Правила, по которым информация, закодированная в генетическом материале, транслируется в белки
Серия кодонов в части молекулы информационной РНК (мРНК). Каждый кодон состоит из трех нуклеотидов , обычно соответствующих одной аминокислоте . Нуклеотиды сокращенно обозначаются буквами A, U, G и C. Это мРНК, которая использует U ( урацил ). ДНК использует вместо этого T ( тимин ). Эта молекула мРНК даст указание рибосоме синтезировать белок в соответствии с этим кодом.
Генетический код — это набор правил, используемых живыми клетками для перевода информации, закодированной в генетическом материале ( последовательности ДНК или РНК нуклеотидных триплетов, или кодонов ), в белки . Перевод осуществляется рибосомой , которая связывает протеиногенные аминокислоты в порядке, указанном информационной РНК (мРНК), используя молекулы транспортной РНК (тРНК) для переноса аминокислот и считывания мРНК по три нуклеотида за раз. Генетический код очень похож у всех организмов и может быть выражен в простой таблице с 64 записями.
Кодоны определяют, какая аминокислота будет добавлена следующей во время биосинтеза белка . За некоторыми исключениями, [1] трехнуклеотидный кодон в последовательности нуклеиновой кислоты определяет одну аминокислоту. Подавляющее большинство генов кодируется с помощью одной схемы (см. таблицу кодонов РНК ). Эту схему часто называют каноническим или стандартным генетическим кодом или просто генетическим кодом, хотя существуют и вариантные коды (например, в митохондриях ).
История
Генетический код
Попытки понять, как кодируются белки, начались после открытия структуры ДНК в 1953 году. Ключевые первооткрыватели, английский биофизик Фрэнсис Крик и американский биолог Джеймс Уотсон , работая вместе в Кавендишской лаборатории Кембриджского университета, выдвинули гипотезу, что информация исходит от ДНК и что существует связь между ДНК и белками. [2] Советско-американский физик Георгий Гамов был первым, кто дал работающую схему синтеза белка из ДНК. [3] Он постулировал, что наборы из трех оснований (триплеты) должны использоваться для кодирования 20 стандартных аминокислот, используемых живыми клетками для построения белков, что позволило бы использовать максимум 4 3 = 64 аминокислоты. [4] Он назвал это взаимодействие ДНК-белок (исходный генетический код) «алмазным кодом». [5]
В 1954 году Гамов создал неформальную научную организацию RNA Tie Club , как и предложил Уотсон, для ученых разных убеждений, которые интересовались тем, как синтезируются белки из генов. Однако клуб мог иметь только 20 постоянных членов, представляющих каждую из 20 аминокислот; и четырех дополнительных почетных членов, представляющих четыре нуклеотида ДНК. [6]
Первый научный вклад клуба, позже отмеченный как «одна из самых важных неопубликованных статей в истории науки» [7] и «самая известная неопубликованная статья в анналах молекулярной биологии» [8] , был сделан Криком. Крик представил машинописную статью под названием «О вырожденных шаблонах и гипотезе адаптера: заметка для Клуба РНК-связей» [9] членам клуба в январе 1955 года, которая «полностью изменила то, как мы думали о синтезе белка», как вспоминал Уотсон. [10] Гипотеза утверждает, что триплетный код не был передан аминокислотам, как думал Гамов, а переносился другой молекулой, адаптером, который взаимодействует с аминокислотами. [8] Адаптер был позже идентифицирован как тРНК. [11]
За этим последовали эксперименты в лаборатории Северо Очоа , которые продемонстрировали, что последовательность полиадениновой РНК (AAAAA...) кодирует полипептид полилизин [ 14] и что последовательность полицитозиновой РНК (CCCCC...) кодирует полипептид полипролин . [ 15] Таким образом, кодон AAA определяет аминокислоту лизин , а кодон CCC определяет аминокислоту пролин . Затем с использованием различных сополимеров были определены большинство оставшихся кодонов.
Последующая работа Хар Гобинда Кораны определила остальную часть генетического кода. Вскоре после этого Роберт В. Холли определил структуру транспортной РНК (тРНК), молекулы-адаптера, которая облегчает процесс перевода РНК в белок. Эта работа была основана на более ранних исследованиях Очоа, что принесло последнему Нобелевскую премию по физиологии и медицине в 1959 году за работу по энзимологии синтеза РНК. [16]
Продолжая эту работу, Ниренберг и Филипп Ледер раскрыли триплетную природу кода и расшифровали его кодоны. В этих экспериментах различные комбинации мРНК пропускались через фильтр, содержащий рибосомы , компоненты клеток, которые транслируют РНК в белок. Уникальные триплеты способствовали связыванию специфических тРНК с рибосомой. Ледер и Ниренберг смогли определить последовательности 54 из 64 кодонов в своих экспериментах. [17] Корана, Холли и Ниренберг получили Нобелевскую премию (1968) за свою работу. [18]
Три стоп-кодона были названы первооткрывателями Ричардом Эпштейном и Чарльзом Стейнбергом. «Янтарь» был назван в честь их друга Харриса Бернстайна, чья фамилия означает «янтарь» на немецком языке. [19] Два других стоп-кодона были названы «охра» и «опал», чтобы сохранить тему «названий цветов».
Расширенные генетические коды (синтетическая биология)
В широкой академической аудитории широко принята концепция эволюции генетического кода от изначального и неоднозначного генетического кода к четко определенному («замороженному») коду с репертуаром из 20 (+2) канонических аминокислот. [20]
Однако существуют различные мнения, концепции, подходы и идеи, что является наилучшим способом его экспериментального изменения. Предлагаются даже модели, которые предсказывают «точки входа» для вторжения синтетических аминокислот в генетический код. [21]
С 2001 года в белки было добавлено 40 неприродных аминокислот путем создания уникального кодона (перекодирования) и соответствующей пары транспортная РНК: аминоацил – тРНК-синтетаза для кодирования их с различными физико-химическими и биологическими свойствами с целью использования в качестве инструмента для исследования структуры и функции белка или для создания новых или улучшенных белков. [22] [23]
Х. Мураками и М. Сисидо расширили некоторые кодоны до четырех и пяти оснований. Стивен А. Беннер построил функциональный 65-й ( in vivo ) кодон. [24]
В 2015 году Н. Будиса , Д. Зёлль и их коллеги сообщили о полной замене всех 20 899 остатков триптофана (кодонов UGG) на неестественный тиенопиррол-аланин в генетическом коде бактерии Escherichia coli . [25]
В 2016 году был создан первый стабильный полусинтетический организм. Это была (одноклеточная) бактерия с двумя синтетическими основаниями (называемые X и Y). Основания пережили деление клеток. [26] [27]
В 2017 году исследователи из Южной Кореи сообщили, что им удалось создать мышь с расширенным генетическим кодом, способную производить белки с неестественными аминокислотами. [28]
В мае 2019 года исследователи сообщили о создании нового штамма "Syn61" бактерии Escherichia coli . Этот штамм имеет полностью синтетический геном, который рефакторирован (все перекрытия расширены), перекодирован (полностью удалено использование трех из 64 кодонов) и дополнительно модифицирован для удаления теперь ненужных тРНК и факторов высвобождения. Он полностью жизнеспособен и растет в 1,6 раза медленнее, чем его аналог дикого типа "MDS42". [29] [30]
Функции
Рамки считывания в последовательности ДНК области митохондриального генома человека, кодирующей гены MT-ATP8 и MT-ATP6 (черным цветом: позиции с 8525 по 8580 в последовательности с номером доступа NC_012920 [31] ). Существует три возможных рамки считывания в прямом направлении 5' → 3', начинающихся с первой (+1), второй (+2) и третьей позиции (+3). Для каждого кодона (квадратные скобки) аминокислота задается митохондриальным кодом позвоночных , либо в рамке +1 для MT-ATP8 (красным цветом), либо в рамке +3 для MT-ATP6 (синим цветом). Ген MT-ATP8 заканчивается стоп-кодоном TAG (красная точка) в рамке +1. Ген MT-ATP6 начинается с кодона ATG (синий круг для аминокислоты M) в рамке +3.
Рамка для чтения
Рамка считывания определяется начальным триплетом нуклеотидов, с которого начинается трансляция. Она задает рамку для ряда последовательных неперекрывающихся кодонов, которая известна как « открытая рамка считывания » (ORF). Например, строка 5'-AAATGAACG-3' (см. рисунок), если считывается с первой позиции, содержит кодоны AAA, TGA и ACG; если считывается со второй позиции, она содержит кодоны AAT и GAA; а если считывается с третьей позиции, она содержит кодоны ATG и AAC. Таким образом, каждая последовательность может быть прочитана в ее направлении 5' → 3' в трех рамках считывания , каждая из которых производит, возможно, отдельную аминокислотную последовательность: в данном примере Lys (K)-Trp (W)-Thr (T), Asn (N)-Glu (E) или Met (M)-Asn (N) соответственно (при трансляции с помощью митохондриального кода позвоночных). Когда ДНК двухцепочечная, определяются шесть возможных рамок считывания : три в прямой ориентации на одной цепи и три в обратной на противоположной цепи. [32] : 330 Рамки кодирования белка определяются стартовым кодоном , обычно первым кодоном AUG (ATG) в последовательности РНК (ДНК).
Три стоп-кодона имеют названия: UAG — янтарный , UGA — опаловый (иногда его также называют умброй ), а UAA — охра . Стоп-кодоны также называются «терминационными» или «бессмысленными» кодонами. Они сигнализируют о высвобождении зарождающегося полипептида из рибосомы, поскольку ни одна родственная тРНК не имеет антикодонов, комплементарных этим стоп-сигналам, что позволяет фактору высвобождения вместо этого связываться с рибосомой. [34]
Эффект мутаций
Примеры заметных мутаций , которые могут возникнуть у людей [35]
В процессе репликации ДНК иногда происходят ошибки в полимеризации второй цепи. Эти ошибки, мутации , могут повлиять на фенотип организма , особенно если они происходят в кодирующей белок последовательности гена. Частота ошибок обычно составляет 1 ошибку на каждые 10–100 миллионов оснований — из-за способности ДНК-полимераз «корректировать» . [36] [37]
Миссенс-мутации и нонсенс-мутации являются примерами точечных мутаций , которые могут вызывать генетические заболевания, такие как серповидноклеточная анемия и талассемия соответственно. [38] [39] [40] Клинически важные миссенс-мутации обычно изменяют свойства кодируемого аминокислотного остатка среди основных, кислых, полярных или неполярных состояний, тогда как нонсенс-мутации приводят к стоп-кодону . [32]
Мутации, которые нарушают последовательность рамки считывания путем инделей ( вставок или делеций ) некратных 3 нуклеотидных оснований, известны как мутации сдвига рамки считывания . Эти мутации обычно приводят к совершенно иной трансляции по сравнению с оригиналом и, вероятно, вызывают считывание стоп-кодона , что усекает белок. [41] Эти мутации могут нарушать функцию белка и, таким образом, редки в последовательностях, кодирующих белок in vivo . Одна из причин, по которой наследование мутаций сдвига рамки является редким, заключается в том, что если транслируемый белок необходим для роста в условиях селективного давления, с которым сталкивается организм, отсутствие функционального белка может привести к смерти до того, как организм станет жизнеспособным. [42] Мутации сдвига рамки считывания могут приводить к тяжелым генетическим заболеваниям, таким как болезнь Тея-Сакса . [43]
Хотя большинство мутаций, которые изменяют последовательности белков, вредны или нейтральны, некоторые мутации имеют преимущества. [44] Эти мутации могут позволить мутантному организму лучше выдерживать определенные экологические стрессы, чем организмы дикого типа , или размножаться быстрее. В этих случаях мутация будет иметь тенденцию становиться более распространенной в популяции посредством естественного отбора . [45] Вирусы , которые используют РНК в качестве своего генетического материала, имеют быструю скорость мутаций, [46] что может быть преимуществом, поскольку эти вирусы тем самым быстро эволюционируют и, таким образом, избегают защитных реакций иммунной системы . [47] В больших популяциях бесполо размножающихся организмов, например, E. coli , могут одновременно возникать множественные полезные мутации. Это явление называется клональной интерференцией и вызывает конкуренцию между мутациями. [48]
Вырождение
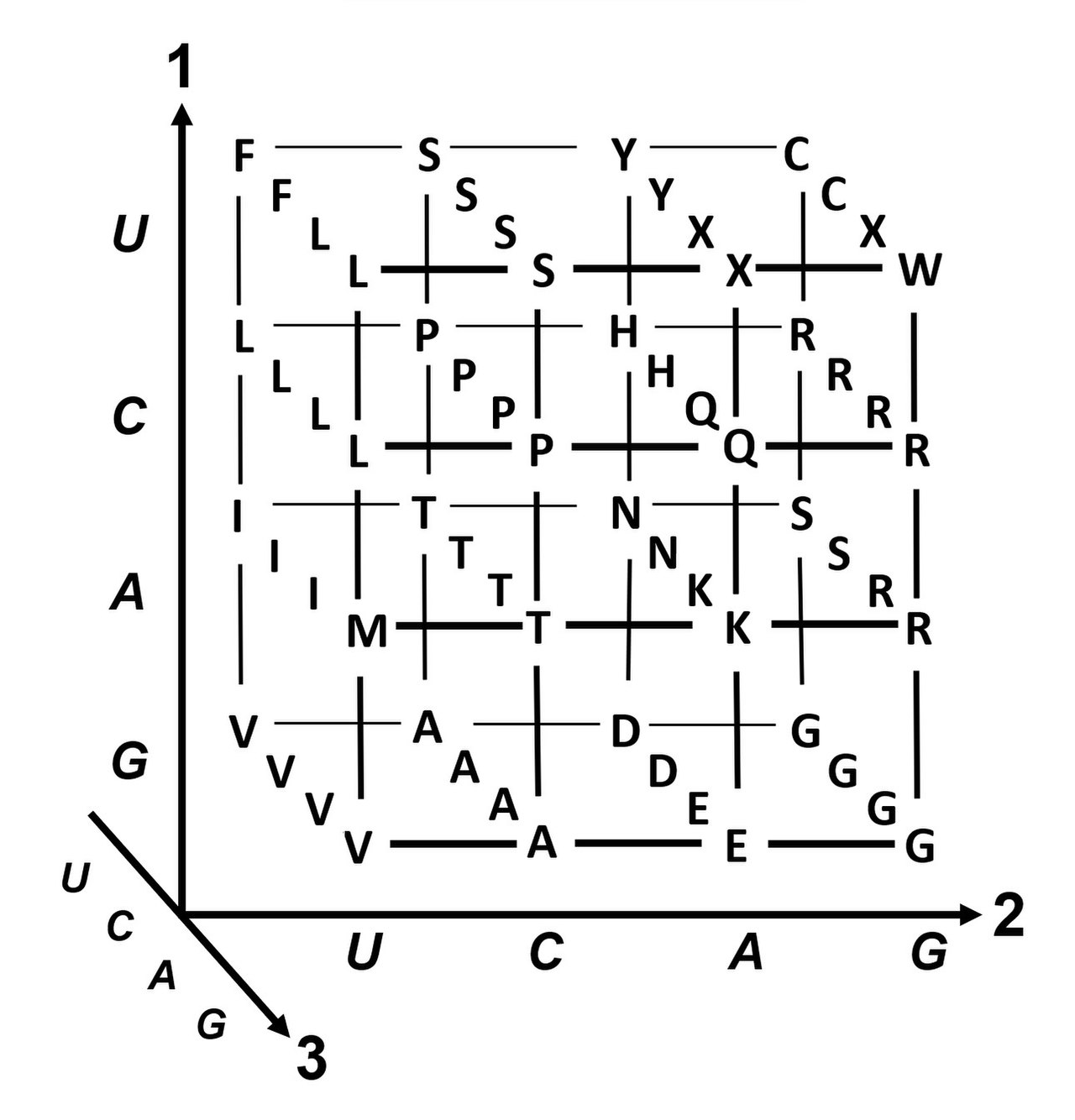
Группировка кодонов по молярному объему аминокислотных остатков и гидропатичности . Доступна более подробная версия .Оси 1, 2, 3 — это первая, вторая и третья позиции в кодоне. 20 аминокислот и стоп-кодоны (X) показаны в однобуквенном коде .
Вырожденность — это избыточность генетического кода. Этот термин был введен Бернфилдом и Ниренбергом. Генетический код обладает избыточностью, но не двусмысленностью (см. таблицы кодонов ниже для полной корреляции). Например, хотя кодоны GAA и GAG оба определяют глутаминовую кислоту (избыточность), ни один из них не определяет другую аминокислоту (не двусмысленность). Кодоны, кодирующие одну аминокислоту, могут отличаться в любой из трех своих позиций. Например, аминокислота лейцин определяется кодонами Y U R или CU N (UUA, UUG, CUU, CUC, CUA или CUG) (разница в первой или третьей позиции указывается с использованием нотации ИЮПАК ), тогда как аминокислота серин определяется кодонами UC N или AG Y (UCA, UCG, UCC, UCU, AGU или AGC) (разница в первой, второй или третьей позиции). [49] Практическим следствием избыточности является то, что ошибки в третьей позиции триплетного кодона вызывают только молчаливую мутацию или ошибку, которая не повлияет на белок, поскольку гидрофильность или гидрофобность поддерживаются эквивалентной заменой аминокислот; например, кодон NUN (где N = любой нуклеотид) имеет тенденцию кодировать гидрофобные аминокислоты. NCN дает аминокислотные остатки, которые имеют небольшой размер и умеренную гидропатичность ; NAN кодирует гидрофильные остатки среднего размера. Генетический код настолько хорошо структурирован для гидропатичности, что математический анализ ( разложение по сингулярным значениям ) 12 переменных (4 нуклеотида x 3 позиции) дает замечательную корреляцию (C = 0,95) для прогнозирования гидропатичности кодируемой аминокислоты непосредственно из последовательности триплетных нуклеотидов, без трансляции. [50] [51] Обратите внимание, что в таблице ниже восемь аминокислот вообще не затронуты мутациями в третьей позиции кодона, тогда как на рисунке выше мутация во второй позиции, вероятно, вызовет радикальное изменение физико-химических свойств кодируемой аминокислоты. Тем не менее, изменения в первой позиции кодонов более важны, чем изменения во второй позиции в глобальном масштабе. [52] Причина может заключаться в том, что изменение заряда (с положительного на отрицательный заряд или наоборот) может произойти только при мутациях в первой позиции определенных кодонов, но не при изменениях во второй позиции любого кодона. Такое изменение заряда может иметь драматические последствия для структуры или функции белка. Этот аспект мог быть в значительной степени недооценен предыдущими исследованиями. [52]
Смещение использования кодона
Частота кодонов, также известная как смещение использования кодонов , может варьироваться от вида к виду с функциональными последствиями для контроля трансляции . Кодон варьируется в зависимости от организма; например, наиболее распространенным кодоном пролина в E. coli является CCG, тогда как у людей это наименее используемый кодон пролина. [53]
Альтернативные генетические коды
Нестандартные аминокислоты
В некоторых белках нестандартные аминокислоты заменяются на стандартные стоп-кодоны в зависимости от связанных сигнальных последовательностей в РНК-мессенджере. Например, UGA может кодировать селеноцистеин , а UAG может кодировать пирролизин . Селеноцистеин стал рассматриваться как 21-я аминокислота, а пирролизин — как 22-я. [55] И селеноцистеин, и пирролизин могут присутствовать в одном и том же организме. [55] Хотя генетический код обычно фиксирован в организме, ахейный прокариот Acetohalobium arabaticum может расширять свой генетический код с 20 до 21 аминокислоты (включая пирролизин) при различных условиях роста. [56]
Вариации
Логотип генетического кода митохондриального генома Globobulimina pseudospinescens от FACIL. Программа способна правильно сделать вывод об использовании простейшего митохондриального кода . [57] Логотип показывает 64 кодона слева направо, предсказанные альтернативы красным цветом (относительно стандартного генетического кода). Красная линия: стоп-кодоны. Высота каждой аминокислоты в стопке показывает, как часто она выравнивается с кодоном в гомологичных доменах белка. Высота стопки указывает на поддержку предсказания.
Первоначально существовал простой и широко принятый аргумент о том, что генетический код должен быть универсальным: а именно, что любое изменение в генетическом коде будет смертельным для организма (хотя Крик утверждал, что вирусы являются исключением). Это известно как аргумент «замороженной случайности» в пользу универсальности генетического кода. Однако в своей основополагающей статье о происхождении генетического кода в 1968 году Фрэнсис Крик все еще утверждал, что универсальность генетического кода во всех организмах была недоказанным предположением и, вероятно, не была верной в некоторых случаях. Он предсказал, что «код универсален (одинаков во всех организмах) или почти таков». [58] Первое изменение было обнаружено в 1979 году исследователями, изучающими митохондриальные гены человека . [59] После этого было обнаружено много незначительных изменений, [60] включая различные альтернативные митохондриальные коды. [61] Эти минорные вариации, например, включают трансляцию кодона UGA как триптофана у видов Mycoplasma и трансляцию CUG как серина, а не лейцина у дрожжей «клады CTG» (например, Candida albicans ). [62] [63] [64] Поскольку вирусы должны использовать тот же генетический код, что и их хозяева, модификации стандартного генетического кода могут мешать синтезу или функционированию вирусного белка. Однако вирусы, такие как тотивирусы, адаптировались к модификации генетического кода хозяина. [65] У бактерий и архей GUG и UUG являются распространенными стартовыми кодонами. В редких случаях некоторые белки могут использовать альтернативные стартовые кодоны. [60]
Удивительно, но различия в интерпретации генетического кода существуют также в человеческих ядерно-кодируемых генах: в 2016 году исследователи, изучающие трансляцию малатдегидрогеназы, обнаружили, что примерно в 4% мРНК, кодирующих этот фермент, стоп-кодон естественным образом используется для кодирования аминокислот триптофана и аргинина. [66] Этот тип перекодировки индуцируется контекстом стоп-кодона с высокой степенью считывания [67] и называется функциональным трансляционным считыванием . [68]
Несмотря на эти различия, все известные природные коды очень похожи. Механизм кодирования одинаков для всех организмов: трехосновные кодоны, тРНК , рибосомы, однонаправленное считывание и трансляция отдельных кодонов в отдельные аминокислоты. [69] Наиболее экстремальные вариации встречаются у некоторых инфузорий, где значение стоп-кодонов зависит от их положения в мРНК. Когда они находятся близко к 3'-концу, они действуют как терминаторы, тогда как во внутренних положениях они либо кодируют аминокислоты, как у Condylostoma magnum [70] , либо запускают сдвиг рамки рибосомы, как у Euplotes . [71]
Происхождение и вариации генетического кода, включая механизмы, лежащие в основе эволюционируемости генетического кода, широко изучались [72] [73], и некоторые исследования были проведены экспериментально по эволюции генетического кода некоторых организмов. [74] [75] [76] [77]
Вывод
Варианты генетических кодов, используемых организмом, могут быть выведены путем идентификации высококонсервативных генов, закодированных в этом геноме, и сравнения его использования кодонов с аминокислотами в гомологичных белках других организмов. Например, программа FACIL выводит генетический код путем поиска аминокислот в гомологичных доменах белков, которые чаще всего выровнены с каждым кодоном. Полученные вероятности аминокислот (или стоп-кодонов) для каждого кодона отображаются в логотипе генетического кода. [57]
По состоянию на январь 2022 года наиболее полное исследование генетических кодов было проведено Шульгиной и Эдди, которые провели скрининг 250 000 прокариотических геномов с помощью своего инструмента Codetta. Этот инструмент использует аналогичный подход к FACIL с более крупной базой данных Pfam . Несмотря на то, что NCBI уже предоставил 27 таблиц трансляции, авторам удалось найти 5 новых вариаций генетического кода (подтвержденных мутациями тРНК) и исправить несколько неверных атрибуций. [78] Позднее Codetta использовался для анализа изменения генетического кода у инфузорий . [79]
Источник
Генетический код является ключевой частью истории жизни , согласно одной из версий, согласно которой самовоспроизводящиеся молекулы РНК предшествовали жизни, какой мы ее знаем. Это гипотеза мира РНК . Согласно этой гипотезе, любая модель возникновения генетического кода тесно связана с моделью перехода от рибозимов (ферментов РНК) к белкам как основным ферментам в клетках. В соответствии с гипотезой мира РНК, молекулы транспортной РНК, по-видимому, эволюционировали до современных аминоацил-тРНК-синтетаз , поэтому последняя не может быть частью объяснения его закономерностей. [80]
Гипотетический случайно эволюционировавший генетический код далее мотивирует биохимическую или эволюционную модель его происхождения. Если бы аминокислоты были случайным образом назначены триплетным кодонам, было бы 1,5 × 10 84 возможных генетических кодов. [81] : 163 Это число находится путем подсчета количества способов, которыми 21 элемент (20 аминокислот плюс одна остановка) могут быть помещены в 64 ячейки, где каждый элемент используется по крайней мере один раз. [82] Однако распределение назначений кодонов в генетическом коде неслучайно. [83] В частности, генетический код кластеризует определенные назначения аминокислот.
Аминокислоты, которые разделяют один и тот же биосинтетический путь, как правило, имеют одинаковое первое основание в своих кодонах. Это может быть эволюционным реликтом раннего, более простого генетического кода с меньшим количеством аминокислот, который позже эволюционировал, чтобы кодировать больший набор аминокислот. [84] Это также может отражать стерические и химические свойства, которые имели другой эффект на кодон в ходе его эволюции. Аминокислоты со схожими физическими свойствами также, как правило, имеют схожие кодоны, [85] [86] уменьшая проблемы, вызванные точечными мутациями и неправильными переводами. [83]
Учитывая неслучайную схему генетического триплетного кодирования, обоснованная гипотеза о происхождении генетического кода могла бы рассмотреть несколько аспектов таблицы кодонов, таких как отсутствие кодонов для D-аминокислот, вторичные шаблоны кодонов для некоторых аминокислот, ограничение синонимичных позиций третьей позицией, небольшой набор всего из 20 аминокислот (вместо числа, приближающегося к 64) и связь шаблонов стоп-кодонов с шаблонами кодирования аминокислот. [87]
Три основные гипотезы рассматривают происхождение генетического кода. Многие модели принадлежат к одной из них или к гибриду: [88]
Случайная заморозка: генетический код был создан случайным образом. Например, ранние тРНК -подобные рибозимы могли иметь разное сродство к аминокислотам, при этом кодоны, появляющиеся из другой части рибозима, демонстрировали случайную изменчивость. Как только было закодировано достаточное количество пептидов , любое крупное случайное изменение в генетическом коде стало бы летальным; поэтому он стал «замороженным». [89]
Стереохимическое сродство: генетический код является результатом высокого сродства между каждой аминокислотой и ее кодоном или антикодоном; последний вариант подразумевает, что молекулы пре-тРНК сопоставляли свои соответствующие аминокислоты с помощью этого сродства. Позже в ходе эволюции это соответствие постепенно заменялось соответствием с помощью аминоацил-тРНК-синтетаз. [87] [90] [91]
Оптимальность: генетический код продолжал развиваться после своего первоначального создания, так что текущий код максимизирует некоторую функцию приспособленности , обычно это своего рода минимизация ошибок. [87] [88] [92]
Гипотезы рассматривали различные сценарии: [93]
Химические принципы управляют специфическим взаимодействием РНК с аминокислотами. Эксперименты с аптамерами показали, что некоторые аминокислоты имеют селективное химическое сродство к своим кодонам. [94] Эксперименты показали, что из 8 протестированных аминокислот, 6 показывают некоторую ассоциацию триплета РНК-аминокислоты. [81] [91]
Биосинтетическое расширение. Генетический код вырос из более простого более раннего кода через процесс «биосинтетического расширения». Первобытная жизнь «открыла» новые аминокислоты (например, как побочные продукты метаболизма ) и позже включила некоторые из них в механизм генетического кодирования. [95] Хотя было найдено много косвенных доказательств, указывающих на то, что в прошлом использовалось меньше типов аминокислот, [96] точные и подробные гипотезы о том, какие аминокислоты вошли в код и в каком порядке, являются спорными. [97] [98] Однако несколько исследований предположили, что Gly, Ala, Asp, Val, Ser, Pro, Glu, Leu, Thr могут принадлежать к группе аминокислот раннего добавления, тогда как Cys, Met, Tyr, Trp, His, Phe могут принадлежать к группе аминокислот позднего добавления. [99] [100] [101] [102]
Естественный отбор привел к назначению кодонов генетического кода, которые минимизируют эффекты мутаций . [103] Недавняя гипотеза [104] предполагает, что триплетный код произошел от кодов, которые использовали более длинные, чем триплетные кодоны (например, квадруплетные кодоны). Более длинное, чем триплетное, декодирование увеличило бы избыточность кодонов и было бы более устойчивым к ошибкам. Эта особенность могла бы позволить точное декодирование в отсутствие сложных трансляционных механизмов, таких как рибосома , например, до того, как клетки начали производить рибосомы.
Информационные каналы: Информационно-теоретические подходы моделируют процесс перевода генетического кода в соответствующие аминокислоты как подверженный ошибкам информационный канал. [105] Внутренний шум (то есть ошибка) в канале ставит перед организмом фундаментальный вопрос: как можно сконструировать генетический код, чтобы противостоять шуму [106] при точном и эффективном переводе информации? Эти модели «искажения скорости» [107] предполагают, что генетический код возник в результате взаимодействия трех конфликтующих эволюционных сил: потребности в разнообразных аминокислотах, [108] в толерантности к ошибкам [103] и в минимальных затратах ресурсов. Код возникает при переходе, когда сопоставление кодонов с аминокислотами становится неслучайным. Возникновение кода регулируется топологией, определяемой вероятными ошибками, и связано с проблемой раскраски карты . [109]
Теория игр: Модели, основанные на сигнальных играх, объединяют элементы теории игр, естественного отбора и информационных каналов. Такие модели использовались для предположения, что первые полипептиды, вероятно, были короткими и имели неферментативную функцию. Теоретико-игровые модели предполагали, что организация цепочек РНК в клетки могла быть необходима для предотвращения «обманного» использования генетического кода, т. е. предотвращения заполнения мира РНК древним эквивалентом вирусов. [110]
Стоп-кодоны: Кодоны для трансляционных остановок также являются интересным аспектом проблемы происхождения генетического кода. В качестве примера для рассмотрения эволюции стоп-кодонов было высказано предположение, что стоп-кодоны таковы, что они с наибольшей вероятностью рано завершат трансляцию в случае ошибки сдвига рамки . [111] Напротив, некоторые стереохимические молекулярные модели объясняют происхождение стоп-кодонов как «неопределяемое». [87]
^ Туранов АА, Лобанов АВ, Фоменко ДЕ, Моррисон ХГ, Согин МЛ, Клобутчер ЛА, Хэтфилд ДЛ, Гладышев ВН (январь 2009). "Генетический код поддерживает целевую вставку двух аминокислот одним кодоном". Science . 323 (5911): 259–61. doi :10.1126/science.1164748. PMC 3088105 . PMID 19131629.
^ Уотсон, Дж. Д.; Крик, Ф. Х. (30 мая 1953 г.). «Генетические последствия структуры дезоксирибонуклеиновой кислоты». Nature . 171 (4361): 964–967. Bibcode :1953Natur.171..964W. doi :10.1038/171964b0. ISSN 0028-0836. PMID 13063483. S2CID 4256010.
^ Штегманн, Ульрих Э. (1 сентября 2016 г.). «Переосмысление 'Генетического кодирования': анализ фактического использования». Британский журнал философии науки . 67 (3): 707–730. doi :10.1093/bjps/axv007. ISSN 0007-0882. PMC 4990703. PMID 27924115 .
^ Крик, Фрэнсис (10 июля 1990 г.). «Глава 8: Генетический код». What Mad Pursuit: A Personal View of Scientific Discovery . Basic Books. стр. 89–101. ISBN9780465091386. OCLC 1020240407.[ постоянная мертвая ссылка ]
^ Штраус, Бернард С. (1 марта 2019 г.). «Мартинас Ичас: «Архивист» Клуба Галстуков РНК». Генетика . 211 (3): 789–795. doi : 10.1534/genetics.118.301754. ISSN 1943-2631. PMC 6404253. PMID 30846543 .
^ "Фрэнсис Крик - Профили в результатах поиска по науке". profiles.nlm.nih.gov . Получено 21 июля 2022 г. .
^ ab Fry, Michael (2022). «Гипотеза адаптера Крика и открытие транспортной РНК: эксперимент, превосходящий теоретические предсказания». Философия, теория и практика в биологии . 14. doi : 10.3998/ptpbio.2628 . ISSN 2475-3025. S2CID 249112573.
^ Крик, Фрэнсис (1955). «О вырожденных шаблонах и гипотезе адаптера: заметка для Клуба РНК-связей». Национальная медицинская библиотека . Получено 21 июля 2022 г.
^ Уотсон, Джеймс Д. (2007). Избегайте скучных людей: уроки жизни в науке. Oxford University Press. стр. 112. ISBN978-0-19-280273-6. OCLC 47716375.
^ Барчишевская, Мирослава З.; Перриг, Патрик М.; Барчишевский, Ян (2016). «тРНК - золотой стандарт в молекулярной биологии». Молекулярные биосистемы . 12 (1): 12–17. дои : 10.1039/c5mb00557d. ПМИД 26549858.
^ Янофски, Чарльз (9 марта 2007 г.). «Установление триплетной природы генетического кода». Cell . 128 (5): 815–818. doi : 10.1016/j.cell.2007.02.029 . PMID 17350564. S2CID 14249277.
^ Nirenberg MW, Matthaei JH (октябрь 1961 г.). «Зависимость бесклеточного синтеза белка в E. coli от природных или синтетических полирибонуклеотидов». Труды Национальной академии наук Соединенных Штатов Америки . 47 (10): 1588–602. Bibcode : 1961PNAS...47.1588N. doi : 10.1073 /pnas.47.10.1588 . PMC 223178. PMID 14479932.
^ Gardner RS, Wahba AJ, Basilio C, Miller RS, Lengyel P, Speyer JF (декабрь 1962 г.). «Синтетические полинуклеотиды и аминокислотный код. VII». Труды Национальной академии наук Соединенных Штатов Америки . 48 (12): 2087–94. Bibcode : 1962PNAS...48.2087G. doi : 10.1073 /pnas.48.12.2087 . PMC 221128. PMID 13946552.
^ Wahba AJ, Gardner RS, Basilio C, Miller RS, Speyer JF, Lengyel P (январь 1963 г.). «Синтетические полинуклеотиды и аминокислотный код. VIII». Труды Национальной академии наук Соединенных Штатов Америки . 49 (1): 116–22. Bibcode : 1963PNAS...49..116W. doi : 10.1073/pnas.49.1.116 . PMC 300638. PMID 13998282 .
^ "Нобелевская премия по физиологии и медицине 1959 года" (пресс-релиз). Королевская шведская академия наук. 1959. Получено 27 февраля 2010. Нобелевская премия по физиологии и медицине 1959 года была присуждена совместно Северо Очоа и Артуру Корнбергу "за открытие механизмов биологического синтеза рибонуклеиновой кислоты и дезоксирибонуклеиновой кислоты".
^ Nirenberg M, Leder P, Bernfield M, Brimacombe R, Trupin J, Rottman F, O'Neal C (май 1965). «Кодовые слова РНК и синтез белка, VII. Об общей природе кода РНК». Труды Национальной академии наук Соединенных Штатов Америки . 53 (5): 1161–8. Bibcode :1965PNAS...53.1161N. doi : 10.1073/pnas.53.5.1161 . PMC 301388 . PMID 5330357.
^ "Нобелевская премия по физиологии и медицине 1968 года" (пресс-релиз). Королевская шведская академия наук. 1968 год . Получено 27 февраля 2010 года . Нобелевская премия по физиологии и медицине 1968 года была присуждена совместно Роберту В. Холли, Хару Гобинду Коране и Маршаллу В. Ниренбергу "за интерпретацию генетического кода и его функции в синтезе белка".
^ Эдгар Б. (октябрь 2004 г.). «Геном бактериофага Т4: археологические раскопки». Генетика . 168 (2): 575–82. doi :10.1093/genetics/168.2.575. PMC 1448817. PMID 15514035 .
^ Будиса, Недилько (23 декабря 2005 г.). Книга в библиотеке Wiley Online Library . doi :10.1002/3527607188. ISBN9783527312436.
^ Кубышкин, В.; Будиса, Н. (2017). «Синтетическое отчуждение микробных организмов с использованием генной инженерии кода: почему и как?». Biotechnology Journal . 12 (8): 1600097. doi :10.1002/biot.201600097. PMID 28671771.
^ Xie J, Schultz PG (декабрь 2005 г.). «Добавление аминокислот в генетический репертуар». Current Opinion in Chemical Biology . 9 (6): 548–54. doi :10.1016/j.cbpa.2005.10.011. PMID 16260173.
^ Wang Q, Parrish AR, Wang L (март 2009). «Расширение генетического кода для биологических исследований». Химия и биология . 16 (3): 323–36. doi :10.1016/j.chembiol.2009.03.001. PMC 2696486. PMID 19318213 .
^ Саймон М. (7 января 2005 г.). Emergent Computation: Emphasizing Bioinformatics. Springer Science & Business Media. стр. 105–106. ISBN978-0-387-22046-8.
^ "Создан первый стабильный полусинтетический организм | KurzweilAI". www.kurzweilai.net . 3 февраля 2017 г. . Получено 9 февраля 2017 г. .
^ Zhang Y, Lamb BM, Feldman AW, Zhou AX, Lavergne T, Li L, Romesberg FE (февраль 2017 г.). «Полусинтетический организм, разработанный для стабильного расширения генетического алфавита». Труды Национальной академии наук Соединенных Штатов Америки . 114 (6): 1317–1322. Bibcode : 2017PNAS..114.1317Z. doi : 10.1073/pnas.1616443114 . PMC 5307467. PMID 28115716 .
^ Han S, Yang A, Lee S, Lee HW, Park CB, Park HS (февраль 2017 г.). «Расширение генетического кода Mus musculus». Nature Communications . 8 : 14568. Bibcode : 2017NatCo...814568H. doi : 10.1038/ncomms14568. PMC 5321798. PMID 28220771 .
^ Циммер, Карл (15 мая 2019 г.). «Ученые создали бактерии с синтетическим геномом. Это искусственная жизнь? — Важным событием для синтетической биологии стало то, что колонии E. coli процветают с ДНК, созданной с нуля людьми, а не природой» . The New York Times . Архивировано из оригинала 2 января 2022 г. . Получено 16 мая 2019 г.
^ Фреденс, Юлиус и др. (15 мая 2019 г.). «Полный синтез Escherichia coli с перекодированным геномом». Nature . 569 (7757): 514–518. Bibcode :2019Natur.569..514F. doi :10.1038/s41586-019-1192-5. PMC 7039709 . PMID 31092918. S2CID 205571025.
^ Homo sapiens mitochondrion, complete genome. "Revised Cambridge Reference Sequence (rCRS): accession NC_012920", National Center for Biotechnology Information . Получено 27 декабря 2017 г.
^ ab King RC, Mulligan P, Stansfield W (10 января 2013 г.). Словарь генетики. OUP USA. стр. 608. ISBN978-0-19-976644-4.
^ Touriol C, Bornes S, Bonnal S, Audigier S, Prats H, Prats AC, Vagner S (2003). «Создание разнообразия изоформ белков путем альтернативной инициации трансляции в кодонах, отличных от AUG». Biology of the Cell . 95 (3–4): 169–78. doi : 10.1016/S0248-4900(03)00033-9 . PMID 12867081.
^ Maloy S (29 ноября 2003 г.). «Как бессмысленные мутации получили свои названия». Курс генетики микроорганизмов . Университет штата Сан-Диего . Получено 10 марта 2010 г.
^ Ссылки на изображение можно найти на странице Wikimedia Commons по адресу: Commons:File:Notable mutations.svg#References.
^ Гриффитс А. Дж., Миллер Дж. Х., Сузуки Д. Т., Левонтин Р. К. и др., ред. (2000). «Спонтанные мутации». Введение в генетический анализ (7-е изд.). Нью-Йорк: WH Freeman. ISBN978-0-7167-3520-5.
^ Freisinger E, Grollman AP, Miller H, Kisker C (апрель 2004 г.). «(Не)переносимость повреждений раскрывает понимание точности репликации ДНК». The EMBO Journal . 23 (7): 1494–505. doi :10.1038/sj.emboj.7600158. PMC 391067. PMID 15057282 .
^ Chang JC, Kan YW (июнь 1979). "бета 0 талассемия, бессмысленная мутация у человека". Труды Национальной академии наук Соединенных Штатов Америки . 76 (6): 2886–9. Bibcode :1979PNAS...76.2886C. doi : 10.1073/pnas.76.6.2886 . PMC 383714 . PMID 88735.
^ Бойле С., Ванде Вельде С., Кливленд Д.В. (октябрь 2006 г.). «АЛС: заболевание двигательных нейронов и их ненейрональных соседей». Нейрон . 52 (1): 39–59. дои : 10.1016/j.neuron.2006.09.018 . ПМИД 17015226.
^ Isbrandt D, Hopwood JJ, von Figura K, Peters C (1996). «Две новые мутации со сдвигом рамки считывания, вызывающие преждевременные стоп-кодоны у пациента с тяжелой формой синдрома Марото-Лами». Human Mutation . 7 (4): 361–3. doi : 10.1002/(SICI)1098-1004(1996)7:4<361::AID-HUMU12>3.0.CO;2-0 . PMID 8723688. S2CID 22693748.
^ Кроу Дж. Ф. (1993). «Насколько много мы знаем о спонтанных темпах мутаций у человека?». Экологический и молекулярный мутагенез . 21 (2): 122–9. Bibcode : 1993EnvMM..21..122C. doi : 10.1002/em.2850210205. PMID 8444142. S2CID 32918971.
^ Льюис Р. (2005). Генетика человека: концепции и применение (6-е изд.). Бостон, Массачусетс: McGraw Hill. стр. 227–228. ISBN978-0-07-111156-0.
^ Sawyer SA, Parsch J, Zhang Z, Hartl DL (апрель 2007 г.). «Распространенность положительного отбора среди почти нейтральных аминокислотных замен у дрозофилы». Труды Национальной академии наук Соединенных Штатов Америки . 104 (16): 6504–10. Bibcode : 2007PNAS..104.6504S. doi : 10.1073/pnas.0701572104 . PMC 1871816. PMID 17409186 .
^ Bridges KR (2002). "Малярия и эритроцит". Гарвард . Архивировано из оригинала 27 ноября 2011 г.
^ Drake JW, Holland JJ (ноябрь 1999 г.). «Скорость мутаций среди РНК-вирусов». Труды Национальной академии наук Соединенных Штатов Америки . 96 (24): 13910–3. Bibcode : 1999PNAS ...9613910D. doi : 10.1073/pnas.96.24.13910 . PMC 24164. PMID 10570172.
^ Холланд Дж., Шпиндлер К., Городиски Ф., Грабау Э., Никол С., ВандеПол С. (март 1982 г.). «Быстрая эволюция РНК-геномов». Science . 215 (4540): 1577–85. Bibcode :1982Sci...215.1577H. doi :10.1126/science.7041255. PMID 7041255.
^ de Visser JA, Rozen DE (апрель 2006 г.). «Клональная интерференция и периодический отбор новых полезных мутаций в Escherichia coli». Genetics . 172 (4): 2093–100. doi :10.1534/genetics.105.052373. PMC 1456385 . PMID 16489229.
^ Уотсон, Джеймс Д. (2008). Молекулярная биология гена. Пирсон/Бенджамин Каммингс. ISBN978-0-8053-9592-1. : 102–117 : 521–522
^ Фюллен Г., Юван Д.К. (1994). «Генетические алгоритмы и рекурсивный ансамблевый мутагенез в белковой инженерии». Complexity International 1.
^ ab Fricke, Markus (2019). «Глобальное значение вторичных структур РНК в последовательностях кодирования белков». Биоинформатика . 35 (4): 579–583. doi :10.1093/bioinformatics/bty678. PMC 7109657. PMID 30101307. S2CID 51968530 .
^ "Таблица частоты использования кодонов (диаграмма) - Genscript". www.genscript.com . Получено 4 февраля 2022 г. .
^ "Таблица использования кодонов". www.kazusa.or.jp .
^ ab Zhang Y, Baranov PV, Atkins JF, Gladyshev VN (май 2005). «Пирролизин и селеноцистеин используют разные стратегии декодирования». Журнал биологической химии . 280 (21): 20740–51. doi : 10.1074/jbc.M501458200 . PMID 15788401.
^ Prat L, Heinemann IU, Aerni HR, Rinehart J, O'Donoghue P, Söll D (декабрь 2012 г.). «Расширение генетического кода бактерий, зависящее от источника углерода». Труды Национальной академии наук Соединенных Штатов Америки . 109 (51): 21070–5. Bibcode : 2012PNAS..10921070P. doi : 10.1073/pnas.1218613110 . PMC 3529041. PMID 23185002 .
^ ab Dutilh BE, Jurgelenaite R, Szklarczyk R, van Hijum SA, Harhangi HR, Schmid M, de Wild B, Françoijs KJ, Stunnenberg HG, Strous M, Jetten MS, Op den Camp HJ, Huynen MA (июль 2011 г.). "FACIL: Быстрый и точный вывод генетического кода и логотип". Bioinformatics . 27 (14): 1929–33. doi :10.1093/bioinformatics/btr316. PMC 3129529 . PMID 21653513.
^ Фрэнсис Крик, 1968. «Происхождение генетического кода». J. Mol. Biol.
^ Barrell BG, Bankier AT, Drouin J (1979). «Другой генетический код в митохондриях человека». Nature . 282 (5735): 189–194. Bibcode :1979Natur.282..189B. doi :10.1038/282189a0. PMID 226894. S2CID 4335828.([1])
^ ab Elzanowski A, Ostell J (7 апреля 2008 г.). "Генетические коды". Национальный центр биотехнологической информации (NCBI) . Получено 10 марта 2010 г.
^ Jukes TH, Osawa S (декабрь 1990 г.). «Генетический код в митохондриях и хлоропластах». Experientia . 46 (11–12): 1117–26. doi :10.1007/BF01936921. PMID 2253709. S2CID 19264964.
^ Fitzpatrick DA, Logue ME, Stajich JE, Butler G (1 января 2006 г.). «Филогения грибов, основанная на 42 полных геномах, полученных из супердерева и комбинированного анализа генов». BMC Evolutionary Biology . 6 : 99. doi : 10.1186/1471-2148-6-99 . PMC 1679813. PMID 17121679 .
^ Santos MA, Tuite MF (май 1995). «Кодон CUG in vivo декодируется как серин, а не лейцин в Candida albicans». Nucleic Acids Research . 23 (9): 1481–6. doi :10.1093/nar/23.9.1481. PMC 306886. PMID 7784200 .
^ Батлер Г., Расмуссен М.Д., Лин М.Ф. и др. (июнь 2009 г.). «Эволюция патогенности и полового размножения в восьми геномах Candida». Nature . 459 (7247): 657–62. Bibcode :2009Natur.459..657B. doi :10.1038/nature08064. PMC 2834264 . PMID 19465905.
^ Тейлор DJ, Баллингер MJ, Боуман SM, Бруенн JA (2013). «Совместная эволюция вируса и хозяина под действием модифицированного ядерного генетического кода». PeerJ . 1 : e50. doi : 10.7717/peerj.50 . PMC 3628385 . PMID 23638388.
^ Hofhuis J, Schueren F, Nötzel C, Lingner T, Gärtner J, Jahn O, Thoms S (2016). «Функциональное сквозное расширение малатдегидрогеназы выявляет модификацию генетического кода». Open Biol . 6 (11): 160246. doi :10.1098/rsob.160246. PMC 5133446. PMID 27881739 .
^ Schueren F, Lingner T, George R, Hofhuis J, Gartner J, Thoms S (2014). «Пероксисомальная лактатдегидрогеназа генерируется путем трансляционного чтения у млекопитающих». eLife . 3 : e03640. doi : 10.7554/eLife.03640 . PMC 4359377 . PMID 25247702.
^ F. Schueren и S. Thoms (2016). «Функциональное трансляционное считывание: перспектива системной биологии». PLOS Genetics . 12 (8): e1006196. doi : 10.1371/journal.pgen.1006196 . PMC 4973966. PMID 27490485 .
^ Кубышкин В., Асеведо-Роча К.Г., Будиса Н. (февраль 2018 г.). «Об универсальных кодирующих событиях в биогенезе белков». Bio Systems . 164 : 16–25. Bibcode : 2018BiSys.164...16K. doi : 10.1016/j.biosystems.2017.10.004 . PMID 29030023.
^ Heaphy SM, Mariotti M, Gladyshev VN, Atkins JF, Baranov PV (ноябрь 2016 г.). «Новые варианты генетического кода инфузорий, включая переназначение всех трех стоп-кодонов на смысловые кодоны у Condylostoma magnum». Молекулярная биология и эволюция . 33 (11): 2885–2889. doi :10.1093/molbev/msw166. PMC 5062323. PMID 27501944 .
^ Лобанов А.В., Хифи С.М., Туранов А.А., Геращенко М.В., Пуччарелли С., Деварадж Р.Р. и др. (январь 2017 г.). «Позиционно-зависимое завершение и широко распространенный обязательный сдвиг рамки в переводе Euplotes». Структурная и молекулярная биология природы . 24 (1): 61–68. дои : 10.1038/nsmb.3330. ПМЦ 5295771 . ПМИД 27870834.
^ Кунин EV, Новожилов AS (февраль 2009). «Происхождение и эволюция генетического кода: всеобщая загадка». IUBMB Life . 61 (2): 91–111. doi :10.1002/iub.146. PMC 3293468. PMID 19117371 .
^ Sengupta S, Higgs PG (июнь 2015 г.). «Пути эволюции генетического кода в древних и современных организмах». Журнал молекулярной эволюции . 80 (5–6): 229–243. Bibcode :2015JMolE..80..229S. doi :10.1007/s00239-015-9686-8. PMID 26054480. S2CID 15542587.
^ Neumann H, Wang K, Davis L, Garcia-Alai M, Chin JW (март 2010 г.). «Кодирование нескольких неестественных аминокислот посредством эволюции рибосомы, декодирующей квадруплет». Nature . 18 (464): 441–444. doi :10.1038/nrm2005. PMID 16926858. S2CID 19385756.
^ Лю CC, Шульц PG (2010). «Добавление новых химических веществ в генетический код». Annual Review of Biochemistry . 79 : 413–444. doi :10.1146/annurev.biochem.052308.105824. PMID 20307192.
^ Chin JW (февраль 2014 г.). «Расширение и перепрограммирование генетического кода клеток и животных». Annual Review of Biochemistry . 83 : 379–408. doi : 10.1146/annurev-biochem-060713-035737. PMID 24555827.
^ Шульгина, Y; Эдди, SR (9 ноября 2021 г.). «Вычислительный экран для альтернативных генетических кодов в более чем 250 000 геномов». eLife . 10 . doi : 10.7554/eLife.71402 . PMC 8629427 . PMID 34751130.
^ Chen, W; Geng, Y; Zhang, B; Yan, Y; Zhao, F; Miao, M (4 апреля 2023 г.). «Остановиться или нет: профилирование переназначенных стоп-кодонов у инфузорий по всему геному». Молекулярная биология и эволюция . 40 (4). doi : 10.1093/molbev/msad064. PMC 1008964. PMID 36952281 .
^ Рибас де Пуплана Л., Тернер Р. Дж., Стир Б. А., Шиммель П. (сентябрь 1998 г.). «Происхождение генетического кода: тРНК старше своих синтетаз?». Труды Национальной академии наук Соединенных Штатов Америки . 95 (19): 11295–300. Bibcode : 1998PNAS...9511295D. doi : 10.1073 /pnas.95.19.11295 . PMC 21636. PMID 9736730.
^ ab Ярус, Майкл (2010). Жизнь из мира РНК: Внутренний предок. Издательство Гарвардского университета. ISBN978-0-674-05075-4.
^ "Функция Mathematica для # возможных расположений элементов в контейнерах? – Технические дискуссионные группы в Интернете — Сообщество Wolfram". community.wolfram.com . Получено 3 февраля 2017 г. .
^ ab Freeland SJ, Hurst LD (сентябрь 1998 г.). «Генетический код — один на миллион». Journal of Molecular Evolution . 47 (3): 238–48. Bibcode : 1998JMolE..47..238F. doi : 10.1007/PL00006381. PMID 9732450. S2CID 20130470.
^ Тейлор Ф.Дж., Коутс Д. (1989). «Код внутри кодонов». Bio Systems . 22 (3): 177–87. Bibcode : 1989BiSys..22..177T. doi : 10.1016/0303-2647(89)90059-2. PMID 2650752.
^ Di Giulio M (октябрь 1989). «Расширение, достигнутое минимизацией расстояний полярности в ходе эволюции генетического кода». Журнал молекулярной эволюции . 29 (4): 288–93. Bibcode : 1989JMolE..29..288D. doi : 10.1007/BF02103616. PMID 2514270. S2CID 20803686.
^ Wong JT (февраль 1980 г.). «Роль минимизации химических расстояний между аминокислотами в эволюции генетического кода». Труды Национальной академии наук Соединенных Штатов Америки . 77 (2): 1083–6. Bibcode : 1980PNAS...77.1083W. doi : 10.1073/pnas.77.2.1083 . PMC 348428. PMID 6928661 .
^ abcd Erives A (август 2011). "Модель прото-антикодоновых РНК-ферментов, требующих гомохиральности L-аминокислот". Journal of Molecular Evolution . 73 (1–2): 10–22. Bibcode :2011JMolE..73...10E. doi :10.1007/s00239-011-9453-4. PMC 3223571 . PMID 21779963.
^ ab Freeland SJ, Knight RD, Landweber LF, Hurst LD (апрель 2000 г.). «Ранняя фиксация оптимального генетического кода». Молекулярная биология и эволюция . 17 (4): 511–18. doi : 10.1093/oxfordjournals.molbev.a026331 . PMID 10742043.
^ Хопфилд Дж. Дж. (1978). «Происхождение генетического кода: проверяемая гипотеза, основанная на структуре тРНК, последовательности и кинетической корректуре». PNAS . 75 (9): 4334–4338. Bibcode :1978PNAS...75.4334H. doi : 10.1073/pnas.75.9.4334 . PMC 336109 . PMID 279919.
^ ab Yarus M, Widmann JJ, Knight R (ноябрь 2009 г.). «Связывание РНК-аминокислот: стереохимическая эра генетического кода». Journal of Molecular Evolution . 69 (5): 406–29. Bibcode : 2009JMolE..69..406Y. doi : 10.1007/s00239-009-9270-1 . PMID 19795157.
^ Браун, Шон М.; Ворачек, Вацлав; Фриланд, Стивен (5 апреля 2023 г.). «Как выглядел бы инопланетный аминокислотный алфавит и почему?». Астробиология . 23 (5): 536–549. Bibcode : 2023AsBio..23..536B. doi : 10.1089/ast.2022.0107. PMID 37022727. S2CID 257983174.
^ Knight RD, Freeland SJ, Landweber LF (июнь 1999). «Отбор, история и химия: три лица генетического кода». Trends in Biochemical Sciences . 24 (6): 241–7. doi :10.1016/S0968-0004(99)01392-4. PMID 10366854.
^ Knight RD, Landweber LF (сентябрь 1998 г.). «Рифма или причина: взаимодействия РНК-аргинина и генетический код». Химия и биология . 5 (9): R215–20. doi : 10.1016/S1074-5521(98)90001-1 . PMID 9751648.
^ Сенгупта С., Хиггс П. Г. (2015). «Пути эволюции генетического кода у древних и современных организмов». Журнал молекулярной эволюции . 80 (5–6): 229–243. Bibcode : 2015JMolE..80..229S. doi : 10.1007/s00239-015-9686-8. PMID 26054480. S2CID 15542587.
^ Брукс DJ, Фреско JR, Леск AM, Сингх M (октябрь 2002 г.). «Эволюция частот аминокислот в белках с течением времени: предполагаемый порядок введения аминокислот в генетический код». Молекулярная биология и эволюция . 19 (10): 1645–55. doi : 10.1093/oxfordjournals.molbev.a003988 . PMID 12270892.
^ Амирновин Р. (май 1997 г.). «Анализ метаболической теории происхождения генетического кода». Журнал молекулярной эволюции . 44 (5): 473–6. Bibcode : 1997JMolE..44..473A. doi : 10.1007/PL00006170. PMID 9115171. S2CID 23334860.
^ Ronneberg TA, Landweber LF, Freeland SJ (декабрь 2000 г.). «Проверка биосинтетической теории генетического кода: факт или артефакт?». Труды Национальной академии наук Соединенных Штатов Америки . 97 (25): 13690–5. Bibcode : 2000PNAS...9713690R. doi : 10.1073/pnas.250403097 . PMC 17637. PMID 11087835.
^ Трифонов, Эдвард Н. (сентябрь 2009 г.). «Происхождение генетического кода и самых ранних олигопептидов». Исследования в области микробиологии . 160 (7): 481–486. doi :10.1016/j.resmic.2009.05.004. PMID 19524038.
^ Хиггс, Пол Г.; Пудриц, Ральф Э. (июнь 2009 г.). «Термодинамическая основа синтеза пребиотических аминокислот и природа первого генетического кода». Астробиология . 9 (5): 483–490. arXiv : 0904.0402 . Bibcode : 2009AsBio...9..483H. doi : 10.1089/ast.2008.0280. ISSN 1531-1074. PMID 19566427. S2CID 9039622.
^ Chaliotis, Anargyros; Vlastaridis, Panayotis; Mossialos, Dimitris; Ibba, Michael; Becker, Hubert D.; Stathopoulos, Constantinos; Amoutzias, Grigorios D. (17 февраля 2017 г.). «Сложная эволюционная история аминоацил-тРНК-синтетаз». Nucleic Acids Research . 45 (3): 1059–1068. doi :10.1093/nar/gkw1182. ISSN 0305-1048. PMC 5388404. PMID 28180287 .
^ Ntountoumi, Chrysa; Vlastaridis, Panayotis; Mossialos, Dimitris; Stathopoulos, Constantinos; Iliopoulos, Ioannis; Promponas, Vasilios; Oliver, Stephen G; Amoutzias, Grigoris D (4 ноября 2019 г.). «Области низкой сложности в белках прокариот выполняют важные функциональные роли и высококонсервативны». Nucleic Acids Research . 47 (19): 9998–10009. doi :10.1093/nar/gkz730. ISSN 0305-1048. PMC 6821194. PMID 31504783 .
^ ab Freeland SJ, Wu T, Keulmann N (октябрь 2003 г.). «Дело в пользу минимизации ошибок стандартного генетического кода». Origins of Life and Evolution of the Biosphere . 33 (4–5): 457–77. Bibcode :2003OLEB...33..457F. doi :10.1023/A:1025771327614. PMID 14604186. S2CID 18823745.
^ Баранов П.В., Венин М., Прован Г. (2009). Gemmell NJ (ред.). "Уменьшение размера кодона как источник триплетного генетического кода". PLOS ONE . 4 (5): e5708. Bibcode :2009PLoSO...4.5708B. doi : 10.1371/journal.pone.0005708 . PMC 2682656 . PMID 19479032.
^ Tlusty T (ноябрь 2007 г.). «Модель возникновения генетического кода как перехода в зашумленном информационном канале». Журнал теоретической биологии . 249 (2): 331–42. arXiv : 1007.4122 . Bibcode : 2007JThBi.249..331T. doi : 10.1016/j.jtbi.2007.07.029. PMID 17826800. S2CID 12206140.
^ Sonneborn TM (1965). Bryson V, Vogel H (ред.). Эволюция генов и белков . Нью-Йорк: Academic Press. С. 377–397.
^ Tlusty T (февраль 2008 г.). "Сценарий искажения скорости для возникновения и эволюции шумных молекулярных кодов". Physical Review Letters . 100 (4): 048101. arXiv : 1007.4149 . Bibcode :2008PhRvL.100d8101T. doi :10.1103/PhysRevLett.100.048101. PMID 18352335. S2CID 12246664.
^ Sella G, Ardell DH (сентябрь 2006 г.). «Коэволюция генов и генетических кодов: замороженный несчастный случай Крика снова». Журнал молекулярной эволюции . 63 (3): 297–313. Bibcode : 2006JMolE..63..297S. doi : 10.1007/s00239-004-0176-7. PMID 16838217. S2CID 1260806.
^ Tlusty T (сентябрь 2010 г.). «Красочное происхождение генетического кода: теория информации, статистическая механика и возникновение молекулярных кодов». Physics of Life Reviews . 7 (3): 362–76. arXiv : 1007.3906 . Bibcode : 2010PhLRv...7..362T. doi : 10.1016/j.plrev.2010.06.002. PMID 20558115. S2CID 1845965.
^ Jee J, Sundstrom A, Massey SE, Mishra B (ноябрь 2013 г.). «Что информационно-асимметричные игры могут рассказать нам о контексте «замороженной аварии» Крика?». Журнал Королевского общества, Интерфейс . 10 (88): 20130614. doi :10.1098/rsif.2013.0614. PMC 3785830. PMID 23985735 .
^ Itzkovitz S, Alon U (2007). «Генетический код почти оптимален для включения дополнительной информации в последовательности кодирования белков». Genome Research . 17 (4): 405–412. doi :10.1101/gr.5987307. PMC 1832087 . PMID 17293451.
Дальнейшее чтение
Гриффитс А. Дж., Миллер Дж. Х., Сузуки Д. Т., Левонтин Р. К., Гилберт В. М. (1999). Введение в генетический анализ (7-е изд.). Сан-Франциско: WH Freeman. ISBN 978-0-7167-3771-1.
Альбертс Б., Джонсон А., Льюис Дж., Рафф М., Робертс К., Уолтер П. (2002). Молекулярная биология клетки (4-е изд.). Нью-Йорк: Garland Science. ISBN 978-0-8153-3218-3.
Лодиш Х.Ф., Берк А., Зипурски С.Л., Мацудайра П., Балтимор Д., Дарнелл Дж.Э. (2000). Молекулярно-клеточная биология (4-е изд.). Сан-Франциско: WH Freeman. ISBN 9780716737063.
Caskey CT, Leder P (апрель 2014 г.). «Код РНК: природный Розеттский камень». Труды Национальной академии наук Соединенных Штатов Америки . 111 (16): 5758–9. Bibcode : 2014PNAS..111.5758C. doi : 10.1073/pnas.1404819111 . PMC 4000803. PMID 24756939 .
Внешние ссылки
На Викискладе есть медиафайлы по теме «Генетический код» .
Генетические коды: Таблицы генетических кодов
База данных использования кодонов — таблицы частот кодонов для многих организмов





{kind=link}