Synapsida [a] — одна из двух основных групп позвоночных животных в группе Amniota , другая — Sauropsida (включающая рептилий и птиц ). Синапсиды были доминирующими наземными животными в позднем палеозое и раннем мезозое , но единственной группой, которая выжила в кайнозое, являются млекопитающие . [7] В отличие от других амниот, у синапсидов есть одно височное окно , отверстие в нижней части крыши черепа за каждой глазной орбитой , оставляющее костную дугу под каждой; это объясняет их название. [8] Отличительное височное окно развилось около 318 миллионов лет назад в позднекаменноугольный период, [1] когда синапсиды и завропсиды разошлись, но впоследствии было объединено с орбитой у ранних млекопитающих.
Животные (базальные амниоты), от которых произошли не млекопитающие синапсиды, традиционно назывались «рептилиями». Поэтому в классической систематике синапсиды описывались как млекопитающие рептилии , а нетерапсиды также назывались пеликозаврами или пеликозавровыми синапсидами. Эти парафилетические термины теперь вышли из употребления и используются только неформально (если вообще используются) в современной литературе. Теперь известно, что все существующие животные, традиционно называемые «рептилиями», более тесно связаны друг с другом, чем с синапсидами, поэтому слово «рептилия» было переопределено, чтобы обозначать только членов Sauropsida (птичья линия Amniota) или даже просто их подклад, и синапсиды не являются частью линии завропсидов в кладистическом смысле. [9] [10] [11] [12] Поэтому называть синапсидов «рептилиями, подобными млекопитающим» неправильно в соответствии с новым определением «рептилии», поэтому теперь их называют стволовыми млекопитающими , протомлекопитающими , парамлекопитающими или панмлекопитающими . [13] [14] [15]
Синапсиды были крупнейшими наземными позвоночными в пермском периоде, 299–251 миллион лет назад, с ними могли сравниться только некоторые крупные парейазавры в конце перми. Большинство линий синапсидов уровня пеликозавров были заменены в конце ранней перми более продвинутыми терапсидами . Численность и разнообразие синапсидов были сильно сокращены пермско-триасовым вымиранием . Известно, что только дицинодонты и эутериодонты терапсид (состоящие из тероцефалий и цинодонтий ) продолжили свое существование в триасовом периоде. Группа цинодонтов Probainognathia , в которую входят Mammaliaformes (млекопитающие и их более близкие предки), была единственными синапсидами, которые выжили после триаса. [16]
В триасовый период завропсиды -архозавры стали одними из самых крупных и многочисленных наземных позвоночных, с которыми по размеру могли соперничать только дицинодонты каннемейериформы , и дали начало динозаврам . Когда все нептичьи динозавры были уничтожены мел-палеогеновым вымиранием , млекопитающие синапсиды снова разнообразились и стали крупнейшими наземными и морскими животными на Земле.
На рубеже 20-го века считалось, что синапсиды являются одним из четырех основных подклассов рептилий . Однако это представление было опровергнуто при более внимательном изучении скелетных останков, поскольку синапсиды отличаются от рептилий своими отличительными височными отверстиями. Эти отверстия в костях черепа позволяли прикреплять более крупные челюстные мышцы, а значит, и более эффективный укус.
Синапсиды впоследствии считались более поздней рептильной линией, которая стала млекопитающими путем постепенной эволюции все более млекопитающих черт, отсюда и название «рептилии, подобные млекопитающим» (также известные как пеликозавры ). Они стали традиционными терминами для всех палеозойских (ранних) синапсид. Более поздние исследования также развенчали это представление, и теперь рептилии классифицируются как Sauropsida (завропсиды), сестринская группа для синапсидов, таким образом делая синапсидов их собственной таксономической группой. [9] [11]
В результате парафилетические термины «змееподобная рептилия» и «пеликозавр» считаются устаревшими и непопулярными в технической литературе, и вместо них используется термин «основное млекопитающее» (или иногда протомлекопитающее или парамлекопитающее). Филогенетически теперь понятно, что синапсиды составляют независимую ветвь древа жизни. [17] Монофилия синапсидов не вызывает сомнений , и такие выражения , как « Синапсиды содержат млекопитающих» и «синапсиды дали начало млекопитающим», оба выражают одну и ту же филогенетическую гипотезу. Эта терминология отражает современный кладистический подход к связям животных, согласно которому единственными действительными группами являются те, которые включают всех потомков общего предка: они известны как монофилетические группы, или клады .
Кроме того, Reptilia (рептилии) были пересмотрены в качестве монофилетической группы и считаются совершенно отличными от Synapsida, входящими в Sauropsida , сестринскую группу Synapsida в пределах Amniota. [18]
Синапсиды традиционно делятся для удобства на терапсид , продвинутую группу синапсидов и ветвь, в которой эволюционировали млекопитающие, и стволовых млекопитающих (ранее известных как пеликозавры ), включающих шесть других более примитивных семейств синапсидов. [19] Стволовые млекопитающие были все довольно похожи на ящериц, с раскинувшейся походкой и, возможно, роговыми щитками , в то время как терапсиды, как правило, имели более прямую позу и, возможно, волосы, по крайней мере, в некоторых формах. В традиционной таксономии синапсиды охватывают два различных класса : низкорослые стволовые млекопитающие дали начало более прямоходящим терапсидам, которые, в свою очередь, дали начало млекопитающим. В традиционной классификации позвоночных стволовые млекопитающие и терапсиды оба считались отрядами подкласса синапсидов. [8] [9]
В филогенетической номенклатуре термины используются несколько иначе, поскольку включены дочерние клады. Большинство статей, опубликованных в 21 веке, рассматривали «Pelycosaur» как неформальную группу примитивных членов. Therapsida по-прежнему используется как клад, включающий как традиционные семейства терапсид, так и млекопитающих.
Хотя Synapsida и Therapsida включают современных млекопитающих, на практике эти два термина используются почти исключительно по отношению к более базальным членам, которые лежат за пределами Mammaliaformes .

Синапсиды развили височное окно позади каждой глазницы на боковой поверхности черепа. Возможно, это обеспечило новые места прикрепления для челюстных мышц. Аналогичное развитие имело место у диапсидов , у которых развилось два, а не одно отверстие позади каждого глаза. Первоначально отверстия в черепе оставляли внутреннюю часть черепа покрытой только челюстными мышцами, но у высших терапсид и млекопитающих клиновидная кость расширилась, чтобы закрыть отверстие. Это оставило нижний край отверстия в виде дуги, простирающейся от нижних краев мозговой коробки.

Синапсиды характеризуются наличием дифференцированных зубов. К ним относятся клыки , моляры и резцы . [20] Тенденция к дифференциации обнаружена у некоторых лабиринтодонтов и ранних анапсидных рептилий в виде увеличения первых зубов на верхней челюсти , образуя своего рода протоклыки. Эта черта впоследствии была утрачена в линии диапсид , но получила дальнейшее развитие у синапсидов. Ранние синапсиды могли иметь два или даже три увеличенных «клыка», но у терапсид картина установилась до одного клыка в каждой половине верхней челюсти. Нижние клыки развились позже.
Переход челюсти является хорошим инструментом классификации , так как большинство других ископаемых признаков, которые создают хронологическую прогрессию от состояния, подобного рептилии, к млекопитающему, следуют прогрессии перехода челюсти. Нижняя челюсть , или нижняя челюсть, состоит из одной кости, несущей зубы, у млекопитающих (зубная кость), тогда как нижняя челюсть современных и доисторических рептилий состоит из конгломерата более мелких костей (включая зубную кость, суставную кость и другие). По мере того, как они эволюционировали в синапсиды, эти челюстные кости уменьшались в размере и либо терялись, либо, в случае суставной, постепенно перемещались в ухо, образуя одну из костей среднего уха: в то время как современные млекопитающие обладают молоточком , наковальней и стременем , базальные синапсиды (как и все другие четвероногие) обладают только стременем. Молоточек произошел от суставной кости (нижней челюстной кости), в то время как наковальня произошла от квадратной кости (черепной кости). [21]
Структуры челюстей млекопитающих также отличаются зубно-чешуйчатым челюстным суставом . В этой форме челюстного сустава зубная кость образует соединение с углублением в чешуйчатой кости , известным как суставная впадина . Напротив, все другие челюстные позвоночные, включая рептилий и не млекопитающих синапсид, обладают челюстным суставом, в котором одна из меньших костей нижней челюсти, сочленовная, соединяется с костью черепа, называемой квадратной костью , образуя сочленовно-квадратный челюстной сустав. У форм, переходных к млекопитающим, челюстной сустав состоит из большой нижней челюстной кости (похожей на зубную кость, обнаруженную у млекопитающих), которая не соединяется с чешуйчатой костью, но соединяется с квадратной костью с помощью отступающей суставной кости.
Со временем, по мере того как синапсиды становились все более млекопитающими и менее «рептильными», у них начало развиваться вторичное небо , разделяющее ротовую и носовую полости . У ранних синапсидов вторичное небо начало формироваться по бокам верхней челюсти , при этом рот и ноздри все еще оставались соединенными.
В конце концов, две стороны нёба начали изгибаться вместе, образуя форму U вместо формы C. Нёбо также начало расширяться назад к горлу, защищая весь рот и создавая полную нёбную кость . Верхняя челюсть также полностью закрыта. В окаменелостях одного из первых эутериодонтов ясно видны зачатки нёба. Более поздний тринаксодон имел полное и полностью закрытое нёбо, образуя четкую прогрессию. [22]

В дополнение к железистой коже, покрытой мехом, которая есть у большинства современных млекопитающих, современные и вымершие синапсиды обладают разнообразными модифицированными кожными покровами, включая остеодермы (костные доспехи, встроенные в кожу), щитки (защитные структуры дермы, часто с роговым покрытием), волосы или мех и чешуевидные структуры (часто образованные из модифицированных волос, как у панголинов и некоторых грызунов ). В то время как кожа рептилий довольно тонкая, у млекопитающих толстый дермальный слой. [23]
Предковый тип кожи синапсидов был предметом обсуждения. Типовой образец старейшего известного синапсида Asaphestera сохранил чешую . [24] Среди ранних синапсидов только два вида мелких варанопидов , как было обнаружено, обладают остеодермами ; [25] окаменелые ряды остеодерм указывают на костную броню на шее и спине. Однако некоторые недавние исследования поставили под сомнение размещение Varanopidae в Synapsida, [26] [27] в то время как другие возражают и склоняются к этому традиционному размещению. [28] [29] Отпечатки кожи указывают на то, что некоторые ранние синапсиды в базальной части обладали прямоугольными щитками на нижней стороне и хвосте. [30] [31] Щитки пеликозавров, вероятно, были неперекрывающимися дермальными структурами с роговым наложением, как у современных крокодилов и черепах . Они отличались по структуре от чешуек ящериц и змей , которые являются эпидермальной особенностью (как волосы млекопитающих или перья птиц). [32] Недавно полученные отпечатки кожи представителей рода Ascendonanus позволяют предположить, что по крайней мере варанопсиды развили чешую, похожую на чешую чешуи чешуйчатых . [33]
В настоящее время точно неизвестно, когда впервые появились такие характеристики млекопитающих, как волосы на теле и молочные железы , поскольку окаменелости лишь изредка дают прямые доказательства наличия мягких тканей. Исключительно хорошо сохранившийся череп Estemmenosuchus , терапсида из верхней перми, сохраняет гладкую кожу с тем, что, по-видимому, является железистыми углублениями, [34] животное, отмеченное как полуводное . [ 35] Древнейшая известная окаменелость, показывающая недвусмысленные отпечатки волос, - это келловейская (поздняя средняя юра ) Castorocauda и несколько современных харамийидов , оба не млекопитающие млекопитающие [36] [37] (см. ниже, однако). Предполагается, что более примитивные члены Cynodontia также имели мех или мехоподобное покрытие, основываясь на их предполагаемом теплокровном метаболизме. [38] Хотя более прямые доказательства наличия меха у ранних цинодонтов были предложены в виде небольших ямок на морде, возможно, связанных с усами , такие ямки также обнаружены у некоторых рептилий, у которых нет усов. [38] Есть доказательства того, что некоторые другие не млекопитающие цинодонты, более базальные, чем Castorocauda , такие как Morganucodon , имели хардеровы железы , которые связаны с уходом и поддержанием меха. Очевидное отсутствие этих желез у не млекопитающих может указывать на то, что мех не возник до этого момента в эволюции синапсидов. [38] Возможно, что мех и связанные с ним черты истинной теплокровности не появились, пока некоторые синапсиды не стали чрезвычайно маленькими и ночными, что потребовало более высокого метаболизма. [38] Считается, что самые древние примеры ночного образа жизни у синапсидов были у видов, которые жили более 300 миллионов лет назад. [39]
Однако позднепермские копролиты из России и, возможно, Южной Африки показывают, что по крайней мере некоторые синапсиды уже имели волосы до млекопитающих в эту эпоху. Это самые древние отпечатки волосоподобных структур на синапсидах. [40] [41]
Ранние синапсиды, еще до своего известного эволюционного дебюта в позднекаменноугольном периоде, [42] могли откладывать яйца с пергаментной оболочкой (кожистые), [43] в которых отсутствовал кальцинированный слой, как у большинства современных рептилий и однопроходных . Это также может объяснить, почему на сегодняшний день нет ископаемых свидетельств яиц синапсидов. [44] Поскольку они были уязвимы к высыханию, секреция из апокриновых желез могла помочь сохранить яйца влажными. [42]
По словам Офтедала, ранние синапсиды могли закапывать яйца во влажную почву, увлажняя их при контакте с влажной кожей, или могли переносить их во влажной сумке, подобной той, что у однопроходных ( ехидны переносят свои яйца и потомство во временной сумке [45] [46] ), хотя это ограничивало подвижность родителя. Последнее могло быть примитивной формой ухода за яйцами у синапсидов, а не просто закапыванием яиц, и ограничение подвижности родителя могло быть решено путем «парковки» яиц в гнездах во время добычи пищи или других видов деятельности и периодического увлажнения, что позволяло делать кладки большего размера, чем они могли бы поместиться в сумке (или сумках) за один раз, и за большими яйцами, которые было бы неудобно носить в сумке, было бы легче ухаживать. Основой предположения Офтедала является тот факт, что многие виды бесхвостых амфибий могут вынашивать икру или головастиков, прикрепленных к коже или заключенных в кожные «мешочки», а также то, как большинство саламандр сворачиваются вокруг своих икринок, чтобы сохранить их влажными, причем обе группы также имеют железистую кожу. [44]
Железы, участвующие в этом механизме, позже эволюционировали в настоящие молочные железы с множественными режимами секреции в сочетании с волосяными фолликулами. Сравнительный анализ эволюционного происхождения компонентов молока поддерживает сценарий, в котором секреции из этих желез эволюционировали в сложное, богатое питательными веществами молоко задолго до того, как возникли настоящие млекопитающие (некоторые из компонентов, возможно, предшествовали разделению между линиями синапсид и завропсид ). Цинодонты почти наверняка могли производить это, что позволило постепенно снизить массу желтка и, следовательно, размер яиц, что привело к все более птенцовым вылупляющимся детенышам, поскольку молоко стало основным источником питания, о чем свидетельствуют небольшой размер тела, наличие надлобковых костей и ограниченная замена зубов у продвинутых цинодонтов, а также у млекопитающих . [42] [43]
Воздушное движение впервые началось у не млекопитающих харамиидных цинодонтов, у Arboroharamiya , Xianshou , Maiopatagium и Vilevolodon, которые имели прекрасно сохранившиеся, покрытые мехом перепонки крыльев, которые тянулись через конечности и хвост. Их пальцы удлиненные, похожие на пальцы летучих мышей и кагуанов , и, вероятно, выполняют схожие функции как опоры крыльев, так и висения на ветвях деревьев. [47]
Среди настоящих млекопитающих воздушное движение впервые происходит у эутриконодонтов рода волатикотериев . У ископаемого волатикотерия имеется прекрасно сохранившийся мохнатый патагий с тонкими морщинами, который очень обширен, «зажимая» плохо сохранившиеся руки и ноги и простираясь до основания хвоста. [48] У близкого родственника аргентоконодона похожая бедренная кость, приспособленная к нагрузкам при полете, что указывает на схожий образ жизни. [49]
Териевые млекопитающие смогли освоить полет и планирование только спустя долгое время после того, как эти ранние воздухоплаватели вымерли, а самые ранние известные планирующие метатерии и летучие мыши появились в палеоцене . [50]
Недавно было обнаружено, что эндотермия была развита еще у офиакодона в позднем карбоне. Наличие фиброламеллярной кости, специализированного типа кости, которая может быстро расти, сохраняя при этом стабильную структуру, показывает, что офиакодон использовал свою высокую внутреннюю температуру тела для обеспечения быстрого роста, сопоставимого с современными эндотермами. [51]


В ходе эволюции синапсид, таксоны-предшественники в начале адаптивных радиаций, как правило, были производными плотоядными. Адаптивные радиации синапсид, как правило, происходили после событий вымирания, которые истощали биосферу и оставляли пустые ниши открытыми для заполнения новыми эволюционировавшими таксонами. У не млекопитающих синапсид, те таксоны, которые дали начало быстро диверсифицирующимся линиям, были как маленькими, так и большими по размеру тела, хотя после позднего триаса, прародители новых линий синапсид, как правило, были маленькими, неспециализированными генералистами. [52]
Самый ранний известный синапсид Asaphestera сосуществовал с самым ранним известным завропсидом Hylonomus , который жил в башкирском веке позднего карбона . [24] [15] Это был один из многих типов примитивных синапсидов, которые теперь неформально объединяются в качестве стволовых млекопитающих или иногда как протомлекопитающих (ранее известных как пеликозавры ). Ранние синапсиды распространились и разнообразились, став крупнейшими наземными животными в позднем каменноугольном и раннепермском периодах, достигая до 6 метров (20 футов) в длину. Они были раскидистыми, громоздкими, возможно, холоднокровными и имели небольшой мозг. У некоторых, таких как Dimetrodon , были большие паруса, которые могли помогать повышать температуру их тела . Несколько реликтовых групп просуществовали до поздней перми, но к середине поздней перми все они либо вымерли, либо эволюционировали в своих преемников — терапсид. [53]

Терапсиды, более продвинутая группа синапсидов, появились в средней перми и включали крупнейших наземных животных в средней и поздней перми . Они включали травоядных и плотоядных, от небольших животных размером с крысу (например, Robertia ), до крупных, громоздких травоядных весом в тонну и более (например, Moschops ). После процветания в течение многих миллионов лет, эти успешные животные были практически уничтожены пермско-триасовым массовым вымиранием около 250 миллионов лет назад, крупнейшим известным вымиранием в истории Земли , возможно, связанным с вулканическим событием Сибирских траппов .

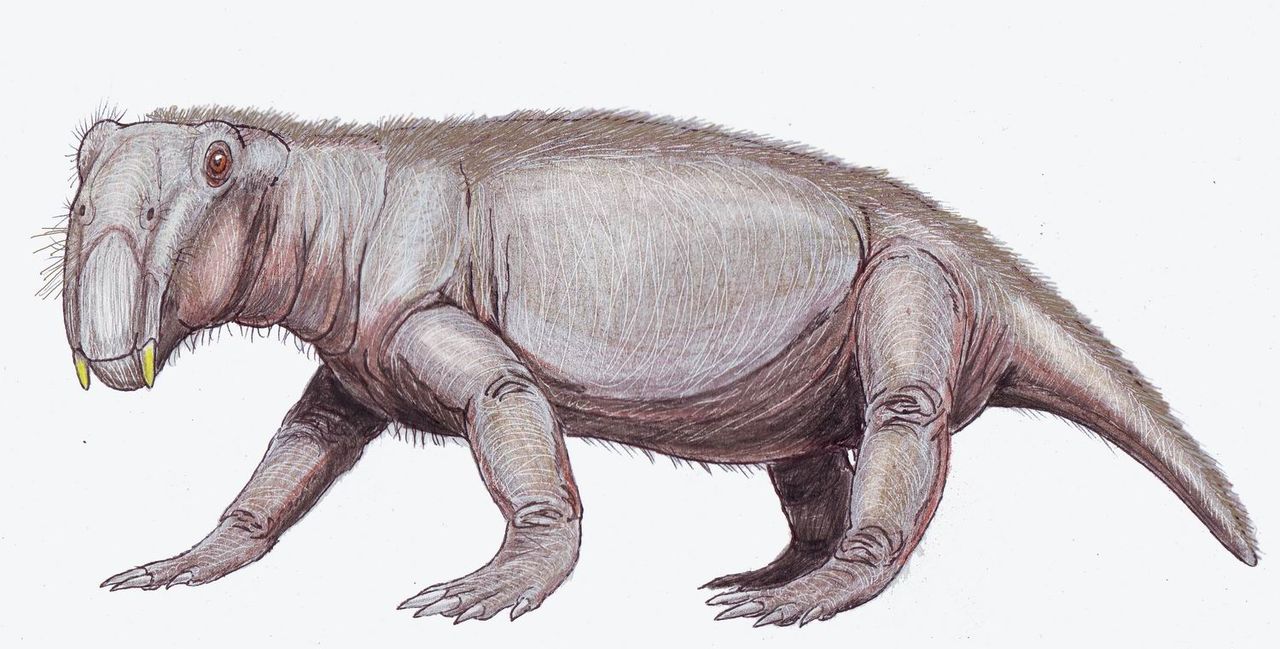
Лишь немногие терапсиды преуспели в новом раннетриасовом ландшафте ; среди них Lystrosaurus и Cynognathus , последний из которых появился позже в раннем триасе. Однако их сопровождали ранние архозавры (вскоре давшие начало динозаврам ) . Некоторые из этих архозавров, такие как Euparkeria , были маленькими и легко сложенными, в то время как другие, такие как Erythrosuchus , были такими же большими или больше самых крупных терапсид.
После пермского вымирания синапсиды насчитывали не более трех сохранившихся клад. Первая включала тероцефалов , которые просуществовали только первые 20 миллионов лет триасового периода. Вторая состояла из специализированных травоядных с клювом, известных как дицинодонты (например, Kannemeyeriidae ), которые включали некоторых членов, достигавших больших размеров (до тонны и более). И, наконец, были все более млекопитающие плотоядные, травоядные и насекомоядные цинодонты, включая эвцинодонтов оленекского яруса , ранним представителем которых был Cynognathus .

В отличие от дицинодонтов, которые были крупными, цинодонты становились все меньше и больше похожими на млекопитающих по мере развития триаса, хотя некоторые формы, такие как Trucidocynodon, оставались крупными. Первые млекопитающие произошли от цинодонтов в раннем норийском веке позднего триаса, около 225 млн лет назад.
В ходе эволюционной последовательности от раннего терапсида к цинодонту, к эуцинодонту и млекопитающему, основная нижняя челюстная кость, зубная кость, заменила соседние кости. Таким образом, нижняя челюсть постепенно стала просто одной большой костью, а несколько более мелких челюстных костей мигрировали во внутреннее ухо и обеспечили сложный слух.

Будь то из-за изменения климата, изменения растительности, экологической конкуренции или комбинации факторов, большинство оставшихся крупных цинодонтов (относящихся к Traversodontidae ) и дицинодонтов (семейства Kannemeyeriidae) исчезли к ретийскому веку, еще до триасово-юрского вымирания , которое убило большинство крупных нединозавровых архозавров . Оставшиеся мезозойские синапсиды были небольшими, от размера землеройки до похожего на барсука млекопитающего Repenomamus .

В юрском и меловом периодах оставшиеся не млекопитающие цинодонты были маленькими, например, тритилодон . Ни один цинодонт не вырастал больше кошки. Большинство юрских и меловых цинодонтов были травоядными , хотя некоторые были плотоядными . Семейство Tritheledontidae , которое впервые появилось около конца триаса, было плотоядным и сохранялось до средней юры . Другое, Tritylodontidae , впервые появилось в то же время, что и трителедонты, но было травоядным. Эта группа вымерла в конце раннего мелового периода. Обычно считается, что дицинодонты вымерли около конца триасового периода, но были доказательства того, что эта группа выжила, в виде шести фрагментов ископаемых костей, которые были найдены в меловых породах Квинсленда , Австралия. [54] Если это правда, это означало бы, что в Гондване существует значительная призрачная линия дицинодонтов . Однако в 2019 году эти окаменелости были повторно описаны как относящиеся к плейстоцену и, возможно, принадлежащие сумчатым дипротодонтидам . [55]
Сегодня 5500 видов живых синапсидов, известных как млекопитающие , включают как водные ( китообразные ), так и летающие ( летучие мыши ) виды, а также крупнейшее животное, когда-либо существовавшее ( синий кит ). Люди также являются синапсидами. Большинство млекопитающих живородящие и рождают живых детенышей, а не откладывают яйца, за исключением однопроходных .
Триасовые и юрские предки современных млекопитающих, а также их близкие родственники, имели высокую скорость метаболизма. Это означало потребление пищи (обычно считавшейся насекомыми) в гораздо большем количестве. Чтобы способствовать быстрому пищеварению , эти синапсиды развили жевание (пережевывание) и специализированные зубы, которые помогали жевать. Конечности также эволюционировали, чтобы двигаться под телом, а не в стороны, что позволило им дышать более эффективно во время передвижения. [56] Это помогло им удовлетворить их более высокие метаболические потребности.
Ниже представлена кладограмма наиболее общепринятой филогении синапсидов, показывающая длинную стволовую линию, включающую Mammalia и последовательно более базальные клады, такие как Theriodontia, Therapsida и Sphenacodontia: [57] [58]
Наибольшая неопределенность в филогении синапсидов лежит среди самых ранних членов группы, включая формы, традиционно помещаемые в Pelycosauria. В качестве одного из самых ранних филогенетических анализов Бринкман и Эберт (1983) поместили семейство Varanopidae с Caseasauria как наиболее базальным ответвлением линии синапсидов. Рейс (1986) удалил Varanopidae из Caseasauria, поместив его в более производное положение на стволе. В то время как большинство анализов находят Caseasauria наиболее базальной кладой синапсидов, анализ Бенсона (2012) поместил кладу, содержащую Ophiacodontidae и Varanopidae, как наиболее базальных синапсидов, при этом Caseasauria заняла более производное положение. Бенсон приписал эту пересмотренную филогению включению в свой анализ посткраниальных характеристик или особенностей скелета, отличных от черепа. Когда были включены только краниальные или черепные признаки, Caseasauria оставалась самой базальной синапсидной кладой. Ниже приведена кладограмма , измененная из анализа Бенсона (2012): [59]
Однако более позднее исследование филогении базальных синапсидов, включающее недавно описанные базальные казеиды и эотиридиды, [60] вернуло Caseasauria на ее позицию сестры по отношению ко всем остальным синапсидам. Броклхерст и др. (2016) [60] продемонстрировали, что многие из посткраниальных признаков, использованных Бенсоном (2012) для объединения Caseasauria с Sphenacodontidae и Edaphosauridae, отсутствовали в недавно обнаруженном посткраниальном материале эотиридид и, следовательно, были приобретены конвергентно.