей рода, включает переднюю половину черепа, семь фрагментарных тазовых спинных позвонков и сочлененный таз и крестец. [14] : 192 [10] : 36 В 1999 году Бритт сообщил об обнаружении скелета цератозавра, принадлежащего молодой особи. Обнаруженный в карьере Боун-Кэбин в Вайоминге, он на 34% меньше голотипа C. nasicornis и состоит из полного черепа, а также 30% остальной части скелета, включая полный таз. [15]
Помимо этих пяти скелетных находок, фрагментарные останки Ceratosaurus были обнаружены в различных местах из стратиграфических зон 2 и 4-6 формации Моррисон, [16] включая некоторые из основных ископаемых мест формации. Национальный памятник динозавров , штат Юта, дал изолированную правую предчелюстную кость (DNM 972). Большая лопатка (скапулокоракоид) была обнаружена в Комо-Блафф в Вайоминге . Другой образец происходит из карьера Драй-Меса в Колорадо и включает левый скапулокоракоид, а также фрагменты позвонков и костей конечностей. В карьере Майгатт-Мур, штат Колорадо, род известен по зубам. [10] : 36
С 1909 по 1913 год немецкие экспедиции Берлинского музея естествознания обнаружили разнообразную фауну динозавров в формации Тендагуру в Германской Восточной Африке , на территории современной Танзании . [17] Хотя обычно считается самым важным местонахождением динозавров в Африке, [17] крупные динозавры-тероподы известны только по немногим и очень фрагментарным останкам. [18] В 1920 году немецкий палеонтолог Вернер Яненш отнес несколько спинных позвонков из карьера «TL» к Ceratosaurus , как Ceratosaurus sp. (неопределенного вида). В 1925 году Яненш назвал новый вид Ceratosaurus , C. roechlingi , на основе фрагментарных останков из карьера «Mw», включающих квадратную кость, малоберцовую кость, фрагментарные хвостовые позвонки и другие фрагменты. Этот образец происходит от особи, значительно более крупной, чем голотип C. nasicornis . [18]

В своей монографии 2000 года Мадсен и Уэллс подтвердили отнесение этих находок к Ceratosaurus . Кроме того, они отнесли несколько зубов к роду, который первоначально был описан Яненшем как возможный вид Labrosaurus , Labrosaurus (?) stechowi . [10] Другие авторы подвергли сомнению отнесение любой из находок из Тендагуру к Ceratosaurus , отметив, что ни один из этих образцов не демонстрирует признаков, диагностических для этого рода. [19] [20] : 66 [14] : 192 [21] В 2011 году Раухут обнаружил, что и C. roechlingi , и Labrosaurus (?) stechowi являются возможными цератозаврами, но обнаружил, что они недиагностичны на уровне рода, и обозначил их как nomina dubia (сомнительные названия). [21] В 1990 году Тимоти Роу и Жак Готье упомянули еще один вид Ceratosaurus из Тендагуру, Ceratosaurus ingens , который предположительно был установлен Яненшем в 1920 году и был основан на 25 изолированных, очень больших зубах длиной до 15 см (5,9 дюйма). [19] [18] Однако Яненш отнес этот вид к Megalosaurus , а не к Ceratosaurus . Поэтому это название может быть простой ошибкой копирования. [10] : 37 [18] Раухут в 2011 году показал, что Megalosaurus ingens не был тесно связан ни с Megalosaurus , ни с Ceratosaurus , но, возможно, представляет собой кархародонтозаврида . [21]
В 2000 и 2006 годах палеонтологи под руководством Октавио Матеуса описали находку из формации Лориньян в центрально-западной Португалии (ML 352) как новый образец цератозавра , состоящий из правой бедренной кости (верхняя бедренная кость), левой большеберцовой кости (большая берцовая кость) и нескольких отдельных зубов, извлеченных из скал пляжа Вальмитао, между муниципалитетами Лориньян и Торриш-Ведраш . [22] [23] Кости были обнаружены в мелкозернистых песчаниках желто-коричневого цвета, которые были отложены реками в качестве пойменных отложений и относятся к нижним уровням группы Порту-Ново, которая, как полагают, относится к позднему кимериджскому ярусу . Дополнительные кости этой особи (SHN (JJS)-65), включая левую бедренную кость, правую большеберцовую кость и частичную левую малоберцовую кость (кость голени), с тех пор были обнажены из-за прогрессирующей эрозии скал . Хотя изначально они были частью частной коллекции, эти дополнительные элементы стали официально кураторскими после того, как частная коллекция была передана в дар Обществу естественной истории в Торриш-Ведраш и была подробно описана в 2015 году. [24] Образец был отнесен к виду Ceratosaurus dentisulcatus Матеусом и коллегами в 2006 году. [23] Обзор 2008 года, проведенный Каррано и Сэмпсоном, подтвердил отнесение к виду Ceratosaurus , но пришел к выводу, что отнесение к какому-либо конкретному виду в настоящее время невозможно. [14] : 192 В 2015 году Элизабет Малафая и коллеги, которые усомнились в обоснованности C. dentisulcatus , отнесли образец к виду Ceratosaurus aff. Ceratosaurus nasicornis . [24]
Другие сообщения включают один зуб, найденный в Мутье , Швейцария . Первоначально названный Яненшем в 1920 году как Labrosaurus meriani , зуб позже был отнесен к Ceratosaurus sp. (неизвестного вида) Мадсеном и Уэллсом. [10] : 35–36 В 2008 году Матиас Сото и Даниэль Переа описали зубы из формации Такуарембо в Уругвае , включая предполагаемую коронку предчелюстного зуба. Он показывает вертикальные полосы на своей внутренней стороне и не имеет зубчиков на переднем крае. Эти особенности в этой комбинации известны только для Ceratosaurus . Авторы, однако, подчеркнули, что отнесение к Ceratosaurus нецелесообразно, поскольку останки скудны, и отмечают, что отнесение европейского и африканского материала к Ceratosaurus следует рассматривать с осторожностью. [25] В 2020 году Сото и его коллеги описали дополнительные зубы цератозавра из той же формации, которые еще раз подтверждают их более раннюю интерпретацию. [26]

Ceratosaurus следовал плану тела , типичному для крупных тероподовых динозавров. [6] Будучи двуногим , он передвигался на мощных ногах, в то время как его руки были уменьшены в размере. Образец USNM 4735, первый обнаруженный скелет и голотип Ceratosaurus nasicornis , был особью длиной 5,3 м (17 футов) или 5,69 м (18,7 фута) согласно отдельным источникам. [2] : 115 [7] Было ли это животное полностью взрослым, неясно. [20] : 66 Отниэль Чарльз Марш в 1884 году предположил, что этот образец весил примерно вдвое меньше, чем современный ему аллозавр . [3] В более поздних отчетах это было пересмотрено до 418 килограммов (922 фунта), 524 кг (1155 фунтов) или 670 кг (1480 фунтов). [27] Три дополнительных скелета, обнаруженных во второй половине 20-го века, были существенно больше. Первый из них, UMNH VP 5278, был оценен Джеймсом Мэдсеном как около 8,8 м (29 футов) в длину, [4] но позже был оценен в 7 м (23 фута) в длину. [28] Его вес был рассчитан как 980 кг (2160 фунтов), 452 кг (996 фунтов) и 700 кг (1540 фунтов) в отдельных работах. [7] [28] [29] Второй скелет, MWC 1, был несколько меньше, чем UMNH VP 5278, и мог весить 275 кг (606 фунтов). [29] Третий, пока неописанный, образец BYUVP 12893 был заявлен как самый большой из обнаруженных, хотя оценки не были опубликованы. [14] : 192 Другой экземпляр, ML 352, обнаруженный в Португалии в 2000 году, был оценен в 6 м (20 футов) в длину и 600 кг (1320 фунтов). [28]

Череп был довольно большим по сравнению с остальной частью тела. [6] Длина у голотипа C. nasicornis составляла 55 см (22 дюйма) , измеренная от кончика морды до затылочного мыщелка , который соединяется с первым шейным позвонком. [2] : 88 Ширину этого черепа трудно реконструировать, так как он сильно искажен, а реконструкция Гилмора 1920 года позже оказалась слишком широкой. [30] Довольно полный череп образца MWC 1, по оценкам, имел длину 60 см (24 дюйма) и ширину 16 см (6,3 дюйма). Этот череп был несколько более вытянутым, чем у голотипа. [10] : 3 Задняя часть черепа была более легко сложена, чем у некоторых других крупных теропод из-за обширных черепных отверстий, однако челюсти были глубокими, чтобы поддерживать пропорционально большие зубы. [7] : 277 Слезная кость образовывала не только задний край анторбитального окна , большого отверстия между глазом и костной ноздрей , но также часть его верхнего края, в отличие от членов родственных Abelisauridae . Квадратная кость , которая была соединена с нижней челюстью на ее нижнем конце, чтобы сформировать челюстной сустав, была наклонена так, что челюстной сустав был смещен назад по отношению к затылочному мыщелку. Это также привело к расширению основания латерального височного окна , большого отверстия позади глаз. [20] : 53
Самой отличительной чертой был выступающий рог, расположенный на средней линии черепа за костными ноздрями, который был образован из сросшихся выступов левой и правой носовых костей . [2] : 82 Из ископаемых известен только костный роговой стержень. У живого животного этот стержень поддерживал кератиновую оболочку. В то время как основание рогового стержня было гладким, его верхние две трети были морщинистыми и выстланы бороздками, которые содержали кровеносные сосуды при жизни. У голотипа роговой стержень имеет длину 13 см (5,1 дюйма) и ширину 2 см (0,79 дюйма) у основания, но быстро сужается до всего 1,2 см (0,47 дюйма) выше и имеет высоту 7 см (2,8 дюйма). [2] : 82 Он длиннее и ниже в черепе MWC 1. [10] : 3 У современного животного рог, вероятно, был бы более удлиненным из-за его кератиновой оболочки. [31] За носовым рогом носовые кости образовали овальную бороздку. И эта бороздка, и носовой рог служат признаками, отличающими Ceratosaurus от родственных родов. [14] : 192 В дополнение к большому носовому рогу, Ceratosaurus обладал меньшими, полукруглыми, костными гребнями перед каждым глазом, похожими на таковые у Allosaurus . Эти гребни были образованы слезными костями. [29] У молодых особей все три рога были меньше, чем у взрослых, и две половины ядра носового рога еще не срослись. [15]
_-_AMNH_-_DSC06292.JPG/1280px-Ceratosaurus_nasicornis_(cast)_-_AMNH_-_DSC06292.JPG)
Предчелюстные кости , которые образовывали кончик морды, содержали всего по три зуба с каждой стороны, меньше, чем у большинства других теропод. [20] : 52 Верхнечелюстные кости верхней челюсти были выложены 15 лезвиевидными зубами с каждой стороны у голотипа. Первые восемь из этих зубов были очень длинными и крепкими, но начиная с девятого зуба они постепенно уменьшаются в размере. Как типично для теропод, они имели мелко зазубренные края, которые содержали около 10 зубцов на 5 мм (0,20 дюйма) у голотипа. [2] : 92 Образец MWC 1 показал всего 11-12, а образец UMNH VP 5278 показал 12 зубов на каждой верхней челюсти. У последнего образца зубы были более крепкими и более изогнутыми. [10] : 3, 27 У всех образцов коронки зубов верхних челюстей были исключительно длинными. У образца UMNH VP 5278 они достигали 9,3 см (3,7 дюйма) в длину, что равно минимальной высоте нижней челюсти. У голотипа они составляют 7 см (2,8 дюйма) в длину, что даже превышает минимальную высоту нижней челюсти. У других теропод сопоставимая длина зубов известна только у возможно близкородственного Genyodectes . [32] Напротив, у нескольких членов Abelisauridae коронки зубов очень короткие. [20] : 92 У голотипа каждая половина зубной кости , зубонесущей кости нижней челюсти , была оснащена 15 зубами, которые, однако, плохо сохранились. Оба образца MWC 1 и UMNH VP 5278 показывают только 11 зубов в каждой зубной кости, которые, как показывает последний образец, были немного прямее и менее прочными, чем зубы верхней челюсти. [10] : 3, 21

Точное количество позвонков неизвестно из-за нескольких щелей в позвоночнике голотипа Ceratosaurus nasicornis . По крайней мере 20 позвонков образовывали шею и спину перед крестцом . В средней части шеи тела позвонков были такой же длины, как и их высота, в то время как в передней и задней частях шеи тела были короче своей высоты. Выступающие вверх остистые отростки были сравнительно большими, а в спинных позвонках были такими же высокими, как и позвоночные тела. Крестец, состоящий из шести сросшихся крестцовых позвонков , был выгнут вверх, а его позвоночные тела были сильно уменьшены по высоте в средней части, как это бывает у некоторых других цератозавров . [20] : 55–58 Хвост состоял примерно из 50 хвостовых позвонков и составлял около половины общей длины животного. В голотипе он был оценен в 2,84 м (9,33 фута). [3] [2] : 115 Хвост был глубоким сверху донизу из-за высоких остистого отростка и удлиненных шевронов , костей, расположенных ниже тел позвонков. Как и у других динозавров, он уравновешивал тело и содержал массивную каудофеморальную мышцу, которая отвечала за толчок вперед во время передвижения, тянущую верхнюю часть бедра назад при сокращении . [20] : 55–58

Лопатка (плечевая кость) была сращена с коракоидом , образуя единую кость без видимой границы между двумя исходными элементами. [ 20] : 58 Голотип C. nasicornis был найден с сочлененной левой рукой, включая неполную кисть. Хотя во время подготовки она была разъединена , заранее был сделан слепок ископаемого, чтобы задокументировать первоначальное относительное положение костей. Кости запястья не были известны ни у одного образца, что заставило некоторых авторов предположить, что они были утрачены в роде. В статье 2016 года Мэтью Каррано и Джона Шуаньер предположили, что, вероятно, присутствовали одна или несколько хрящевых (не костных) костей запястья, на что указывает зазор между костями предплечья и пястными костями, а также текстура поверхности внутри этого зазора, видимая в слепке. [33] В отличие от большинства более продвинутых теропод, у которых было только по три пальца на каждой руке (пальцы I–III), у Ceratosaurus сохранилось четыре пальца, а палец IV был уменьшен в размере. Первая и четвертая пястные кости были короткими, а вторая была немного длиннее третьей. Пястная кость и особенно первые фаланги были пропорционально очень короткими, в отличие от большинства других базальных теропод. У голотипа сохранились только первые фаланги пальцев II, III и IV. Общее количество фаланг и когтей (костей когтей) неизвестно. Анатомия пястной кости I указывает на то, что фаланги изначально присутствовали и на этом пальце. Стопа (ped) состояла из трех опорных пальцев, пронумерованных от II до IV. Палец I, который у теропод обычно редуцирован до прибылого когтя , не касающегося земли, у голотипа не сохранился. Марш в своем первоначальном описании 1884 года предположил, что этот палец был утрачен у цератозавра , но Чарльз Гилмор в своей монографии 1920 года отметил область прикрепления на второй плюсневой кости , демонстрирующую наличие этого пальца. [2] : 112
Уникальный среди теропод, Ceratosaurus обладал небольшими, удлиненными и неправильной формы остеодермами (кожными костями) вдоль средней линии своего тела. Такие остеодермы были обнаружены над остистыми отростками шейных позвонков 4 и 5, а также над хвостовыми позвонками 4–10, и, вероятно, образовывали непрерывный ряд, который мог простираться от основания черепа до большей части хвоста. Как предположил Гилмор в 1920 году, их положение в каменном матриксе, вероятно, отражает их точное положение в живом животном. Остеодермы над хвостом были обнаружены отделенными от остистого отростка на 25 мм (0,98 дюйма)–38 мм (1,5 дюйма), что, возможно, объясняется наличием кожи и мышц между ними, в то время как остеодермы шеи были гораздо ближе к остистым отросткам. Помимо средней линии тела, кожа содержала дополнительные остеодермы, на что указывает большая, примерно четырехугольная пластина размером 58 мм (2,3 дюйма) на 70 мм (2,8 дюйма), найденная вместе с голотипом. Однако положение этой пластины на теле неизвестно. [2] : 113–114 Образец UMNH VP 5278 также был обнаружен с рядом остеодерм, которые были описаны как аморфные по форме. Хотя большинство этих косточек были найдены на расстоянии не более 5 м от скелета, они не были напрямую связаны с какими-либо позвонками, в отличие от голотипа C. nasicornis , поэтому их первоначальное положение на теле не может быть выведено из этого образца. [10] : 32

В своем первоначальном описании голотипа Ceratosaurus nasicornis и последующих публикациях Марш отметил ряд характеристик , которые были неизвестны всем другим тероподам, известным в то время. [14] : 185 Две из этих особенностей, сросшийся таз и сросшаяся плюсна, были известны у современных птиц и, по словам Марша, ясно демонстрируют тесную связь между последними и динозаврами. [34] Чтобы отделить род от Allosaurus , Megalosaurus и целурозавров , Марш сделал Ceratosaurus единственным членом как нового семейства , Ceratosauridae , так и нового инфраотряда , Ceratosauria. [14] : 185 Это было подвергнуто сомнению в 1892 году Эдвардом Дринкером Коупом , главным соперником Марша в Войнах костей , который утверждал, что отличительные черты, такие как носовой рог, просто показывают, что C. nasicornis был отдельным видом, но недостаточны для обоснования отдельного рода. Следовательно, он отнес C. nasicornis к роду Megalosaurus , создав новую комбинацию Megalosaurus nasicornis . [35]
Хотя Ceratosaurus сохранялся как отдельный род во всех последующих анализах, [2] : 76 его родственные связи оставались спорными в течение следующего столетия. Как Ceratosauridae, так и Ceratosauria не были широко приняты, идентифицировано было лишь несколько и малоизвестных дополнительных членов. На протяжении многих лет отдельные авторы классифицировали Ceratosaurus в пределах Deinodontidae , Megalosauridae , Coelurosauria , Carnosauria и Deinodontoidea . [10] : 2 В своей ревизии 1920 года Гилмор утверждал, что этот род был наиболее базальным тероподом, известным после триаса , и не был так тесно связан ни с одним другим современным тероподом, известным в то время. Таким образом, он заслуживает своего собственного семейства: Ceratosauridae. [2] : 76 Однако только после создания кладистического анализа в 1980-х годах первоначальное заявление Марша о Ceratosauria как об отдельной группе получило поддержку. В 1985 году было обнаружено , что недавно открытые южноамериканские роды Abelisaurus и Carnotaurus тесно связаны с Ceratosaurus . В 1986 году Готье признал Coelophysoidea тесно связанным с Ceratosaurus , хотя эта клада не входит в Ceratosauria в большинстве последних анализов. С тех пор было признано много дополнительных членов Ceratosauria. [14] : 185
Ceratosauria рано отделилась от эволюционной линии, ведущей к современным птицам, и считается базальным внутри теропод. [36] Сам Ceratosauria содержит группу производных (не базальных) членов семейств Noasauridae и Abelisauridae , которые заключены в скобки внутри клады Abelisauroidea , а также ряд базальных членов, таких как Elaphrosaurus , Deltadromeus и Ceratosaurus . Положение Ceratosaurus среди базальных цератозавров является предметом споров. Некоторые анализы рассматривали Ceratosaurus как наиболее производного из базальных членов, образуя сестринский таксон Abelisauroidea. [14] : 187 [37] Оливер Раухут в 2004 году предложил Genyodectes в качестве сестринского таксона Ceratosaurus , поскольку оба рода характеризуются исключительно длинными зубами в верхней челюсти. [32] Раухут объединил Ceratosaurus и Genyodectes в семейство Ceratosauridae, [32] за которым последовало несколько более поздних отчетов. [38] [39] [40] [31]
Шуо Ван и его коллеги в 2017 году пришли к выводу, что Noasauridae не были вложены в Abelisauroidea, как предполагалось ранее, а были более базальными, чем Ceratosaurus . Поскольку ноазавриды использовались в качестве фиксированной точки для определения клад Abelisauroidea и Abelisauridae, эти клады, следовательно, включали бы гораздо больше таксонов по определению, включая Ceratosaurus . В последующем исследовании 2018 года Рафаэль Делькур принял эти результаты, но указал, что, как следствие, Abelisauroidea необходимо будет заменить на старый синоним Ceratosauroidea, который до сих пор использовался редко. Для Abelisauridae Делькур предложил новое определение, которое исключает Ceratosaurus , что позволяет использовать название в его традиционном смысле. Ван и его коллеги также обнаружили, что Ceratosaurus и Genyodectes образуют кладу с аргентинским родом Eoabelisaurus . [40] Делькур использовал название Ceratosauridae для обозначения этой же клады и предложил определить Ceratosauridae как содержащую все таксоны, которые более тесно связаны с Ceratosaurus, чем с абелизавридом Carnotaurus . [31]

Следующая кладограмма, показывающая родственные связи цератозавра , основана на филогенетическом анализе, проведенном Диего Полом и Оливером Раухутом в 2012 году: [38]
Череп из средней юры Англии, по-видимому, демонстрирует носовой рог, похожий на рог Ceratosaurus . В 1926 году Фридрих фон Хюне описал этот череп как Proceratosaurus (что означает «до Ceratosaurus »), предполагая, что он был предшественником позднеюрского Ceratosaurus . [41] Сегодня Proceratosaurus считается базальным членом Tyrannosauroidea , гораздо более продвинутой клады тероподных динозавров. [42] Носовой рог мог бы развиться независимо у обоих родов. [14] : 185 Оливер Раухут и его коллеги в 2010 году сгруппировали Proceratosaurus в его собственное семейство, Proceratosauridae . Эти авторы также отметили, что носовой рог сохранился не полностью, что открывает возможность того, что он представлял собой переднюю часть более обширного гребня головы, как это наблюдается у некоторых других процератозавридов, таких как Гуаньлун . [42]
В формации Моррисон и Лориньян окаменелости цератозавра часто находят вместе с останками других крупных теропод, включая мегалозаврида Torvosaurus [43] и аллозаврида Allosaurus . Местонахождение Гарден-Парк в Колорадо содержало, помимо Ceratosaurus , окаменелости, приписываемые Allosaurus . Карьер Драй-Меса в Колорадо, а также карьер Кливленд-Ллойд и Национальный памятник динозаврам в Юте содержат останки по крайней мере трех крупных теропод: Ceratosaurus , Allosaurus и Torvosaurus . [30] [44] Аналогичным образом, Комо-Блафф и близлежащие местонахождения в Вайоминге содержали останки Ceratosaurus , Allosaurus и по крайней мере одного крупного мегалозаврида. [45] Цератозавр был редким представителем фауны теропод, так как его численность уступала аллозавру в среднем в 7,5 раза в местах их совместного обитания. [46]
Несколько исследований пытались объяснить, как эти симпатрические виды могли снизить прямую конкуренцию. Дональд Хендерсон в 1998 году утверждал, что Ceratosaurus сосуществовал с двумя отдельными потенциальными видами Allosaurus , которые он обозначил как «морфы»: морфа с укороченной мордой, высоким и широким черепом и короткими, выступающими назад зубами, и морфа, характеризующаяся более длинной мордой, низким черепом и длинными вертикальными зубами. Вообще говоря, чем больше сходства между симпатрическими видами в отношении их морфологии, физиологии и поведения, тем более интенсивной будет конкуренция между этими видами. Хендерсон пришел к выводу, что короткомордый морф Allosaurus занимал иную экологическую нишу , чем длинномордый морф и Ceratosaurus . Более короткий череп в этой морфе уменьшил бы изгибающие моменты, возникающие во время укуса, таким образом увеличивая силу укуса, сопоставимую с состоянием, наблюдаемым у кошек. Однако Ceratosaurus и другие морфы Allosaurus имели длинномордые черепа, которые лучше по сравнению с черепами собак. Более длинные зубы использовались бы как клыки для быстрых, режущих укусов, при этом сила укуса была бы сосредоточена на меньшей площади из-за более узкого черепа. По словам Хендерсона, большое сходство в форме черепа между Ceratosaurus и длинномордой морфой Allosaurus указывает на то, что эти формы напрямую конкурировали друг с другом. Поэтому Ceratosaurus мог быть вытеснен из мест обитания, где доминировал длинномордый морф. Действительно, Ceratosaurus очень редко встречается в карьере Кливленд-Ллойд, где обитает длинномордый морф Allosaurus , но, по-видимому, более распространен как в Гарден-Парке, так и в карьере Драй-Меса, где он встречается вместе с короткомордым морфом. [30]

Более того, Хендерсон предположил, что Ceratosaurus мог избегать конкуренции, предпочитая различные виды добычи. Эволюция его чрезвычайно удлиненных зубов могла быть прямым результатом конкуренции с длиннорылым морфом Allosaurus . Оба вида также могли предпочитать различные части туш, когда выступали в роли падальщиков. Удлиненные зубы Ceratosaurus могли служить визуальными сигналами, облегчающими распознавание членов того же вида, или для других социальных функций. Кроме того, большой размер этих теропод, как правило, снижал конкуренцию, поскольку количество возможных видов добычи увеличивается с размером. [30]
Фостер и Дэниел Чур в исследовании 2006 года согласились с Хендерсоном в том, что цератозавр и аллозавр в целом разделяли одни и те же места обитания и охотились на одни и те же типы добычи, что означает, что у них, вероятно, были разные стратегии питания, чтобы избежать конкуренции. По мнению этих исследователей, это также подтверждается разными пропорциями черепа, зубов и рук. [46] Однако различие между двумя морфами аллозавра было поставлено под сомнение некоторыми более поздними исследованиями. Кеннет Карпентер в исследовании 2010 года обнаружил, что короткомордые особи аллозавра из карьера Кливленд-Ллойд представляют собой случаи экстремальных индивидуальных вариаций, а не отдельный таксон. [ 47] Кроме того, череп USNM 4734 из местности Гарден-Парк, который лег в основу анализа Хендерсоном короткомордой морфы, позже оказался реконструированным слишком коротко. [48]
В исследовании 2004 года Роберт Баккер и Гэри Бир предположили, что цератозавр был в первую очередь специализирован на водной добыче, такой как двоякодышащие рыбы , крокодилы и черепахи. Как показал статистический анализ выпавших зубов из 50 отдельных мест в районе Комо-Блафф, зубы как цератозавров , так и мегалозавридов были наиболее распространены в местах обитания внутри и вокруг источников воды, таких как влажные поймы , берега озер и болота. Цератозавры также иногда встречались в наземных местообитаниях. Однако аллозавриды были одинаково распространены в наземных и водных местообитаниях. Из этих результатов Баккер и Бир пришли к выводу, что цератозавры и мегалозавриды, должно быть, в основном охотились вблизи и внутри водоемов, а цератозавры также иногда питались тушами более крупных динозавров. Исследователи также отметили длинное, низкое и гибкое тело цератозавра и мегалозаврид. По сравнению с другими тероподами Моррисона, цератозавр показал более высокие невральные отростки на передних хвостовых позвонках, которые были вертикальными, а не наклоненными к спине. Вместе с глубокими шевронными костями на нижней стороне хвоста они указывают на глубокий, «крокодилоподобный» хвост, возможно, приспособленный для плавания. Напротив, аллозавриды имеют более короткое, высокое и жесткое тело с более длинными ногами. Они были приспособлены к быстрому бегу по открытой местности и охоте на крупных травоядных динозавров, таких как зауроподы и стегозавры, но, как предполагают Баккер и Бир, сезонно переключались на водную добычу, когда крупные травоядные отсутствовали. [45] Однако в 2019 году эта теория была оспорена Юнем, который предположил, что цератозавр был просто более способен охотиться на водную добычу, чем другие тероподы формации Моррисон, а не был полностью полуводным. [49]

В своей популярной книге 1986 года «Ереси динозавров» Беккер утверждал, что кости верхней челюсти были лишь слабо прикреплены к окружающим костям черепа, что позволяло некоторую степень движения внутри черепа, состояние, называемое краниальным кинезисом . Аналогичным образом, кости нижней челюсти могли бы двигаться друг против друга, а квадратная кость могла бы качаться наружу, расширяя нижнюю челюсть в челюстном суставе. В совокупности эти особенности позволяли бы животному расширять челюсти, чтобы глотать более крупные пищевые продукты. [50] В исследовании 2008 года Кейси Холлидей и Лоуренс Уитмер переоценили аналогичные утверждения, сделанные в отношении других динозавров, заключив, что наличие мышечного краниального кинезиса не может быть доказано ни для одного вида динозавров и, вероятно, отсутствовало у большинства. [51]
На лобковой ступне аллозавра видны следы зубов другого теропода, вероятно, цератозавра или торвозавра . Расположение кости в теле (вдоль нижнего края туловища и частично прикрытая ногами) и тот факт, что она была одной из самых массивных в скелете, указывает на то, что аллозавр был добычей падальщиков. [ 52] Костный комплекс в верхнеюрском карьере Майгатт-Мур сохраняет необычно высокую частоту следов укусов теропод, большинство из которых можно отнести к аллозавру и цератозавру , в то время как другие могли быть сделаны заурофаганаксом или торвозавром, учитывая размер полос. В то время как положение следов укусов на травоядных динозаврах согласуется с хищничеством или ранним доступом к останкам, следы укусов, обнаруженные на материале аллозавра, предполагают добычу падальщиков либо от других теропод, либо от другого аллозавра . Необычно высокая концентрация следов укусов теропод по сравнению с другими сообществами может быть объяснена либо более полным использованием ресурсов тероподами в сухой сезон, либо предпочтителен сбор в других местах. [53]
В 1884 году Марш считал носовой рог цератозавра «самым мощным оружием» как для наступательных, так и для оборонительных целей, и Гилмор в 1920 году согласился с этой интерпретацией. [3] : 331 [2] : 82 Использование рога в качестве оружия в настоящее время в целом считается маловероятным. [4] В 1985 году Дэвид Норман считал, что рог «вероятно, не был предназначен для защиты от других хищников», а мог вместо этого использоваться для внутривидовых сражений между самцами цератозавров, боровшимися за права на размножение. [54] Грегори С. Пол в 1988 году предположил аналогичную функцию и проиллюстрировал двух цератозавров, участвующих в несмертельном бодунстве. [7] В 1990 году Роу и Готье пошли дальше, предположив, что носовой рог цератозавра «вероятно, использовался только в демонстрационных целях» и не играл никакой роли в физических столкновениях. [19] Если бы рог использовался для демонстрации, он, вероятно, был бы ярко окрашен. [29] Также предполагалось, что демонстрационная функция существует для ряда остеодерм, проходящих по средней линии тела. [19]

Сильно укороченные пястные кости и фаланги Ceratosaurus поднимают вопрос о том, сохранила ли рука хватательную функцию, предполагаемую для других базальных теропод. Внутри Ceratosauria еще более экстремальную редукцию руки можно наблюдать у абелизаврид, где рука утратила свою первоначальную функцию [ 55] и у Limusaurus . В статье 2016 года об анатомии руки Ceratosaurus Каррано и Джона Шуаньер подчеркнули большое морфологическое сходство руки с руками других базальных теропод, предположив, что она все еще выполняла свою первоначальную хватательную функцию, несмотря на свое укорочение. Хотя сохранились только первые фаланги, вторые фаланги были бы подвижными, на что указывают хорошо развитые суставные поверхности, и пальцы, вероятно, допускали бы такую же степень движения, как и у других базальных теропод. Как и у других теропод, кроме абелизаврид, первый палец был слегка повернут внутрь при сгибании . [33]

Под наблюдением Марша, вероятно, во время подготовки черепа, был сделан слепок мозговой полости голотипа, что позволило Маршу сделать вывод, что мозг «был среднего размера, но сравнительно намного больше, чем у травоядных динозавров». Однако кости черепа были впоследствии скреплены цементом, поэтому точность этого слепка не могла быть проверена более поздними исследованиями. [3] [2] : 93
Вторая, хорошо сохранившаяся мозговая коробка была найдена с образцом MWC 1 в Фруте, штат Колорадо, и была подвергнута компьютерной томографии палеонтологами Кентом Сандерсом и Дэвидом Смитом, что позволило реконструировать внутреннее ухо , крупные области мозга и черепные пазухи , транспортирующие кровь от мозга. В 2005 году исследователи пришли к выводу, что у Ceratosaurus была типичная для базальных теропод мозговая полость, похожая на таковую у Allosaurus . Отпечатки обонятельных луковиц , в которых находится обоняние, хорошо сохранились. Хотя они похожи на таковые у Allosaurus , они были меньше, чем у Tyrannosaurus , который, как полагают, был оснащен очень острым обонянием. Полукружные каналы , которые отвечают за чувство равновесия и, следовательно, позволяют делать выводы о привычной ориентации головы и передвижении, похожи на те, что встречаются у других теропод. У теропод эти структуры, как правило, консервативны, что говорит о том, что функциональные требования во время передвижения были схожи у разных видов. Самый передний из полукружных каналов был увеличен, что обычно встречается у двуногих животных. Ориентация бокового полукружного канала указывает на то, что голова и шея удерживались горизонтально в нейтральном положении. [56]
Голотип C. nasicornis был найден со сросшимися левыми плюсневыми костями II–IV. [57] Марш в 1884 году посвятил короткую статью этой неизвестной в то время особенности у динозавров, отметив близкое сходство с состоянием, наблюдаемым у современных птиц. [34] Наличие этой особенности у Ceratosaurus стало спорным в 1890 году, когда Георг Баур предположил, что слияние у голотипа было результатом зажившего перелома . Это утверждение повторил в 1892 году Коуп, утверждая, что C. nasicornis следует классифицировать как вид Megalosaurus из-за недостаточных анатомических различий между этими родами. [35] Однако с тех пор были описаны примеры сросшихся плюсневых костей у динозавров, которые не имеют патологического происхождения, включая таксоны более базальные, чем Ceratosaurus . [57] Осборн в 1920 году объяснил, что не наблюдается аномального роста костей, и что сращение необычно, но, вероятно, не патологическое. [2] : 112 Рональд Раткевич в 1976 году утверждал, что это сращение ограничило способность животного к бегу, но это утверждение было отвергнуто Полом в 1988 году, который отметил, что та же самая особенность встречается у многих быстро движущихся животных сегодня, включая наземных птиц и копытных. [7] Анализ 1999 года, проведенный Дарреном Танке и Брюсом Ротшильдом, показал, что сращение действительно было патологическим, что подтвердило более раннее утверждение Баура. [57] Другие сообщения о патологиях включают в себя стрессовый перелом в кости стопы, отнесенной к роду, [58], а также сломанный зуб неопознанного вида цератозавра , который показывает признаки дальнейшего износа, полученного после перелома. [57]

Все находки североамериканских цератозавров происходят из формации Моррисон, последовательности мелководных морских и (преимущественно) аллювиальных осадочных пород на западе Соединенных Штатов и самого плодородного источника костей динозавров на континенте. Согласно радиометрическому датированию , возраст формации колеблется от 156,3 миллионов лет у ее основания [59] до 146,8 миллионов лет в верхней части [60] , что помещает ее в поздний оксфордский , кимериджский и ранний титонский ярусы поздней юры. Цератозавр известен из кимериджских и титонских слоев формации. [20] : 49 Формация Моррисон интерпретируется как полузасушливая среда с четко выраженными влажными и сухими сезонами . Бассейн Моррисона простирался от Нью-Мексико до Альберты и Саскачевана, образовавшись, когда предшественники Передового хребта Скалистых гор начали продвигаться на запад. Отложения из их обращенных на восток водосборных бассейнов переносились ручьями и реками и отлагались в болотистых низинах, озерах, речных руслах и поймах. [61] Эта формация по возрасту похожа на формацию Лориньян в Португалии и формацию Тендагуру в Танзании. [62]
Формация Моррисон фиксирует среду и время, в течение которых доминировали гигантские динозавры-завроподы. [63] Другие динозавры, известные из Моррисон, включают тероподов Koparion , Stokesosaurus , Ornitholestes , Allosaurus и Torvosaurus , завропод Apatosaurus , Brachiosaurus , Camarasaurus и Diplodocus , а также птицетазовых Camptosaurus , Dryosaurus , Nanosaurus , Gargoyleosaurus и Stegosaurus . [64] Аллозавр , на долю которого приходилось от 70 до 75% всех образцов теропод, находился на верхнем трофическом уровне пищевой сети Моррисон . [44] Другие позвоночные, которые разделяли эту палеосреду, включали лучепёрых рыб , лягушек , саламандр , черепах, таких как Uluops , сфенодонтов , ящериц , наземных и водных крокодиломорфов, таких как Hoplosuchus , и несколько видов птерозавров, таких как Harpactognathus и Mesadactylus . Раковины двустворчатых моллюсков и водных улиток также распространены. Флора того периода была обнаружена по окаменелостям зелёных водорослей , грибов , мхов , хвощей , цикадовых , гинкго и нескольких семейств хвойных . Растительность варьировалась от лесов, выстилающих реки, из древовидных папоротников и папоротников ( галерейные леса ) до папоротниковых саванн с редкими деревьями, такими как хвойное дерево Brachyphyllum, похожее на араукарию . [65]
Частичный образец Ceratosaurus указывает на присутствие рода в португальском члене Порту-Ново формации Лориньян. Многие из динозавров формации Лориньян либо являются теми же родами, что и те, что были обнаружены в формации Моррисон, либо имеют близкого аналога. [62] Помимо Ceratosaurus , исследователи также отметили, что присутствие Allosaurus и Torvosaurus в португальских породах в основном известно из Моррисона, в то время как Lourinhanosaurus до сих пор был зарегистрирован только из Португалии. Травоядные динозавры из члена Порту-Ново включают, среди прочего, завропод Dinheirosaurus и Zby , а также стегозавра Miragaia . [66] [22] [23] Во время поздней юры Европа только что была отделена от Северной Америки все еще узким Атлантическим океаном . Португалия, как часть Пиренейского полуострова , все еще была отделена от других частей Европы. По мнению Матеуса и коллег, сходство между португальской и североамериканской фаунами теропод указывает на наличие временного сухопутного моста , допускающего фаунистический обмен. [22] [23] Однако Малафая и коллеги выдвинули более сложный сценарий, поскольку другие группы, такие как зауроподы, черепахи и крокодилы, демонстрируют явно разный видовой состав в Португалии и Северной Америке. Таким образом, начальное разделение этих фаун могло привести к взаимообмену в некоторых, но аллопатрическому видообразованию в других группах. [24]
{{cite book}}: |journal=проигнорировано ( помощь )