Копытные ( / ˈ ʌ ŋ ɡ j ʊ l eɪ t s , - ɡ j ə -, - l ɪ t s , - l ə t s / UNG -gyuu-layts, -gyə-, -lits, -ləts ) являются членами разнообразной клады Euungulata («истинные копытные»), которая в основном состоит из крупных млекопитающих с копытами . Когда-то часть клады «Ungulata» вместе с кладой Paenungulata , «Ungulata» с тех пор была определена как полифилетическая и, следовательно, недействительная клада на основе молекулярных данных. В результате настоящие копытные с тех пор были переклассифицированы в более новую кладу Euungulata в 2001 году в пределах клады Laurasiatheria , в то время как Paenungulata были переклассифицированы в отдаленную кладу Afrotheria . [4] Современные копытные делятся на два отряда: Perissodactyla , включающие лошадей , носорогов и тапиров ; и Artiodactyla, включающие крупный рогатый скот , антилоп , свиней , жирафов , верблюдов , овец , оленей и бегемотов , среди прочих. Китообразные, такие как киты , дельфины и морские свиньи, также классифицируются как парнокопытные, хотя у них нет копыт. Большинство наземных копытных используют копытные кончики пальцев ног для поддержки веса тела во время стояния или движения. Два других отряда копытных, Notoungulata и Litopterna , оба обитавшие в Южной Америке, вымерли в конце плейстоцена, около 12 000 лет назад.
Термин означает, грубо говоря, «имеющий копыта» или «копытное животное». Как описательный термин, «копытное» обычно исключает китообразных, поскольку они не обладают большинством типичных морфологических характеристик других копытных, но недавние открытия указывают на то, что они также произошли от ранних парнокопытных . [5] Копытные, как правило, травоядны, и многие используют специализированные кишечные бактерии, чтобы переваривать целлюлозу, хотя некоторые представители могут отклоняться от этого: несколько видов свиней и вымершие энтелодонты всеядны, в то время как китообразные и вымершие мезонихии являются плотоядными.
Копытное происходит от позднелатинского прилагательного ungulatus ' копытный ' . Ungulatus - уменьшительная форма латинского unguis ' ноготь ' (ноготь пальца руки; ноготь пальца ноги). [6]
Euungulata — это клад (или в некоторых таксономиях большой отряд) млекопитающих. Два существующих отряда копытных — Perissodactyla (непарнокопытные) и Artiodactyla (парнокопытные) . Hyracoidea (даманы) , Sirenia (морские коровы) (дюгони и ламантины) и Proboscidea (слоны) в прошлом были сгруппированы в кладе «Ungulata», позже обнаруженной как полифилетическая и теперь недействительная клада. Три отряда Paenungulata теперь считаются кладой и сгруппированы в кладе Afrotheria, в то время как Euungulata теперь сгруппированы в кладе Laurasiatheria . [1]
В 2009 году морфологические [7] [8] [9] [10] и молекулярные [11] [12] исследования показали, что трубкозубы, даманы, морские коровы и слоны были более тесно связаны друг с другом и с сенгисами , тенреками и златокротами , чем с непарнокопытными и парнокопытными, и образуют кладу Afrotheria . Слоны, морские коровы и даманы были сгруппированы вместе в кладу Paenungulata , в то время как трубкозуб считался либо близким родственником им, либо близким родственником сенгисов в кладе Afroinsectiphilia . [13] Это яркий пример конвергентной эволюции . [14]
В настоящее время ведутся споры о том, является ли эта меньшая группа Euungulata кладистической (основанной на эволюции) группой или просто фенетической группой ( форма таксона ) или народным таксоном (похожей, но не обязательно родственной). Некоторые исследования действительно обнаружили, что мезаксонские копытные и параксонские копытные образуют монофилетическую линию, [15] [16] [17] тесно связанную либо с Ferae ( хищники и панголины ) [18] [19] в кладе Fereuungulata, либо с летучими мышами . [20] Другие исследования обнаружили, что эти два отряда не так уж и тесно связаны, поскольку некоторые относят непарнокопытных к близким родственникам летучих мышей, а Ferae к Pegasoferae [21] , а другие относят парнокопытных к близким родственникам летучих мышей. [22]






Ниже приведена упрощенная таксономия (предполагая, что копытные действительно образуют естественную группу) с существующими семействами в порядке родства. Имейте в виду, что все еще были некоторые серые зоны конфликта, такие как случай с родством между семействами пекорана и семействами усатых китов . Смотрите каждое семейство для родства видов, а также противоречий в их соответствующих статьях.
Ниже представлено общее мнение о филогении семейств копытных. [22] [23]


Perissodactyla и Artiodactyla включают большинство крупных наземных млекопитающих. Эти две группы впервые появились в конце палеоцена , быстро распространившись на большое количество видов на многочисленных континентах, и с тех пор развивались параллельно. Некоторые ученые полагали, что современные копытные произошли от эволюционной ступени млекопитающих, известных как кондиларты . [24] Самым ранним известным членом этой группы, возможно, был крошечный Protungulatum , млекопитающее, которое сосуществовало с последним из нептичьих динозавров 66 миллионов лет назад. [25] Однако многие специалисты не считают его настоящим плацентарным, не говоря уже о копытном. [26] Загадочные диноцераты были одними из первых крупных травоядных млекопитающих, хотя их точные отношения с другими млекопитающими все еще обсуждаются, и одна из теорий заключается в том, что они могут быть просто дальними родственниками современных копытных; самое последнее исследование восстанавливает их как часть истинного сообщества копытных, наиболее близкого к Carodnia . [27]
В Австралии недавно вымерший сумчатый хамелеон (Chaeropus) также развил копыта, похожие на копыта парнокопытных [28] , что является примером конвергентной эволюции .

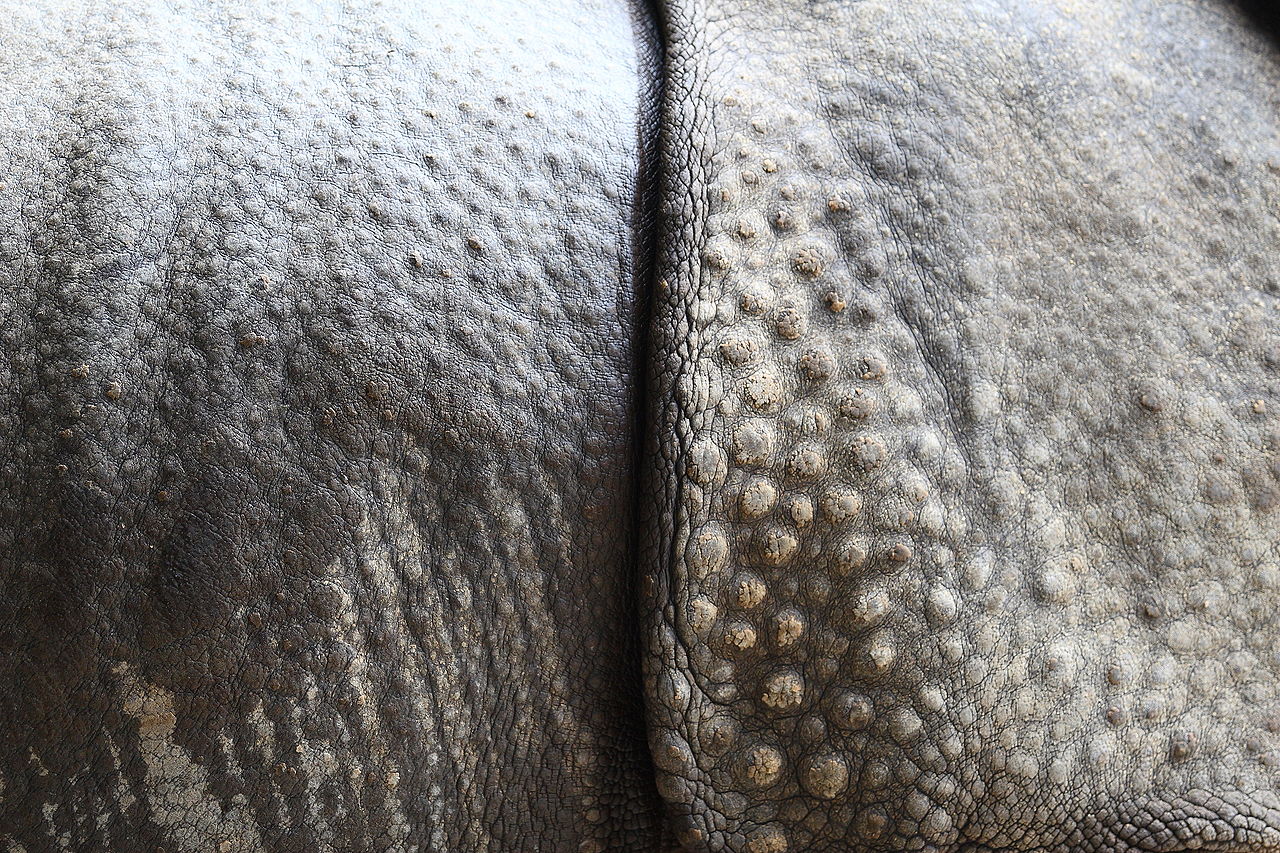
Считалось, что непарнокопытные произошли от Phenacodontidae , небольших животных размером с овцу, которые уже демонстрировали признаки анатомических особенностей, которые унаследуют их потомки (например, редукция пальцев I и V). [30] К началу эоцена , 55 миллионов лет назад (Mya), они разнообразились и распространились, заняв несколько континентов. Лошади и тапиры оба эволюционировали в Северной Америке; [31] носороги, по-видимому, произошли в Азии от животных, похожих на тапиров, а затем колонизировали Америку в среднем эоцене (около 45 Mya). Из примерно 15 семейств выжили только три (McKenna and Bell, 1997; Hooker, 2005). Эти семейства были очень разнообразны по форме и размеру; они включали огромных бронтотериев и причудливых халикотериев . Самый большой непарнокопытный, азиатский носорог, называемый Paraceratherium , достигал веса 15 тонн (17 тонн), что более чем в два раза превышало вес слона . [32]
В кладистическом исследовании было обнаружено, что антракобуниды и десмостилии — две линии, которые ранее классифицировались как афротерии (точнее, ближе к слонам) — были классифицированы как клада, тесно связанная с непарнокопытными. [2] Десмостилии были крупными земноводными четвероногими с массивными конечностями и коротким хвостом. [33] [ отсутствует длинная цитата ] Они вырастали до 1,8 метра (6 футов) в длину и, как считалось, весили более 200 килограммов (440 фунтов). Их ископаемые останки были известны от северного побережья Тихого океана , [34] от южной Японии через Россию , Алеутские острова и тихоокеанское побережье Северной Америки до южной оконечности Нижней Калифорнии . Их зубная и скелетная форма предполагает, что десмостилии были водными травоядными, зависящими от прибрежных местообитаний. Их название происходит от их весьма отличительных коренных зубов, в которых каждый выступ был преобразован в полые колонны, так что типичный коренной зуб напоминал бы скопление труб или, в случае изношенных коренных зубов, вулканы. Они были единственными морскими млекопитающими, которые вымерли.
Южноамериканские меридиангуляты включают несколько похожих на тапира пиротериев и астрапотериев , мезаксонических литоптернов и разнообразных нотоунгулятов . В целом, меридиангуляты, как говорят, произошли от животных, таких как Hyopsodus . [30] Некоторое время их отношения с другими копытными были загадкой. Некоторые палеонтологи даже оспаривали монофилию меридиангулят, предполагая, что пиротериев могут быть более тесно связаны с другими млекопитающими, такими как Embrithopoda (африканский отряд, который был связан со слонами ), чем с другими южноамериканскими копытными. [35] Недавнее исследование, основанное на костном коллагене, показало, что по крайней мере литоптерны и нотоунгуляты были тесно связаны с непарнокопытными. [36]
Древнейшие известные окаменелости , отнесенные к Equidae, датируются ранним эоценом , 54 миллиона лет назад. Их отнесли к роду Hyracotherium , но типовой вид этого рода теперь не считается членом этого семейства, а другие виды были разделены на разные роды. Эти ранние Equidae были животными размером с лисицу с тремя пальцами на задних лапах и четырьмя на передних. Они были травоядными, питающимися относительно мягкими растениями, и уже были приспособлены к бегу. Сложность их мозга предполагает, что они уже были внимательными и умными животными. [37] Более поздние виды уменьшили количество пальцев и развили зубы, более подходящие для измельчения травы и другой жесткой растительной пищи.
Носороги отделились от других непарнокопытных к раннему эоцену. Ископаемые останки Hyrachyus eximus, найденные в Северной Америке, относятся к этому периоду. Этот небольшой безрогий предок больше напоминал тапира или небольшую лошадь, чем носорога. Три семейства, иногда объединяемые в надсемейство Rhinocerotoidea, эволюционировали в позднем эоцене: Hyracodontidae , Amynodontidae и Rhinocerotidae , тем самым создав взрыв разнообразия, не имевший себе равных некоторое время, пока изменения окружающей среды радикально не уничтожили несколько видов.
Первые тапириды, такие как Heptodon , появились в раннем эоцене. [38] Они выглядели очень похожими на современные формы, но были примерно в два раза меньше и не имели хоботка. Первые настоящие тапиры появились в олигоцене . К миоцену такие роды, как Miotapirus, были почти неотличимы от современных видов. Считалось, что азиатские и американские тапиры разошлись около 20–30 миллионов лет назад; а тапиры мигрировали из Северной Америки в Южную Америку около 3 миллионов лет назад в рамках Великого американского обмена . [39]
Непарнокопытные были доминирующей группой крупных наземных травоядных вплоть до олигоцена. Однако рост трав в миоцене (около 20 млн лет назад) привел к серьезным изменениям: виды парнокопытных с их более сложными желудками были лучше приспособлены к грубой, низкокалорийной диете и вскоре стали выдающимися. Тем не менее, многие виды непарнокопытных выживали и процветали до позднего плейстоцена (около 10 000 лет назад), когда они столкнулись с давлением человеческой охоты и изменением среды обитания.

Считалось, что парнокопытные произошли от небольшой группы кондилартов, Arctocyonidae , которые были неспециализированными, внешне похожими на енотов или медведей всеядными животными из раннего палеоцена (около 65–60 миллионов лет назад). У них были относительно короткие конечности без специализаций, связанных с их родственниками (например, редуцированные боковые пальцы, сросшиеся кости и копыта), [30] и длинные, тяжелые хвосты. Их примитивная анатомия делает маловероятным, что они могли преследовать добычу, но с их мощными пропорциями, когтями и длинными клыками они могли одолеть более мелких животных во время неожиданных атак. [30] Очевидно, эти млекопитающие вскоре эволюционировали в две отдельные линии: мезонихий и парнокопытных.

Первые парнокопытные выглядели как современные оленьки или свиньи: маленькие, коротконогие существа, которые питались листьями и мягкими частями растений . К позднему эоцену (46 миллионов лет назад) уже развились три современных подотряда: Suina ( группа свиней ); Tylopoda ( группа верблюдов ); и Ruminantia ( группа коз и крупного рогатого скота ). Тем не менее, парнокопытные были далеки от того, чтобы доминировать в то время: непарнокопытные были гораздо более успешными и гораздо более многочисленными. Парнокопытные выживали в нишевых ролях, обычно занимая пограничные местообитания , и, предположительно, именно в это время у них развилась сложная пищеварительная система , которая позволяла им выживать на низкосортной пище. В то время как большинство парнокопытных занимали ниши, оставленные несколькими вымершими непарнокопытными, одна линия парнокопытных начала выходить в моря.

Традиционная теория эволюции китообразных заключалась в том, что китообразные были связаны с мезонихиями . У этих животных были необычные треугольные зубы, очень похожие на зубы примитивных китообразных. Вот почему ученые долгое время считали, что китообразные произошли от формы мезонихий. Сегодня многие ученые считают, что китообразные произошли от той же группы, которая дала начало бегемотам. Эта предполагаемая предковая группа, вероятно, разделилась на две ветви около 54 миллионов лет назад . [5] Одна ветвь эволюционировала в китообразных , возможно, начавшись около 52 миллионов лет назад с протокита Pakicetus и других ранних предков китообразных, известных как Archaeoceti , которые в конечном итоге претерпели водную адаптацию в полностью водных китообразных . [40] Другая ветвь стала антракотериями , большим семейством четвероногих зверей, самые ранние из которых в позднем эоцене напоминали тощих бегемотов со сравнительно маленькими и узкими головами. Все ветви антракотериев, за исключением той, которая эволюционировала в Hippopotamidae , вымерли в плиоцене , не оставив потомков. [41]
Семейство Raoellidae считается самым близким к китообразным семейством парнокопытных. [42] [43] Соответственно, новые теории эволюции китообразных предполагают, что киты и их предки избежали хищничества, а не конкуренции, постепенно приспосабливаясь к океану. [44] [45] [46]

Мезонихии изображались как «волки на копытах» и были первыми крупными хищниками среди млекопитающих, появившимися в палеоцене. [47] Ранние мезонихии имели пять пальцев на ногах, которые, вероятно, опирались на землю во время ходьбы ( стопохождение ), но более поздние мезонихии имели четыре пальца, которые заканчивались крошечными копытцами на всех пальцах ног и были все более приспособлены к бегу. Подобно бегущим представителям парнокопытных, мезонихии ( например, Pachyaena ) ходили на своих пальцах ( пальцехождение ). [47] Мезонихии очень плохо себя чувствовали в конце эпохи эоцена, и только один род, Mongolestes , [48] выжил в эпоху раннего олигоцена , поскольку изменился климат и возникла жесткая конкуренция со стороны более приспособленных креодонтов .

Копытные животные были в большом разнообразии в ответ на половой отбор и экологические события; у большинства копытных отсутствует ключичная кость . [49] Наземные копытные были по большей части травоядными, некоторые из них были травоядными . Однако были исключения из этого правила, так как свиньи, пекари, бегемоты и дукеры, как известно, имели всеядную диету. Некоторые китообразные были единственными современными копытными, которые были плотоядными; усатые киты потребляют значительно меньших животных по отношению к размеру своего тела, таких как мелкие виды рыб и криль ; зубатые киты, в зависимости от вида, могут потреблять широкий спектр видов: кальмаров , рыбу, акул и другие виды млекопитающих, такие как тюлени и другие киты. С точки зрения экосистемы копытные колонизировали все уголки планеты, от гор до океанских глубин ; от лугов до пустынь , а некоторые были одомашнены людьми .
У копытных животных развились специализированные адаптации, особенно в области придатков черепа, зубной системы и морфологии ног, включая модификацию таранной кости (одной из костей лодыжки на конце голени) с короткой, крепкой головкой.
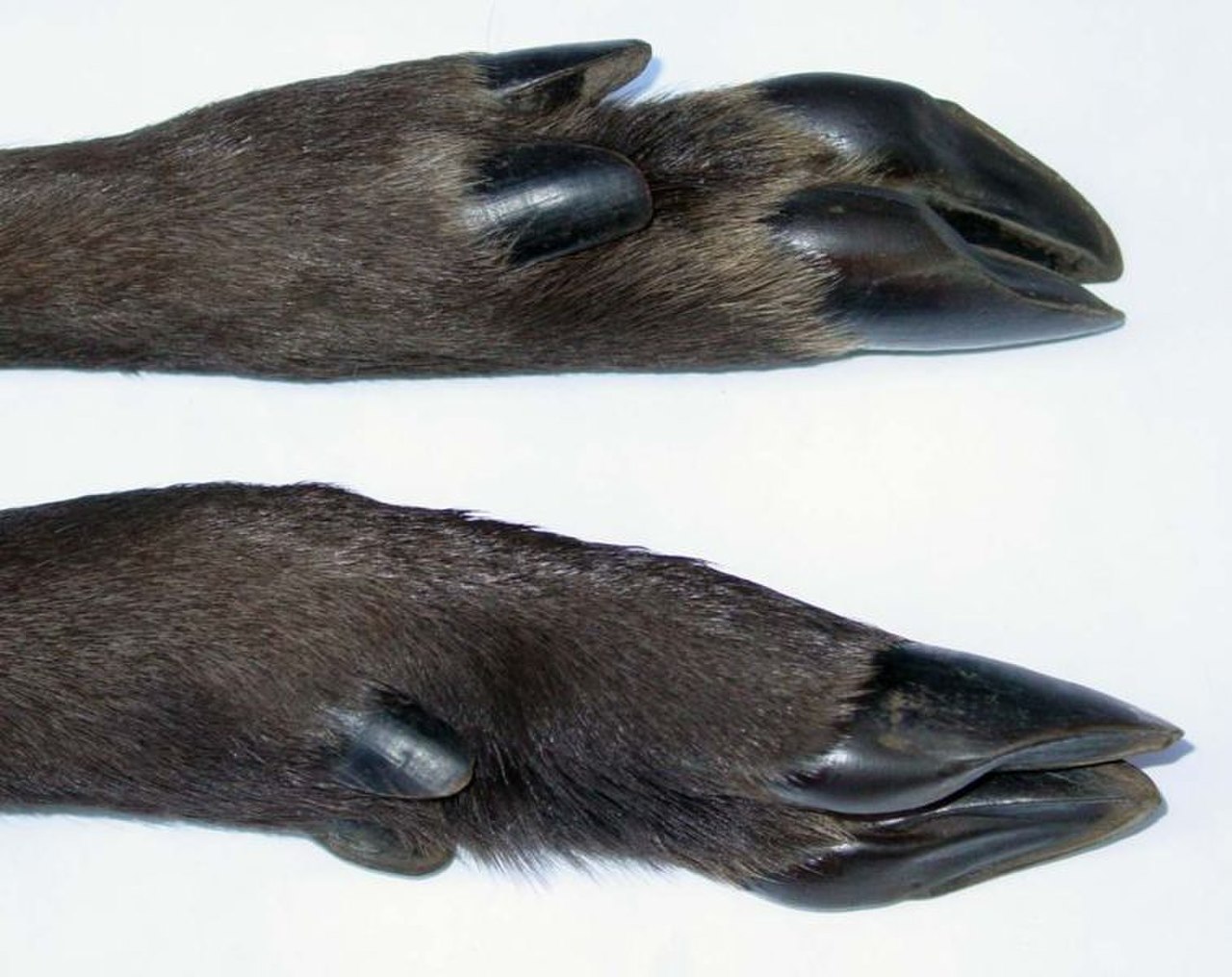
Копыто — это кончик пальца копытного млекопитающего , укрепленный толстым роговым ( кератиновым ) покрытием. Копыто состоит из твердой или резиновой подошвы и твердой стенки, образованной толстым гвоздем , намотанным на кончик пальца. Как подошва, так и край стенки копыта обычно выдерживают вес животного. Копыта постоянно растут и постоянно изнашиваются при использовании. У большинства современных копытных лучевая и локтевая кости сращены по длине передней конечности; ранние копытные, такие как арктоциониды , не имели этой уникальной скелетной структуры. [50] Сращение лучевой и локтевой костей не позволяет копытному вращать переднюю конечность. Поскольку эта скелетная структура не имеет определенной функции у копытных, она считается гомологичной характеристикой, которую копытные разделяют с другими млекопитающими. Эта черта могла быть передана от общего предка. Хотя два отряда копытных в разговорной речи были основаны на количестве пальцев у их членов («непарнокопытные» для непарнокопытных и «четнопалые» для наземных парнокопытных), это не точная причина, по которой они были сгруппированы. У тапиров четыре пальца спереди, но они были членами отряда «непарнокопытных»; пекари и современные китообразные были членами отряда «парнопалых», но у пекари три пальца спереди, а киты были крайним примером, поскольку у них вместо копыт ласты. Ученые классифицировали их в соответствии с распределением их веса на пальцы ног.
У непарнокопытных мезаксоническая стопа, что означает, что вес распределяется на третий палец на всех ногах благодаря плоской симметрии их стоп. Произошла редукция пальцев от общего предка, классическим примером являются лошади с их одинарными копытами. В результате появилось альтернативное название для непарнокопытных — почти устаревшее Mesaxonia. Непарнокопытные были не единственной линией млекопитающих, у которых развилась эта черта; меридиангуляты развили мезаксоническую стопу много раз.
Наземные парнокопытные имеют параксоническую стопу, что означает, что вес распределяется на третий и четвертый пальцы на всех ногах. Большинство этих млекопитающих имеют раздвоенные копыта, с двумя меньшими, известными как прибылые пальцы , которые были расположены выше на ноге. Самые ранние китообразные ( археоцеты ), также имели эту характеристику в дополнение к тому, что также имели как астрагал , так и кубовидную кость в лодыжке, которые были дополнительными диагностическими признаками парнокопытных. [51]

У современных китообразных передние конечности стали грудными плавниками , а задние были внутренними и редуцированными. Иногда гены, кодирующие более длинные конечности, заставляют современных китообразных развивать миниатюрные ноги (известно как атавизм ). Основной способ передвижения — движение вверх-вниз хвостовым плавником, называемым хвостовым плавником, который используется для продвижения , в то время как грудные плавники вместе со всей хвостовой частью обеспечивают контроль направления. Все современные китообразные по-прежнему сохраняют свои пальцы, несмотря на внешний вид, предполагающий обратное.
У большинства копытных развились редуцированные клыки и специализированные моляры , включая бунодонтные (низкие, округлые бугорки) и гипсодонтные (высокие коронки) зубы. Развитие гипсодонтии представляло особый интерес, поскольку эта адаптация была тесно связана с распространением лугов в миоцене около 25 миллионов лет назад. По мере того, как лесные биомы приходили в упадок, луга расширялись, открывая новые ниши для млекопитающих. Многие копытные перешли от поедания трав к выпасу, и, возможно, из-за абразивного кремнезема в траве гипсодонтия стала обычным явлением. Однако недавние доказательства связывают эволюцию гипсодонтии с открытыми, песчаными местообитаниями, а не с самой травой. Это называется гипотезой «песок, а не трава» . [52]
У некоторых копытных полностью отсутствуют верхние резцы, а вместо них есть зубная подушечка , помогающая при ощипывании. [53] [54] Это можно обнаружить у верблюдов, жвачных животных и некоторых зубатых китов; современные усатые киты были примечательны тем, что у них вместо кита есть ус, чтобы отфильтровывать криль из воды. С другой стороны, зубы эволюционировали как оружие или сексуальное проявление, наблюдаемое у свиней и пекари, некоторых видов оленей, кабарги, бегемотов, клюворылых китов и нарвала с его длинным клыком. [55]

У копытных животных развилось множество черепных придатков, которые можно найти у оленьих (за исключением кабарги). У быков и антилоп размер и форма рогов сильно различаются, но основная структура всегда представляет собой пару простых костных выступов без ответвлений, часто имеющих спиральную, скрученную или рифленую форму, каждый из которых покрыт постоянной оболочкой из кератина . Уникальная структура рогов является единственной однозначной морфологической особенностью полорогих, которая отличает их от других пекоранов . [56] [57] Развитие рогов у самцов связано с половым отбором, [58] [59] в то время как наличие рогов у самок, вероятно, обусловлено естественным отбором. [58] [60] Рога самок обычно меньше, чем у самцов, и иногда имеют другую форму. Считается, что рога самок полорогих животных развились для защиты от хищников или для выражения территориальности, поскольку у нетерриториальных самок, которые способны использовать криптос для защиты от хищников, рога часто отсутствуют. [60]
Рога носорога, в отличие от рогов других рогатых млекопитающих, состоят только из кератина. Эти рога опираются на носовой гребень черепа животного.
Рога уникальны для оленьих и встречаются в основном у самцов: единственные самки оленьих с рогами — это карибу и северные олени , у которых рога обычно меньше, чем у самцов. Тем не менее, плодовитые самки других видов оленей иногда способны производить рога, обычно из-за повышенного уровня тестостерона. [61] Каждый рог растет из точки крепления на черепе, называемой ножкой. Пока рог растет, он покрыт высокососудистой кожей , называемой бархатом, которая снабжает кислородом и питательными веществами растущую кость. [62] Рога считаются одним из самых преувеличенных случаев мужских вторичных половых признаков в животном мире, [63] и растут быстрее, чем любая другая кость млекопитающих. [64] Рост происходит на кончике, изначально как хрящ, который затем минерализуется, чтобы стать костью. Как только рог достигает своего полного размера, бархат теряется, и кость рога умирает. Эта мертвая костная структура — зрелый рог. В большинстве случаев кость у основания разрушается остеокластами , и рога в конечном итоге отпадают. [62] В результате быстрого роста рога предъявляют существенные требования к питанию оленей; таким образом, они могут представлять собой честный сигнал об эффективности метаболизма и способности собирать пищу. [65]
Оссиконы — это роговидные (или похожие на оленьи рога) выросты, которые можно обнаружить на головах жирафов и самцов окапи . Они похожи на рога антилоп и крупного рогатого скота , за исключением того, что они происходят из окостеневшего хряща [66] и что оссиконы остаются покрытыми кожей и мехом , а не рогом.
Черепные придатки вилорога уникальны. Каждый «рог» вилорога состоит из тонкой, сплющенной с боков пластины кости, которая растет из лобных костей черепа, образуя постоянное ядро. Как и у жирафовых, кожа покрывает костные ядра, но у вилорога она развивается в кератиновую оболочку, которая сбрасывается и отрастает заново ежегодно. В отличие от рогов семейства полорогих, роговые оболочки вилорога разветвлены, каждая оболочка имеет направленный вперед зубец (отсюда и название вилорог). Рога самцов хорошо развиты.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )