Кортико -базальные ганглии-таламо-кортикальная петля ( петля CBGTC ) представляет собой систему нейронных цепей в мозге . Петля включает связи между корой , базальными ганглиями , таламусом и обратно в кору. Она имеет особое значение для гиперкинетических и гипокинетических двигательных расстройств , таких как болезнь Паркинсона и болезнь Хантингтона , [1], а также для психических расстройств контроля, таких как синдром дефицита внимания и гиперактивности (СДВГ), [2], обсессивно-компульсивное расстройство (ОКР), [3] и синдром Туретта . [4]
Петля CBGTC в основном состоит из модуляторных дофаминергических проекций от pars compacta черной субстанции и вентральной области покрышки , а также возбуждающих глутаматергических проекций от коры к полосатому телу , где эти проекции образуют синапсы с возбуждающими и тормозными путями, которые возвращаются обратно в кору. Первоначально эта петля была предложена как часть модели базальных ганглиев, называемой моделью параллельной обработки , которая была подвергнута критике и модифицирована в другую модель, называемую моделью центра-окружения . [5]
Текущие схемы организации характеризуют взаимодействия кортико-базальных ганглиев как сегрегированную параллельную обработку, что означает, что существует небольшая конвергенция отдельных корковых областей в базальных ганглиях. Считается, что это объясняет топографически организованную функциональность полосатого тела. [4] Полосатое тело организовано по ростро-каудальной оси, при этом ростральная скорлупа и хвостатое тело обслуживают ассоциативные и когнитивные функции, а каудальные области обслуживают сенсомоторную функцию. [6] Иногда, когда выраженной целью является полосатое тело, петлю называют кортико-стриатно-таламо-кортикальной петлей . [7]
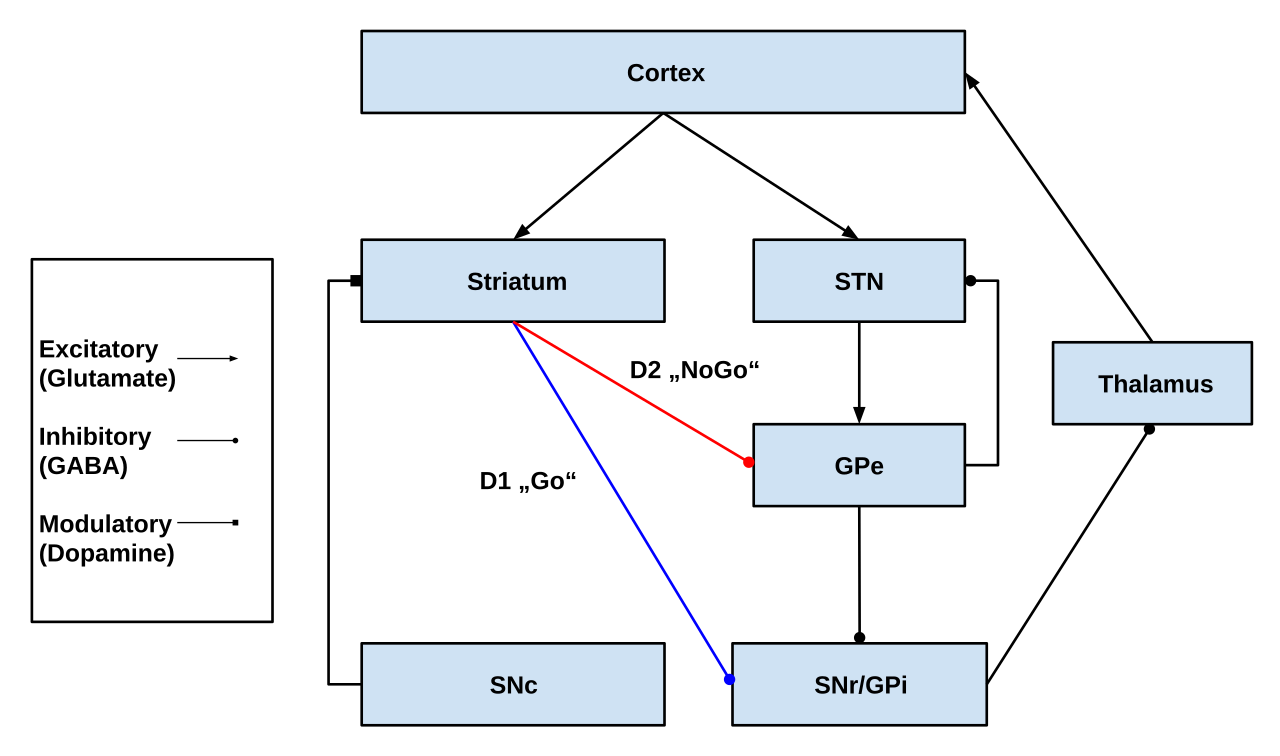
Две основные входные структуры контура — полосатое тело и субталамическое ядро (STN). Полосатое тело получает входные сигналы как от коры, так и от компактной части черной субстанции ( SNc), тогда как STN получает только корковые входные сигналы.
Из полосатого тела выходят два пути. Один путь называется непрямым (или NoGo) путем и является тормозным. Он проецируется на наружный бледный шар (GPe) и тормозит его, что приводит к растормаживанию внутреннего бледного шара (GPi), что приводит к торможению таламуса. Этот путь также, в результате торможения GPe, растормаживает субталамическое ядро, что приводит к возбуждению GPi и, следовательно, к торможению таламуса.
Второй путь называется прямым (или Go) путем и является возбуждающим. Этот путь ингибирует GPi, что приводит к растормаживанию таламуса. Прямой путь в основном состоит из моносинаптических связей, управляемых дофаминовым рецептором D1 , аденозиновым рецептором A1 и мускариновым ацетилхолиновым рецептором M4 , в то время как непрямой путь опирается на связи, управляемые дофаминовым рецептором D2 , аденозиновым рецептором A2A и мускариновым ацетилхолиновым рецептором M1 . [1] [8]
Параллельные петли CBGTC были разделены в соответствии с функциями связанных корковых областей. Одна схема включает разделение на лимбические и моторные петли, при этом моторные петли содержат непрямые и прямые пути, которые, в свою очередь, взаимосвязаны с лимбической петлей, которая проецируется в вентральный стриатум. [9] Петля также была разделена на лимбическую, ассоциативную, глазодвигательную и моторную цепи [4] для объяснения роли дофамина в базальных ганглиях в мотивационных состояниях. [10] Пятипетлевое разделение, основанное на первичных корковых мишенях, было описано следующим образом: [11]
Проблема, выявленная в текущей анатомии схемы, заключается в том, что временная задержка между прямыми и непрямыми путями должна приводить к тому, что эта схема не работает. Чтобы преодолеть это, гипотеза центрального окружения предполагает, что гиперпрямой путь от коры будет подавлять другие входы, помимо одного сфокусированного коркового входа. Однако синхронизация активности базальных ганглиев и момент конечности, а также исследования повреждений не подтверждают эту гипотезу [12]
Для объяснения того, как действия выбираются в базальных ганглиях, были предложены две модели. Модель актер-критик предполагает, что действия генерируются и оцениваются «критиком» в вентральном полосатом теле, в то время как действия выполняются «актером» в дорсальном полосатом теле. Другая модель предполагает, что базальные ганглии действуют как механизм отбора, где действия генерируются в коре и выбираются на основе контекста базальными ганглиями. [13] Цикл CBGTC также участвует в дисконтировании вознаграждения, причем активность увеличивается при неожиданном или большем, чем ожидалось, вознаграждении. [2] Один обзор поддержал идею о том, что кора участвует в обучении действиям независимо от их результата, в то время как базальные ганглии участвуют в выборе соответствующих действий на основе ассоциативного вознаграждения, основанного на обучении методом проб и ошибок. [14]
Петля CBGTC вовлечена во многие заболевания. Например, при болезни Паркинсона дегенерация дофаминергических нейронов, приводящая к снижению активности возбуждающего пути, как полагают, приводит к гипокинезии, [15] а при болезни Хантингтона дегенерация ГАМКергических нейронов, управляющих тормозным путем, как полагают, приводит к судорожным движениям тела. [2] Кодегенерация лимбических проекций вместе с моторными проекциями может приводить ко многим психиатрическим симптомам этих в первую очередь моторных заболеваний. [9] При ОКР петля может быть дисфункциональной, с дисбалансом между косвенными и прямыми путями, приводящим к нежелательным мыслям , которые «застревают». [3] При СДВГ снижение тонической дофаминергической сигнализации, приводящее к чрезмерному обесцениванию отсроченных вознаграждений, как полагают, приводит к снижению внимания. [2]
Петля CBGTC изучалась в связи с сознанием , выбором действия, в связи с другими цепями и в контексте памяти и познания. [16] [17] Модель петли CBGTC критиковалась как чрезмерно упрощенная и слишком жестко применяемая, учитывая доказательства анатомического и функционального перекрытия и взаимодействия между прямыми и непрямыми путями. [18] Петля также исследовалась в контексте глубокой стимуляции мозга . [16] По состоянию на 2013 год велись интенсивные дебаты относительно разделения цепи, взаимодействия путей, количества путей и общей анатомии. [17]
{{cite journal}}: Цитировать журнал требует |journal=( помощь )