Криль ( SG : krill) — мелкие и исключительно морские ракообразные отряда Euphausiacea , встречающиеся во всех мировых океанах. [1] Название «криль» происходит от норвежского слова krill , что означает «мелкая рыбка », [2] которое также часто относят к видам рыб.
Криль считается важным соединением трофического уровня – в самом низу пищевой цепи . Они питаются фитопланктоном и, в меньшей степени, зоопланктоном , а также являются основным источником пищи для многих крупных животных. В Южном океане биомасса одного вида, антарктического криля , составляет около 379 000 000 тонн, [3] что делает его одним из видов с наибольшей общей биомассой. Ежегодно более половины этой биомассы съедают киты, тюлени , пингвины, морские птицы, кальмары и рыбы. Большинство видов криля совершают большие ежедневные вертикальные миграции , обеспечивая тем самым пищу хищникам у поверхности ночью и в более глубоких водах в течение дня.
Криль коммерчески вылавливается в Южном океане и в водах вокруг Японии. Общий мировой вылов составляет 150 000–200 000 тонн в год, большая часть которого добывается в море Скотия . Большая часть улова криля используется для аквакультуры и корма для аквариумов , в качестве наживки в спортивной рыбалке или в фармацевтической промышленности. В Японии, на Филиппинах и в России криль также используется в пищу человеком и известен в Японии как окиами (オキアミ) . Их едят как камарон в Испании и на Филиппинах. На Филиппинах криль также известен как аламан и используется для приготовления соленой пасты под названием багонг .
Отряд Euphausiacea включает два семейства . Более распространенный Euphausiidae содержит 10 различных родов и в общей сложности 85 видов. Из них род Euphausia является самым крупным, насчитывающим 31 вид. [4] Менее известное семейство, Bentheuphausiidae , имеет только один вид , Bentheuphausia amblyops , батипелагический криль, обитающий в глубоких водах ниже 1000 м (3300 футов). Считается самым примитивным из существующих видов криля. [5]
По состоянию на 2013 год [обновлять]отряд Euphausiacea считается монофилетическим из-за нескольких уникальных консервативных морфологических характеристик ( аутапоморфии ), таких как обнаженные нитевидные жабры и тонкие торакоподы [9] , а также результатов молекулярных исследований. [10] [11] [12]
Было много теорий о местонахождении отряда Euphausiacea. С момента первого описания Thysanopode tricuspide Анри Милном-Эдвардсом в 1830 году сходство их двуветвистых торакопод побудило зоологов сгруппировать эвфаузиид и Mysidacea в отряд Schizopoda , который был разделен Йоханом Эриком Вести Боасом в 1883 году на два отдельных отряда. [13] Позже Уильям Томас Калман (1904) причислил Mysidacea к надотряду Peracarida , а эвфаузиииды к надотряду Eucarida , хотя даже до 1930-х годов отстаивался отряд Schizopoda. [9] Позже было также предложено, чтобы отряд Euphausiacea был сгруппирован с Penaeidae (семейство креветок) в Decapoda на основе сходства в развитии, как отметили Роберт Герни и Изабелла Гордон . [14] [15] Причиной этой дискуссии является то, что криль разделяет некоторые морфологические особенности десятиногих и других мизид. [9]
Молекулярные исследования не смогли однозначно сгруппировать их, возможно, из-за нехватки ключевых редких видов, таких как Bentheuphausia amblyops у криля и Amphionides reynaudii у Eucarida. Одно исследование подтверждает монофилию Eucarida (с базальной Mysida), [16] другое группирует Euphausiacea с Mysida (Schizopoda), [11] а еще одно группирует Euphausiacea с Hoplocarida . [17]
График
Ни одна из сохранившихся окаменелостей не может быть однозначно отнесена к Euphausiacea. Некоторые вымершие таксоны эумалакостраков , как полагают, относятся к эвфаузиацеям, таким как Anthracophausia , Crangopsis – теперь отнесенные к Aeschronectida (Hoplocarida) [7] – и Palaeomysis . [18] Все датировки событий видообразования были оценены с помощью методов молекулярных часов , которые установили, что последний общий предок криля семейства Euphausiidae (отряд Euphausiacea минус Bentheuphausia amblyops ) жил в нижнем меловом периоде около 130 миллионов лет назад . [11]
Виды с неритическим распространением включают четыре вида рода Nyctiphanes . [21] Они очень многочисленны вдоль апвеллинговых регионов систем течений Калифорнии , Гумбольдта , Бенгелы и Канарских островов . [22] [23] [24] Другой вид, имеющий только неритическое распространение, — это E. Crystallorophias , который является эндемиком побережья Антарктики. [25]
Виды с эндемичным распространением включают Nyctiphanes capensis , который встречается только в течении Бенгела, [21] E. mucronata в течении Гумбольдта, [26] и шесть видов Euphausia , произрастающих в Южном океане.
В Антарктике известно семь видов: [27] один из рода Thysanoessa ( T. macrura ) и шесть из Euphausia . Антарктический криль ( Euphausia superba ) обычно обитает на глубинах, достигающих 100 м (330 футов) [28] , тогда как ледяной криль ( Euphausia Crystallorophias ) достигает глубины 4000 м (13 100 футов), хотя обычно они обитают на глубинах не более 300–600 м. м (1000–2000 футов). [29] Криль совершает вертикальные миграции (DVM) большими стаями, и акустические данные показали, что эти миграции достигают глубины до 400 метров. [30] Оба встречаются на широтах к югу от 55° ю.ш. , при этом E. Crystallorophias преобладает к югу от 74° ю.ш. [31] и в регионах пакового льда . Другие виды, известные в Южном океане, — E. frigida , E. longirostris , E. triacantha и E. vallentini . [32]
Анатомия и морфология
Объяснение анатомии криля с использованием Euphausia superba в качестве моделиЖабры криля видны снаружи .
У криля сложные сложные глаза . Некоторые виды адаптируются к различным условиям освещения за счет использования экранирующих пигментов . [33]
У них есть две усики и несколько пар грудных ног, называемых переоподами или торакоподами , названными так потому, что они прикреплены к грудной клетке. Их количество варьируется в зависимости от рода и вида. К этим грудным ногам относятся ноги для кормления и ноги для ухода.
Криль, вероятно, является родственной кладой десятиногих, потому что все виды имеют пять пар плавательных ног , которые вместе с последними называются «пловцами», очень похожими на ноги лобстера или пресноводного рака .
Длина большинства взрослых особей криля составляет около 1–2 см (0,4–0,8 дюйма). Некоторые виды вырастают до размеров порядка 6–15 сантиметров (2,4–5,9 дюйма). Самый крупный вид криля, Thysanopoda spinicaudata , обитает глубоко в открытом океане . [34] Криль можно легко отличить от других ракообразных, таких как настоящие креветки , по видимым снаружи жабрам . [35]
За исключением Bentheuphausia amblyops , криль — биолюминесцентные животные, имеющие органы, называемые фотофорами , которые могут излучать свет. Свет генерируется в результате ферментативно -катализируемой реакции хемилюминесценции , в которой люциферин (разновидность пигмента) активируется ферментом люциферазой . Исследования показывают, что люциферин многих видов криля представляет собой флуоресцентный тетрапиррол , аналогичный, но не идентичный люциферину динофлагеллят [ 36] и что криль, вероятно, не производит это вещество сам, а приобретает его как часть своего рациона, содержащего динофлагелляты. [37] Фотофоры криля — это сложные органы с линзами и способностями к фокусировке, которые можно вращать с помощью мышц. [38] Точная функция этих органов пока неизвестна; Возможности включают спаривание, социальное взаимодействие или ориентацию, а также форму камуфляжа против освещения для компенсации их тени от верхнего окружающего света. [39] [40]
Экология
Процессы в биологическом насосе
Фитопланктон преобразует CO2, который растворился из атмосферы в поверхностные слои океанов (90 Гт/год), в твердый органический углерод (РОУ) во время первичного производства (~ 50 Гт С/год). Затем фитопланктон потребляется крилем и мелкими травоядными зоопланктоном, которые, в свою очередь, становятся объектом добычи более высоких трофических уровней. Любой неизрасходованный фитопланктон образует агрегаты и вместе с фекальными гранулами зоопланктона быстро тонет и выносится из смешанного слоя (< 12 Гт С/год). Криль, зоопланктон и микробы перехватывают фитопланктон на поверхности океана и погружают детритные частицы на глубину, потребляя и выдыхая этот POC до CO2 (растворенный неорганический углерод, DIC), так что лишь небольшая часть углерода, произведенного на поверхности, попадает в глубокий океан ( т. е. глубины > 1000 м). В качестве корма для криля и более мелкого зоопланктона они также физически дробят частицы на более мелкие, более медленные или нетонущие кусочки (из-за неаккуратного кормления, копрорексии в случае фрагментации фекалий), замедляя экспорт ВОУ. При этом растворенный органический углерод (DOC) высвобождается либо непосредственно из клеток, либо косвенно посредством бактериальной солюбилизации (желтый кружок вокруг DOC). Затем бактерии могут реминерализовать DOC в DIC (CO2, микробное садоводство). Вертикально мигрирующий криль, более мелкий зоопланктон и рыба могут активно переносить углерод на глубину, потребляя ВОУ в поверхностном слое ночью и метаболизируя его в дневное время, в мезопелагических глубинах проживания. В зависимости от истории жизни вида активный перенос может происходить и на сезонной основе. Числа представляют собой потоки углерода (Гт C год-1) в белых прямоугольниках и массы углерода (Гт С) в темных прямоугольниках. [41]
Кормление
Многие крили являются фильтраторами : [23] их передние придатки , торакоподы, образуют очень тонкие гребни, с помощью которых они могут отфильтровывать пищу из воды. Эти фильтры могут быть очень тонкими у видов (таких как Euphausia spp.), которые питаются в основном фитопланктоном , в частности , диатомовыми водорослями , которые являются одноклеточными водорослями . Криль в основном всеяден , [42] хотя некоторые виды являются плотоядными , питаясь мелким зоопланктоном и личинками рыб . [43]
Криль является важным элементом водной пищевой цепи . Криль преобразует первичную продукцию своей добычи в форму, пригодную для потребления более крупными животными, которые не могут питаться непосредственно мельчайшими водорослями. Северный криль и некоторые другие виды имеют относительно небольшую фильтрующую корзину и активно охотятся на копепод и более крупный зоопланктон. [43]
Хищничество
Крилем питаются многие животные: от мелких животных, таких как рыбы или пингвины, до более крупных, таких как тюлени и усатые киты . [44]
Нарушения экосистемы , приводящие к сокращению популяции криля, могут иметь далеко идущие последствия . Так, во время цветения кокколитофор в Беринговом море в 1998 г. [45] концентрация диатомей в пострадавшем районе снизилась. Криль не может питаться более мелкими кокколитофорами, и, следовательно, популяция криля (в основном E. pacifica ) в этом регионе резко сократилась. Это, в свою очередь, повлияло на другие виды: популяция буревестников сократилась. Считалось, что этот инцидент стал одной из причин, по которой лосось не нерестился в тот сезон. [46]
Несколько одноклеточных эндопаразитоидных инфузорий рода Collinia могут заражать виды криля и опустошать пораженные популяции. Такие заболевания были зарегистрированы для Thysanoessa inermis в Беринговом море, а также для E. pacifica , Thysanoessa spinifera и T. gregaria у тихоокеанского побережья Северной Америки. [47] [48] Некоторые эктопаразиты семейства Dajidae (эпикаридовые изоподы ) поражают криль (а также креветок и мизид ); Одним из таких паразитов является Oculophryxus bicaulis , который был обнаружен на криле Stylocheiron affine и S. longicorne . Он прикрепляется к глазному стеблю животного и высасывает кровь из его головы; очевидно, он подавляет размножение хозяина, поскольку ни одно из больных животных не достигло зрелости. [49]
Изменение климата представляет собой еще одну угрозу для популяций криля. [50]
Пластмассы
Предварительные исследования показывают, что криль может переваривать микропластик диаметром менее 5 мм (0,20 дюйма), расщепляя его и выводя обратно в окружающую среду в более мелкой форме. [51]
История жизни и поведение
Науплиус Euphausia pacifica вылупляется из яйца задом наперед .
Жизненный цикл криля относительно хорошо изучен, несмотря на незначительные различия в деталях от вида к виду. [14] [23] После вылупления криля они проходят несколько личиночных стадий — науплиус , псевдометанауплиус , метанауплиус , калиптопсис и фурцилия , каждая из которых делится на подстадии. Стадия псевдометанауплиуса присуща только видам, которые откладывают яйца в яйцевидный мешок: так называемые «мякоть-нерестовики». Личинки растут и линяют неоднократно по мере развития, заменяя свой жесткий экзоскелет, когда он становится слишком маленьким. Мелкие животные линяют чаще, чем более крупные. Запасы желтка в их организме питают личинок на стадии метанауплиуса.
На стадии калиптопсиса дифференцировка зашла достаточно далеко, чтобы у них появились рот и пищеварительный тракт, и они начали питаться фитопланктоном. К этому времени их запасы желтка исчерпаны, и личинки должны достичь фотической зоны — верхних слоев океана, где процветают водоросли. На стадиях фурцилий добавляются сегменты с парами плавников, начиная с самых передних сегментов. Каждая новая пара становится функциональной только при следующей линьке. Количество сегментов, добавляемых на любой из стадий фурцилий, может варьировать даже в пределах одного вида в зависимости от условий окружающей среды. [52] После последней стадии фурцилии незрелая несовершеннолетняя особь приобретает форму, похожую на взрослую, а затем у нее развиваются гонады и происходит половое созревание. [53]
Воспроизведение
Голова самки криля мешко-нерестового вида Nematoscelis difficilis с выводковым мешком. Яйца имеют диаметр 0,3–0,4 миллиметра (0,012–0,016 дюйма).
Во время брачного сезона, который зависит от вида и климата, самец откладывает сперматозоид в половое отверстие самки (так называемый теликум ). Самки могут нести в яичниках несколько тысяч яиц , что может составлять до трети массы тела животного. [54] Криль может иметь несколько выводков за один сезон, причем интервалы между выводками составляют порядка дней. [24] [55]
Криль использует два типа механизма нереста. [24] 57 видов родов Bentheuphausia , Euphausia , Meganyctiphanes , Thysanoessa и Thysanopoda являются «широковещательными производителями»: самка выпускает оплодотворенные яйца в воду, где они обычно тонут, рассеиваются и остаются одни. Эти виды обычно вылупляются на стадии науплиуса 1, но недавно было обнаружено, что они иногда вылупляются на стадии метанауплиуса или даже на стадии калиптописа. [56] Остальные 29 видов других родов являются «производителями мешков», где самка несет с собой икру, прикрепленную к самым задним парам торакопод, пока они не вылупляются как метанауплии, хотя некоторые виды, такие как Nematoscelis difficilis , могут вылупляться как науплиусы или как науплиусы. псевдометанауплиус. [57]
линька
Линька происходит всякий раз, когда особь перерастает свой жесткий экзоскелет. Молодые животные, растущие быстрее, линяют чаще, чем старшие и крупные. Частота линьки широко варьируется в зависимости от вида и даже в пределах одного вида зависит от многих внешних факторов, таких как широта, температура воды и наличие пищи. Например, у субтропического вида Nyctiphanes simplex общий период между линькой составляет от двух до семи дней: личинки линяют в среднем каждые четыре дня, а молодые и взрослые особи - в среднем каждые шесть дней. Для E. superba в антарктическом море наблюдались периоды между линькой от 9 до 28 дней в зависимости от температуры от -1 до 4 °C (от 30 до 39 °F), а у Meganyctiphanes norvegica в Северном море периоды между линьками также варьируются от 9 до 28 дней, но при температуре от 2,5 до 15 ° C (от 36,5 до 59,0 ° F). [58] E. superba способен уменьшать размер своего тела, когда не хватает пищи, а также линяет, когда его экзоскелет становится слишком большим. [59] Подобное сокращение также наблюдалось для E. pacifica , вида, встречающегося в Тихом океане от полярных до умеренных зон, как адаптация к аномально высоким температурам воды. Предполагается, что сокращение численности произойдет и для других видов криля умеренного пояса. [60]
Продолжительность жизни
Некоторые высокоширотные виды криля могут жить более шести лет (например, Euphausia superba ); другие, такие как вид средних широт Euphausia pacifica , живут всего два года. [6] Продолжительность жизни субтропических или тропических видов еще короче, например, Nyctiphanes simplex , который обычно живет всего шесть-восемь месяцев. [61]
Роение
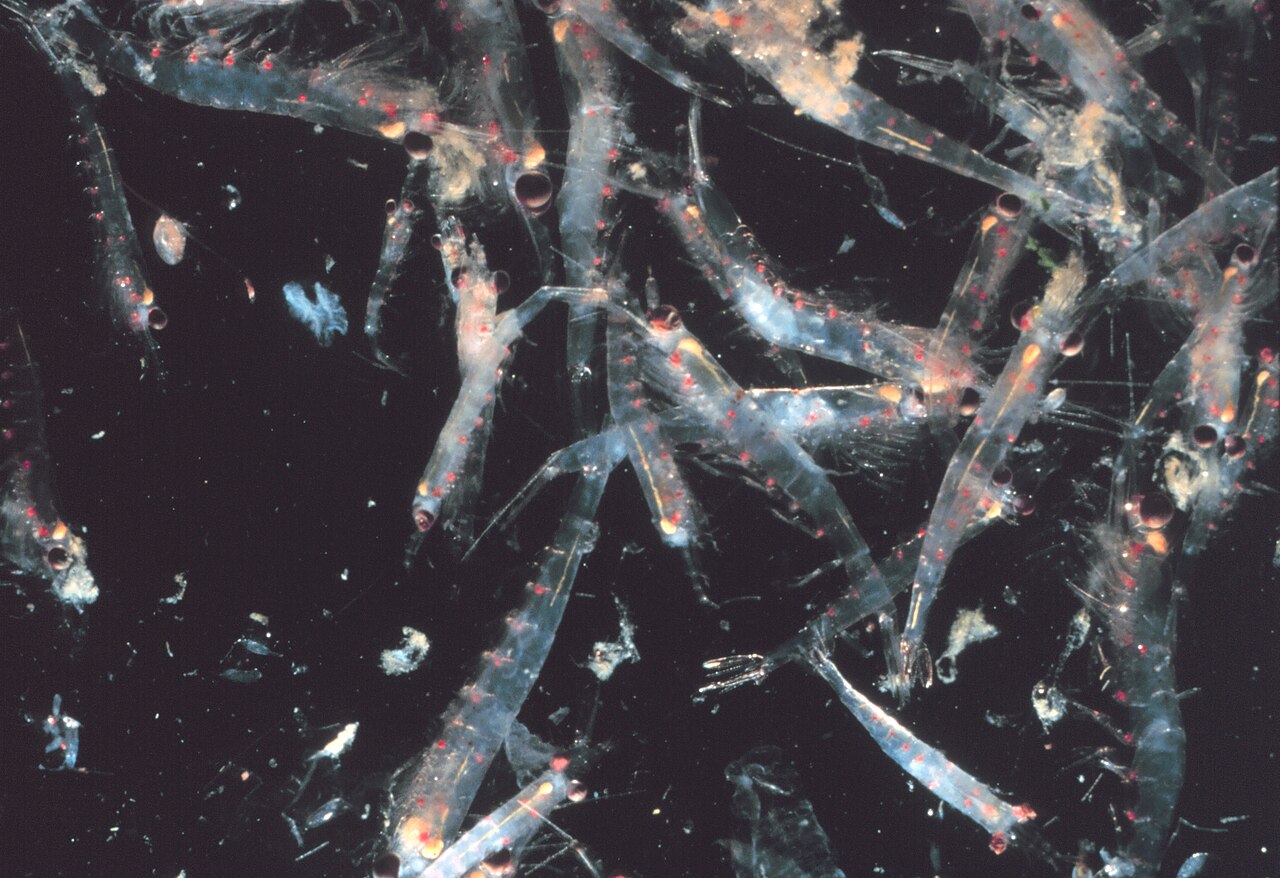
Рой криля
Большинство криля — роящиеся животные; размеры и плотность таких стай различаются в зависимости от вида и региона. У Euphausia superba стаи достигают от 10 000 до 60 000 особей на кубический метр. [62] [63] Роение — это защитный механизм, сбивающий с толку более мелких хищников, которые хотели бы выделить особей. В 2012 году Гандоми и Алави представили, по всей видимости, успешный стохастический алгоритм для моделирования поведения стаев криля. Алгоритм основан на трех основных факторах: «(i) движение, вызванное присутствием других особей, (ii) деятельность по поиску пищи и (iii) случайное распространение». [64]
Криль обычно совершает суточную вертикальную миграцию . Предполагалось, что день они проводят на большей глубине, а ночью поднимаются к поверхности. Чем глубже они заходят, тем больше снижают свою активность [65] , по-видимому, для уменьшения встреч с хищниками и для сохранения энергии. Плавательная активность криля зависит от наполненности желудка. Сытые животные, питавшиеся у поверхности, плавают менее активно и поэтому опускаются ниже перемешанного слоя. [66] Когда они тонут, они производят фекалии , что подразумевает их роль в антарктическом углеродном цикле . Криль с пустым желудком плавает активнее и направляется к поверхности.
Вертикальная миграция может происходить 2–3 раза в день. Некоторые виды (например, Euphausia superba , E. pacifica , E. hanseni , Pseudeuphausia latifrons и Thysanoessa spinifera ) в течение дня образуют наземные стаи для кормления и размножения, хотя такое поведение опасно, поскольку делает их чрезвычайно уязвимыми для хищников. [67]
Экспериментальные исследования с использованием Artemia salina в качестве модели показывают, что вертикальные миграции криля на несколько сотен метров, группами глубиной в десятки метров, могут в совокупности создавать достаточное количество нисходящих струй воды, чтобы оказать существенное влияние на перемешивание океана. [68]
Плотные стаи могут спровоцировать безумную жажду кормления среди рыб, птиц и хищников-млекопитающих, особенно у поверхности. Если их потревожить, рой разбегается, а некоторые особи даже мгновенно линяют, оставляя экзувию в качестве приманки. [69]
Криль обычно плавает со скоростью 5–10 см/с (2–3 длины тела в секунду), [70] используя свои плавальцы для движения. Их более крупные миграции зависят от океанских течений. В случае опасности они демонстрируют реакцию бегства , называемую омаром : щелкая хвостовыми структурами, тельсонами и уроподами , они относительно быстро движутся назад по воде, достигая скорости в диапазоне от 10 до 27 длин тела в секунду, что для крупного криля например, E. superba , означает около 0,8 м/с (3 фута/с). [71] Их навыки плавания побудили многих исследователей классифицировать взрослых особей криля как микронектонные формы жизни, т.е. мелких животных, способных к индивидуальному движению против (слабых) течений. Личиночные формы криля обычно считаются зоопланктоном. [72]
Биогеохимические циклы
Роль антарктического криля в биогеохимических циклах
Криль (как стаи, так и отдельные особи) питается фитопланктоном на поверхности (1), оставляя лишь часть его тонуть в виде фитодетритных агрегатов (2), которые легко распадаются и не могут опускаться ниже постоянного термоклина. Криль также выделяет фекальные гранулы (3) во время питания, которые могут опускаться на глубину моря, но могут быть съедены (копрофагия) и разложены по мере спуска (4) крилем, бактериями и зоопланктоном. В краевой зоне льда поток фекальных частиц может достигать больших глубин (5). Криль также выпускает линьки, которые тонут и способствуют выбросу углерода (6). Питательные вещества, такие как железо и аммоний, высвобождаются крилем во время неаккуратного питания, выделения и пищеварения (7, другие высвобождаемые питательные вещества см. на рис. 2), и если они высвобождаются вблизи поверхности, они могут стимулировать выработку фитопланктона и дальнейшее сокращение атмосферного CO2. Некоторые взрослые особи криля постоянно обитают глубже в толще воды, потребляя органический материал на глубине (8). Любой углерод (в виде органического вещества или CO2), который опускается ниже постоянного термоклина, не подвергается сезонному перемешиванию и будет храниться в глубинах океана в течение как минимум года (9). Плавательные движения мигрирующих взрослых особей криля могут смешивать богатую питательными веществами воду с глубины (10), что дополнительно стимулирует первичную продукцию. Другой взрослый криль является кормом на морском дне, выделяя вдыхаемый CO2 на глубине и может поедаться донными хищниками (11). Личинки криля, которые в Южном океане обитают подо льдом, подвергаются обширной суточной вертикальной миграции (12), потенциально перенося CO2 ниже постоянного термоклина. Криль потребляется многими хищниками, включая усатых китов (13), что приводит к хранению некоторой части углерода криля в виде биомассы в течение десятилетий, прежде чем кит умрет, опустится на морское дно и будет съеден глубоководными организмами. [41]
Во время линьки криля они выделяют растворенный кальций, фторид и фосфор из экзоскелета (1). Хитин (органический материал), образующий экзоскелет, способствует попаданию потока органических частиц в глубины океана. Криль вдыхает часть энергии, полученной в результате потребления фитопланктона или других животных в виде углекислого газа (2), при плавании из средних/глубоких вод на поверхность в больших стаях криля, смешивая воду, которая потенциально доставляет питательные вещества в бедные питательными веществами поверхностные воды (3 ), аммоний и фосфат выделяются из жабр и при выделении вместе с растворенным органическим углеродом азот (например, мочевина) и фосфор (ДОУ, ДОН и ДОФ, 2 и 4). Криль выделяет быстро тонущие фекальные гранулы, содержащие твердые частицы органического углерода, азота и фосфора (POC, PON и POP) и железа, последнее из которых биодоступно при выщелачивании в окружающие воды вместе с DOC, DON и DOP (5). [41]
Человеческое использование
Пластины глубокой заморозки антарктического криля для использования в качестве корма для животных и сырья для приготовления пищи.
История сбора урожая
Криль собирали в качестве источника пищи для людей и домашних животных, по крайней мере, с 19 века, а возможно, и раньше, в Японии, где он был известен как окиами . Крупномасштабный промысел развился в конце 1960-х — начале 1970-х годов и сейчас ведется только в антарктических водах и морях вокруг Японии. Исторически крупнейшими странами, занимающимися промыслом криля, были Япония и Советский Союз, а после распада последнего — Россия и Украина . [76] Пик вылова, который в 1983 году составил около 528 000 тонн только в Южном океане (из которых Советский Союз забрал 93%), теперь управляется в качестве меры предосторожности против чрезмерного вылова рыбы. [77]
В 1993 году два события привели к сокращению вылова криля: Россия вышла из отрасли; а Конвенция о сохранении морских живых ресурсов Антарктики (АНТКОМ) определила максимальные квоты на вылов для устойчивой добычи антарктического криля. После пересмотра в октябре 2011 года Комиссия решила не менять квоту. [78]
Ежегодный вылов в Антарктике стабилизировался на уровне около 100 000 тонн, что составляет примерно одну пятидесятую квоты АНТКОМа на вылов. [79] Основным сдерживающим фактором, вероятно, были высокие затраты, а также политические и юридические проблемы. [80] Японский промысел насыщен примерно на 70 000 тонн. [81]
Хотя криль встречается по всему миру, предпочтительнее ловить рыбу в Южных океанах, поскольку в этих регионах криль более «уловист» и многочисленн. В частности, в антарктических морях, которые считаются нетронутыми, они считаются «чистым продуктом». [76]
В 2018 году было объявлено, что почти все компании по добыче криля, работающие в Антарктиде, с 2020 года откажутся от деятельности на огромных территориях вокруг Антарктического полуострова, включая «буферные зоны» вокруг гнездовых колоний пингвинов. [82]
Хотя общая биомасса антарктического криля может достигать 400 миллионов тонн , антропогенное воздействие на этот ключевой вид растет: за 2010–2014 годы общий вылов рыбы увеличился на 39% до 294 000 тонн. [79] Основными странами, участвующими в добыче криля, являются Норвегия (56% общего вылова в 2014 г.), Республика Корея (19%) и Китай (18%). [79]
Криль является богатым источником белка и жирных кислот омега-3 , которые в начале 21 века разрабатываются в качестве продуктов питания для человека, пищевых добавок в виде масляных капсул, корма для скота и корма для домашних животных . [76] [78] [83] Криль имеет соленый вкус и несколько более сильный рыбный вкус, чем креветки. Продукты массового потребления и продукты промышленного производства необходимо очистить от кожуры, чтобы удалить несъедобный экзоскелет . [83]
Криль (и другие планктонные креветки , особенно виды Acetes ) наиболее широко потребляются в Юго-Восточной Азии, где его ферментируют (с неповрежденной раковиной) и обычно мелко измельчают для приготовления креветочной пасты . Его можно жарить и есть в сочетании с белым рисом или использовать для придания аромата умами широкому спектру традиционных блюд. [85] [86] Жидкость, образующуюся в процессе ферментации, также собирают в виде рыбного соуса . [87]
Биологическая робототехника
Криль — ловкие пловцы в промежуточном режиме чисел Рейнольдса , в котором не так много решений для беспилотной подводной робототехники, и он вдохновил роботизированные платформы как на изучение их передвижения, так и на поиск конструктивных решений для подводных роботов. [88]
^ "Криль". Интернет-словарь этимологии . Проверено 22 июня 2010 г.
^ А. Аткинсон; В. Сигел; Е.А. Пахомов; М.Дж. Джессопп; В. Леб (2009). «Переоценка общей биомассы и годового производства антарктического криля» (PDF) . Глубоководные исследования . Часть I. 56 (5): 727–740. Бибкод : 2009DSRI...56..727A. дои : 10.1016/j.dsr.2008.12.007.
^ Волкер Сигел (2011). Сигел В. (ред.). «Euphausiidae Дана, 1852». Мировая база данных Euphausiacea . Всемирный реестр морских видов . Проверено 25 ноября 2011 г.
^ аб С. Никол; Ю. Эндо (1999). «Промысел криля: развитие, управление и последствия для экосистемы». Водные живые ресурсы . 12 (2): 105–120. дои : 10.1016/S0990-7440(99)80020-5. S2CID 84158071.
^ abcd Андреас Маас; Дитер Валошек (2001). «Развитие личинок Euphausia superba Dana, 1852 и филогенетический анализ Euphausiacea» (PDF) . Гидробиология . 448 : 143–169. дои : 10.1023/А: 1017549321961. S2CID 32997380. Архивировано из оригинала (PDF) 18 июля 2011 года.
^ Бернадетт Казанова (1984). «Phylogénie des Euphausiacés (Crustacés Eucarides)» [Филогения Euphausiacea (Crustacea: Eucarida)]. Бюллетень Национального музея естественной истории (на французском языке). 4 : 1077–1089.
^ М. Евгения Д'Амато; Гордон В. Харкинс; Тулио де Оливейра; Питер Р. Теске; Марк Дж. Гиббонс (2008). «Молекулярное датирование и биогеография неритического криля Nyctiphanes» (PDF) . Морская биология . 155 (2): 243–247. дои : 10.1007/s00227-008-1005-0. S2CID 17750015. Архивировано из оригинала (PDF) 17 марта 2012 года . Проверено 4 июля 2010 г.
^ abc Саймон Н. Джарман (2001). «Эволюционная история криля, выведенная на основе анализа последовательности ядерной большой субъединицы рДНК». Биологический журнал Линнеевского общества . 73 (2): 199–212. дои : 10.1111/j.1095-8312.2001.tb01357.x .
^ Йохан Эрик Вести Боас (1883). «Studien über die Verwandtschaftsbeziehungen der Malacostracen» [Исследования взаимоотношений Малакострака]. Morphologisches Jahrbuch (на немецком языке). 8 : 485–579.
^ Триша Спирс, Рональд В. ДеБри, Лоуренс Г. Абеле и Катажина Чодил (2005). Бойко, Кристофер Б. (ред.). «Монофилия перакарид и межпорядковая филогения, выведенная из последовательностей ядерной рибосомальной ДНК малых субъединиц (Crustacea: Malacostraca: Peracarida)» (PDF) . Труды Биологического общества Вашингтона . 118 (1): 117–157. doi :10.2988/0006-324X(2005)118[117:PMAIPI]2.0.CO;2. S2CID 85557065.{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )
^ К. Меланд; Э. Уиллассен (2007). «Разобщенность «Мизидацеи» (Рачка)». Молекулярная филогенетика и эволюция . 44 (3): 1083–1104. CiteSeerX 10.1.1.653.5935 . doi :10.1016/j.ympev.2007.02.009. ПМИД 17398121.
^ Аб Д'Амато, МЭ и др. : «Молекулярное датирование и биогеография неритического криля Nyctiphanes. Архивировано 17 марта 2012 г. в Wayback Machine », в журнале Marine Biology vol. 155, нет. 2 , стр. 243–247, август 2008 г.
^ Волкер Сигел (2011). В. Сигел (ред.). «Никтифан Сарс, 1883». Мировая база данных Euphausiacea . Всемирный реестр морских видов . Проверено 18 июня 2011 г.
^ abc Хайме Гомес-Гутьеррес; Карлос Дж. Робинсон (2005). «Эмбриональное, раннее время развития личинок, механизм вылупления и межвыводковый период эвфаузиид, нерестящихся в мешочках, Nyctiphanes simplex Hansen». Журнал исследований планктона . 27 (3): 279–295. дои : 10.1093/plankt/fbi003 .
^ С. Н. Джарман; Н.Г. Эллиотт; С. Николь; А. МакМинн (2002). «Генетическая дифференциация антарктического прибрежного криля Euphausia Crystallorophias». Наследственность . 88 (4): 280–287. дои : 10.1038/sj.hdy.6800041 . ПМИД 11920136.
^ Р. Эскрибано; В. Марин; К. Иррибаррен (2000). «Распространение Euphausia mucronata в зоне апвеллинга полуострова Медхильонес, север Чили: влияние минимального слоя кислорода». Сциентия Марина . 64 (1): 69–77. дои : 10.3989/scimar.2000.64n169 .
^ П. Брюггеман. «Эуфаузия кристаллорофиас». Подводный путеводитель по острову Росс и проливу Мак-Мердо, Антарктида . Калифорнийский университет, Сан-Диего .
^ "Криль, Euphausia superba" . MarineBio.org . Проверено 25 февраля 2009 г.
^ Дж. А. Кирквуд (1984). «Путеводитель по эвфаузиацеям Южного океана». Заметки об исследованиях ANARE . 1 :1–45.
^ Бьянки, Даниэле; Мислан, КАС (январь 2016 г.). «Глобальные закономерности суточного времени и скорости вертикальной миграции по акустическим данным». Лимнология и океанография . 61 (1). дои : 10.1002/lno.10219 .
^ А. Сала; М. Аззали; А. Руссо (2002). «Криль моря Росса: распространение, численность и демография Euphausia superba и Euphausia Crystallorophias во время итальянской антарктической экспедиции (январь – февраль 2000 г.)». Сциентия Марина . 66 (2): 123–133. дои : 10.3989/scimar.2002.66n2123 .
^ Г.В. Хози; М. Фукучи; С. Кавагути (2003). «Развитие системы непрерывного наблюдения за планктоном Южного океана» (PDF) . Прогресс в океанографии . 58 (2–4): 263–283. Бибкод : 2003Proce..58..263H. doi :10.1016/j.pocean.2003.08.007.[ постоянная мертвая ссылка ]
^ Э. Гейтен. «Меганиктипанес норвегика». Университет Лестера . Архивировано из оригинала 1 июля 2009 года . Проверено 25 февраля 2009 г.
^ Э. Бринтон (1953). « Thysanopoda spinicauda , новый батипелагический гигантский эвфаузиидный ракообразный, со сравнительными заметками о T. cornuta и T. egregia ». Журнал Вашингтонской академии наук . 43 : 408–412.
^ "Эуфаузиацея". Тасманский институт аквакультуры и рыболовства. Архивировано из оригинала 30 сентября 2009 года . Проверено 6 июня 2010 г.
^ О. Шимомура (1995). «Роль двух крайне нестабильных компонентов F и P, участвующих в биолюминесценции эвфаузиидных креветок». Журнал биолюминесценции и хемилюминесценции . 10 (2): 91–101. дои : 10.1002/bio.1170100205. ПМИД 7676855.
^ Джей Си Данлэп; Дж. У. Гастингс; О. Шимомура (1980). «Кроссреактивность между светоизлучающими системами отдаленно родственных организмов: новый тип светоизлучающих соединений». Труды Национальной академии наук . 77 (3): 1394–1397. Бибкод : 1980PNAS...77.1394D. дои : 10.1073/pnas.77.3.1394 . JSTOR 8463. PMC 348501 . ПМИД 16592787.
^ Пи Джей Херринг; Э.А. Виддер (2001). «Биолюминесценция планктона и нектона». В Дж. Х. Стиле; С.А. Торп; К.К. Турекян (ред.). Энциклопедия наук об океане . Том. 1. Академик Пресс , Сан-Диего. стр. 308–317. ISBN978-0-12-227430-5.
^ С.М. Линдси; М. И. Латц (1999). Экспериментальные доказательства люминесцентного затенения некоторых эвфаузиидных ракообразных . Собрание Американского общества лимнологии и океанографии (ASLO) по водным наукам. Санта-Фе.
^ Сёнке Джонсен (2005). «Красное и черное: биолюминесценция и окраска животных морских глубин» (PDF) . Интегративная и сравнительная биология . 4 (2): 234–246. дои : 10.1093/icb/45.2.234 . PMID 21676767. S2CID 247718. Архивировано из оригинала (PDF) 2 октября 2005 г.
^ abcd Каван, Э.Л., Белчер, А., Аткинсон, А., Хилл, С.Л., Кавагути, С., МакКормак, С., Мейер, Б., Никол, С., Ратнараджа, Л., Шмидт, К. и Стейнберг, Д.К. (2019) «Важность антарктического криля в биогеохимических циклах». Природные коммуникации , 10 (1): 1–13. дои : 10.1038/s41467-019-12668-7.Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0.
^ GC Криппс; А. Аткинсон (2000). «Жирнокислотный состав как показатель плотоядности антарктического криля Euphausia superba ». Канадский журнал рыболовства и водных наук . 57 (С3): 31–37. дои : 10.1139/f00-167.
^ аб Олав Сэтер; Тронд Эрлинг Эллингсен; Вигго Мор (1986). «Липиды североатлантического криля» (PDF) . Журнал исследований липидов . 27 (3): 274–285. ПМИД 3734626.
^ MJ Шрамм (10 октября 2007 г.). «Крошечный криль: гиганты в морской пищевой цепи». Национальная программа морских заповедников NOAA . Проверено 4 июня 2010 г.
^ Дж. Вейер (1999). «Изменяющиеся течения окрашивают Берингово море в новый оттенок синего». Земная обсерватория НОАА . Проверено 15 июня 2005 г.
^ Р. Д. Бродер; Г.Х. Крузе; П.А. Ливингстон; Г. Уолтерс; Дж. Янелли; Г.Л. Шварцман; М. Степаненко; Т. Уилли-Эчеверрия (1998). Проект отчета Международного семинара FOCI по последним условиям в Беринговом море . НОАА . стр. 22–26.
^ Дж. Роуч (17 июля 2003 г.). «Ученые обнаружили загадочного убийцу криля» . Национальные географические новости . Архивировано из оригинала 24 июля 2003 года.
^ Х. Гомес-Гутьеррес; У. Т. Петерсон; А. де Робертис; Р.Д. Бродер (2003). «Массовая смертность криля, вызванная паразитоидными инфузориями». Наука . 301 (5631): 339. doi :10.1126/science.1085164. PMID 12869754. S2CID 28471713.
^ Джей Ди Шилдс; Х. Гомес-Гутьеррес (1996). « Oculophryxus bicaulis , новый род и вид изопод даджидов, паразитирующих на эвфаузиид Stylocheiron affine Hansen». Международный журнал паразитологии . 26 (3): 261–268. дои : 10.1016/0020-7519(95)00126-3. ПМИД 8786215.
↑ Расти Дорнин (6 июля 1997 г.). «Популяция антарктического криля сокращается». CNN . Проверено 18 июня 2011 г.
^ Доктор медицины Найт (1984). «Изменения в морфогенезе личинок в популяции Euphausia pacifica в заливе Южной Калифорнии с зимы по лето 1977–1978 годов» (PDF) . Отчет CalCOFI . XXV . Архивировано из оригинала (PDF) 3 августа 2019 года . Проверено 5 ноября 2017 г.
^ Р. М. Росс; Л. Б. Кетин (1986). «Насколько продуктивен антарктический криль?». Бионаука . 36 (4): 264–269. дои : 10.2307/1310217. JSTOR 1310217.
^ Джанин Кузен-Руди (2000). «Сезонное размножение, множественный нерест и плодовитость северного криля Meganyctiphanes norvegica и антарктического криля Euphausia superba ». Канадский журнал рыболовства и водных наук . 57 (С3): 6–15. дои : 10.1139/f00-165.
^ Дж. Гомес-Гутьеррес (2002). «Механизм вылупления и замедленное вылупление икры трех широконерестящихся видов эвфаузиид в лабораторных условиях». Журнал исследований планктона . 24 (12): 1265–1276. дои : 10.1093/планкт/24.12.1265 .
^ Э. Бринтон; доктор медицинских наук Оман; А. В. Таунсенд; доктор медицины Найт; А. Л. Бриджман (2000). Эуфаузииды Мирового океана. Серия компакт-дисков с Всемирной базой данных по биоразнообразию, Springer Verlag . ISBN978-3-540-14673-5. Архивировано из оригинала 26 февраля 2012 года . Проверено 4 декабря 2009 г.
^ Ф. Бухгольц (2003). «Опыты по физиологии южного и северного криля, Euphausia superba и Meganyctiphanes norvegica с акцентом на линьку и рост – обзор». Морское и пресноводное поведение и физиология . 36 (4): 229–247. Бибкод : 2003MFBP...36..229B. дои : 10.1080/10236240310001623376. S2CID 85121989.
^ Х.-К. Шин; С. Никол (2002). «Использование взаимосвязи между диаметром глаза и длиной тела для выявления последствий длительного голодания антарктического криля Euphausia superba». Серия «Прогресс в области морской экологии ». 239 : 157–167. Бибкод : 2002MEPS..239..157S. дои : 10.3354/meps239157 .
^ Б. Маринович; М. Мангель (1999). «Криль может сокращаться в результате экологической адаптации к временно неблагоприятным условиям» (PDF) . Экологические письма . 2 : 338–343.
^ Дж. Г. Гомес (1995). «Схемы распространения, численность и динамика популяций эвфаузиид Nyctiphanes simplex и Euphausia eximia у западного побережья Нижней Калифорнии, Мексика» (PDF) . Серия «Прогресс в области морской экологии ». 119 : 63–76. Бибкод : 1995MEPS..119...63G. дои : 10.3354/meps119063 .
^ У. Килс; П. Маршалл (1995). «Der Krill, wie er schwimmt und frisst – neue Einsichten mit neuen Methoden (« Антарктический криль – как он плавает и питается – новые идеи с новыми методами »)». У И. Гемпеля; Г. Хемпель (ред.). Biologie der Polarmeere – Erlebnisse und Ergebnisse ( Опыт и результаты биологии полярных океанов ) . Фишер Верлаг . стр. 201–210. ISBN978-3-334-60950-7.
^ Р. Пайпер (2007). Необыкновенные животные: Энциклопедия любопытных и необычных животных . Гринвуд Пресс . ISBN978-0-313-33922-6.
^ Гандоми, АХ; Алави, АХ (2012). «Стадо криля: новый алгоритм оптимизации, основанный на биотехнологиях». Коммуникации в нелинейной науке и численном моделировании . 17 (12): 4831–4845. Бибкод : 2012CNSNS..17.4831G. doi :10.1016/j.cnsns.2012.05.010.
^ Дж. С. Яффе; доктор медицинских наук Оманн; А. де Робертис (1999). «Сонарные оценки уровней дневной активности Euphausia pacifica в заливе Саанич» (PDF) . Канадский журнал рыболовства и водных наук . 56 (11): 2000–2010. doi : 10.1139/cjfas-56-11-2000. S2CID 228567512. Архивировано из оригинала (PDF) 20 июля 2011 года.
^ Герайнт А. Тарлинг; Магнус Л. Джонсон (2006). «Насыщение вызывает у криля ощущение погружения». Современная биология . 16 (3): 83–84. дои : 10.1016/j.cub.2006.01.044 . ПМИД 16461267.
^ Дэн Ховард (2001). «Криль» (PDF) . У Германа А. Карла; Джон Л. Чин; Эдвард Юбер; Питер Х. Стауффер; Джеймс В. Хендли II (ред.). За Золотыми воротами – океанография, геология, биология и проблемы окружающей среды в заливе Фараллонс. Геологическая служба США . стр. 133–140. Циркуляр 1198 . Проверено 8 октября 2011 г.
^ Уишарт, Скай (июль – август 2018 г.). «Эффект криля». География Новой Зеландии (152): 24.
^ Дэвид А. Демер; Стефан Г. Конти (2005). «Новая модель целевой силы указывает на увеличение количества криля в Южном океане». Журнал морских наук ICES . 62 (1): 25–32. дои : 10.1016/j.icesjms.2004.07.027 .
^ У. Килс (1982). «Плавательное поведение, плавательные способности и энергетический баланс антарктического криля Euphausia superba». Научная серия БИОМАССА 3, Серия исследований БИОМАССЫ : 1–122. Архивировано из оригинала 2 июня 2020 года . Проверено 11 ноября 2017 г.
^ С. Никол; Ю. Эндо (1997). «Мировой промысел криля». Технический документ ФАО по рыболовству . 367 .
^ Ратнараджа, Л., Боуи, А.Р., Ланнузель, Д., Майнерс, К.М. и Никол, С. (2014) «Биогеохимическая роль усатых китов и криля в круговороте питательных веществ в Южном океане». ПЛОС ОДИН , 9 (12): е114067. дои :10.1371/journal.pone.0114067
^ Хопкинс, Т.Л., Эйнли, Д.Г., Торрес, Дж.Дж., Ланкрафт, Т.М., 1993. Трофическая структура в открытых водах краевой ледниковой зоны в районе слияния Скотия-Уэдделл весной (1983). Полярная биология 13, 389–397.
^ Ланкрафт, Т.М., Релсенбихлер, К.Р., Робинсон, Б.Х., Хопкинс, Т.Л., Торрес, Дж.Дж., 2004. Сообщество микронектона и макрозоопланктона с преобладанием криля в проливе Крокер, Антарктида, с оценкой хищничества рыб. Глубоководные исследования II 51, 2247–2260.
↑ abc Гроссман, Элизабет (14 июля 2015 г.). «Ученые думают, нужно ли защищать криль от чрезмерной охоты со стороны человека». Международное общественное радио (PRI) . Проверено 1 апреля 2017 г.
^ «Промысел криля и устойчивость: антарктический криль (Euphausia superba)» . Комиссия по сохранению морских живых ресурсов Антарктики. 23 апреля 2015 года . Проверено 1 апреля 2017 г.
^ abc «Криль – биология, экология и рыболовство». Комиссия по сохранению морских живых ресурсов Антарктики. 28 апреля 2015 года . Проверено 1 апреля 2017 г.
^ Минтурн Дж. Райт (1987). «Собственность Антарктиды, ее живые и минеральные ресурсы». Журнал права и окружающей среды . 4 (2): 49–78.
^ С. Никол; Дж. Фостер (2003). «Последние тенденции в промысле антарктического криля». Водные живые ресурсы . 16 : 42–45. дои : 10.1016/S0990-7440(03)00004-4.
^ Джош, Габбатисс (10 июля 2018 г.). «Индустрия лова криля поддерживает огромный заповедник в Антарктическом океане для защиты пингвинов, тюленей и китов». Независимый . Проверено 10 июля 2018 г.
^ ab "Почему криль?". Юго-западный научный центр рыболовства, Национальное управление океанических и атмосферных исследований США. 22 ноября 2016 года . Проверено 1 апреля 2017 г.
↑ Чизмен, Массачусетс (22 июля 2011 г.). «Масло криля: ответное письмо агентства, уведомление GRAS № GRN 000371». FDA США . Проверено 3 июня 2015 г.
^ Омори, М. (1978). «Мировой промысел зоопланктона: обзор». Морская биология . 48 (3): 199–205. дои : 10.1007/BF00397145. S2CID 86540101.
^ Понгсеткул, Джаксума; Бенджакул, Соттават; Сампавапол, Пеннани; Осако, Кадзуфуми; Фэйтхонг, Нандша (17 сентября 2014 г.). «Химический состав и физические свойства пасты из соленых креветок (Капи) производства Таиланда». Международные водные исследования . 6 (3): 155–166. дои : 10.1007/s40071-014-0076-4 .
^ Абэ, Кенджи; Сузуки, Кенджи; Хасимото, Канехиса (1979). «Использование криля в качестве материала для рыбного соуса». Ниппон Суйсан Гаккаиси . 45 (8): 1013–1017. дои : 10.2331/suisan.45.1013 .
^ Оливейра Сантос, Сара; Так, Нильс; Су, Юньсин; Куэнка-Хименес, Франциско; Моралес-Лопес, Оскар; Гомес-Вальдес, П. Антонио; М. Вильгельмус, Моника (13 июня 2023 г.). «Плеобот: модульное роботизированное решение для метахронного плавания». Научные отчеты . 13 (1): 9574. Бибкод : 2023NatSR..13.9574O. дои : 10.1038/s41598-023-36185-2. ПМЦ 10264458 . ПМИД 37311777.
дальнейшее чтение
Боден, Брайан П.; Джонсон, Мартин В .; Бринтон, Эдвард: «Эуфаузиацея (Рачки) северной части Тихого океана». Бюллетень Океанографического института Скриппса . Том 6, номер 8, 1955 год.
Бринтон, Эдвард : «Эуфаузииды вод Юго-Восточной Азии». Отчет Наги, том 4, часть 5. Ла-Хойя: Калифорнийский университет, Океанографический институт Скриппса, 1975.
Конвей, главный вице-президент; Уайт, Р.Г.; Хьюг-Дит-Сайлс, Ж.; Галиен, CP; Робинс, Д.Б.: Путеводитель по прибрежному и поверхностному зоопланктону юго-западной части Индийского океана. Архивировано 23 октября 2012 г. в Wayback Machine , Order Euphausiacea, Периодическая публикация Ассоциации морской биологии Соединенного Королевства № 15, Плимут, Великобритания, 2003 г. .
Эверсон, И. (ред.): Криль: биология, экология и рыболовство . Оксфорд, Блэквелл Сайенс; 2000. ISBN 0-632-05565-0 .
Хамнер, Уильям М. (май 1984 г.). «Криль — неиспользованный дар моря?». Национальная география . Том. 165, нет. 5. стр. 626–642. ISSN 0027-9358. ОКЛК 643483454.
Мочлин, Дж.: Euphausiacea: взрослые особи. Архивировано 15 мая 2011 г. в Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Идентификационные листы взрослых особей криля со множеством линейных рисунков. PDF- файл, 2 Мб .
Мочлин, Дж.: Euphausiacea: Личинки. Архивировано 19 апреля 2012 г. в Wayback Machine , Conseil International pour l'Exploration de la Mer, 1971. Идентификационные листы личиночных стадий криля со множеством линейных рисунков. PDF-файл, 3 Мб.
Тетт, П.: Биология эвфаузиид , конспекты лекций из курса морской биологии 2003 года, проведенного в Университете Нейпира .
Тетт, П.: Биолюминесценция , конспекты лекций из издания того же курса 1999/2000 гг.
Внешние ссылки
Викискладе есть медиафайлы, связанные с крилем.
Wikispecies содержит информацию, связанную с Евфаузией .
Найдите криль в Викисловаре, бесплатном словаре.
Веб-камера крилевого аквариума Австралийского антарктического отдела










{kind=link}