Фибриллы (от лат. fibra [1] ) — структурные биологические материалы, которые встречаются практически во всех живых организмах . Не путать с волокнами или нитями , фибриллы, как правило, имеют диаметр от 10 до 100 нанометров (тогда как волокна представляют собой микро- и миллимасштабные структуры, а нити имеют диаметр приблизительно 10–50 нанометров). Фибриллы обычно не встречаются по отдельности, а являются частями более крупных иерархических структур, обычно встречающихся в биологических системах. Из-за распространенности фибрилл в биологических системах их изучение имеет большое значение в областях микробиологии , биомеханики и материаловедения .
Фибриллы состоят из линейных биополимеров и характеризуются стержнеобразными структурами с высоким отношением длины к диаметру. Они часто спонтанно выстраиваются в спиральные структуры. В задачах биомеханики фибриллы можно охарактеризовать как классические балки с приблизительно круглой площадью поперечного сечения в нанометровом масштабе. Таким образом, простые уравнения изгиба балок можно применять для расчета прочности фибрилл на изгиб в условиях сверхнизкой нагрузки. Как и у большинства биополимеров, отношения напряжения и деформации фибрилл имеют тенденцию показывать характерную область носка-пятки перед линейной эластичной областью . [2] В отличие от биополимеров, фибриллы не ведут себя как однородные материалы, поскольку было показано, что предел текучести изменяется в зависимости от объема, что указывает на структурные зависимости. [3] Было показано, что гидратация оказывает заметное влияние на механические свойства фибриллярных материалов. Было показано, что присутствие воды (альдегида) снижает жесткость коллагеновых фибрилл, а также увеличивает их скорость релаксации напряжения и прочность. [4] С биологической точки зрения содержание воды действует как механизм, повышающий жесткость фибриллярных структур, обеспечивая более высокое поглощение энергии и большую деформационную способность.
Механические прочностные свойства фибрилл возникают на молекулярном уровне. Силы, распределенные в волокне, представляют собой растягивающую нагрузку, переносимую фибриллой, и сдвигающие силы, ощущаемые из-за взаимодействия с другими молекулами фибрилл. Прочность на разрыв отдельных молекул коллагена в результате контролируется ковалентной химией между молекулами. Прочность на сдвиг между двумя молекулами коллагена контролируется слабыми дисперсионными и водородными связями взаимодействиями и некоторыми молекулярными ковалентными сшивками . Скольжение в системе происходит, когда эти межмолекулярные связи сталкиваются с приложенным напряжением, превышающим их прочность взаимодействия. [5] Разрыв межмолекулярных связей не приводит немедленно к разрушению, напротив, они играют существенную роль в рассеивании энергии, что снижает напряжение, ощущаемое материалом в целом, и позволяет ему выдерживать разрушение. Эти связи, часто водородные связи и дисперсионные взаимодействия Ван-дер-Ваальса , действуют как «жертвенные» связи, существующие с целью снижения напряжения в сети. Молекулярные ковалентные сшивки также играют ключевую роль в формировании сетей фибрилл. В то время как сшивание молекул может привести к прочным структурам, слишком большое количество сшивок в биополимерных сетях с большей вероятностью приведет к разрушению, поскольку сеть не способна рассеивать энергию, что приводит к материалу, который является прочным, но не жестким. Это наблюдается в обезвоженном или состаренном коллагене, что объясняет, почему с возрастом ткани человека становятся более хрупкими . [6]
Различия в структуре между фибриллами разного происхождения обычно определяются с помощью рентгеновской дифракции. [2] Сканирующий электронный микроскоп (СЭМ) можно использовать для наблюдения за специфическими деталями более крупных видов фибрилл, такими как характерные полосы 67 нм в коллагене, но часто он недостаточно точен для определения полной структуры.
Натуральные материалы демонстрируют сочетание обычно противоречивых механических свойств ( мягкость и прочность ) из-за их иерархических структур фибрилл в различных масштабах длины. [7] Эти фибриллы часто ориентированы в одном направлении, что приводит к анизотропной механической реакции в полученном биокомпозитном материале. [8] Это главное преимущество, поскольку большинство этих материалов выдерживают напряжения в одном направлении, и поэтому более высокий предел текучести и предел прочности при разрушении в направлении приложенного напряжения обеспечивает структурную целостность материала. Макро-, микро- и нанофибриллы позволяют материалу противостоять разрушению с помощью ряда механизмов сопротивления разрушению:
Эти механизмы работают вместе, чтобы противостоять разрушению, позволяя этим материалам выдерживать миллионы циклов нагрузки без сбоев, что необходимо для подвижных живых существ. Другим механическим преимуществом биополимеров является их способность к деформации, что является результатом существования прочных фибриллярных структур в более податливом матричном материале. Хорошая деформируемость интерфейсных матриц играет ключевую роль в обеспечении переориентации компонентов во время деформации. [8]
Фибриллогенез — это расширение тонких фибрилл, которое распространено в коллагеновых волокнах соединительной ткани . Точные механизмы фибриллогенеза до сих пор неизвестны, хотя многие гипотезы, вытекающие из фундаментальных исследований, помогают обнаружить множество возможных механизмов. В ранних экспериментах коллаген I можно было перегонять из тканей и рекомбинировать в фибриллы с контролем растворов. Более поздние исследования помогают понять состав и структуру участков связывания на мономерах коллагена. Коллаген синтезируется как растворимый предшественник, проколлаген, который поддерживает самосборку коллагена. Поскольку коллагеновые фибриллы имеют почти 50 связующих компонентов in vivo, определенное требование для генерации фибриллогенеза in vivo все еще остается скрытым. [9]
С помощью кислого или солевого раствора коллаген можно извлечь из тканей и перестроить в фибриллы, изменяя температуру или значение pH. [10] Эксперименты обнаружили силу притяжения между мономерами коллагена, которая способствует перестройке. [11] Коллаген служит предшественником, проколлагеном, в реакции синтеза, которая идентифицирует самополимеризацию коллагена.
В природе существует более 30 коллагенов, которые схожи по химическому составу, но различаются по кристаллической структуре. Безусловно, наиболее распространены коллагены I и II. Они инициативно образуют фибриллы in vitro, в то время как фибронектин, связывающие фибронектин, связывающие коллаген интегрины и коллаген V необходимы для формирования коллагена I, а коллаген XI — для формирования коллагена II. Таким образом, клеточные механизмы играют ключевую роль в процессе самосборки белка.
.png/1280px-Collagen_biosynthesis_(en).png)
Коллаген является основным структурным белком вне клеток во многих соединительных тканях животных. [12] Как основной компонент соединительной ткани, он имеет наибольшее количество среди белков у млекопитающих, занимая от 25% до 35% всего содержания белков в организме.
Фибриллы коллагена упакованы в извитую структуру. Кривая напряжения/деформации коллагена, например, сухожилия, может быть подразделена на несколько областей. Область небольших деформаций, область «носка», соответствует удалению макроскопической извитости, распрямлению, в коллагеновых фибриллах, видимых в световой микроскоп. При более крупных деформациях, области «пятки» и «линейной», никаких дальнейших структурных изменений не наблюдается.
Тропоколлаген — это молекулярный компонент волокна, состоящий из трех левозакрученных полипептидных цепей (красной, зеленой, синей), закрученных друг вокруг друга, образуя правозакрученную тройную спираль.
Мышцы сокращаются и растягиваются посредством управляемого скольжения/схватывания миозина, взаимодействующего с актиновыми волокнами. Актин состоит из двух полипептидов в спирали, а миозин имеет небольшую сердцевидную структуру, поперечный мостик. Процессы связывания и расцепления присоединения поперечных мостиков к актиновым филаментам помогают относительному движению этих коллагенов и, следовательно, всей мышцы.
Эластин — это волокнистый белок, распространенный в различных мягких тканях, таких как кожа, кровеносные сосуды и легочная ткань. Каждый мономер соединяется друг с другом, образуя трехмерную сеть, способную выдерживать более 200% деформации. [13]
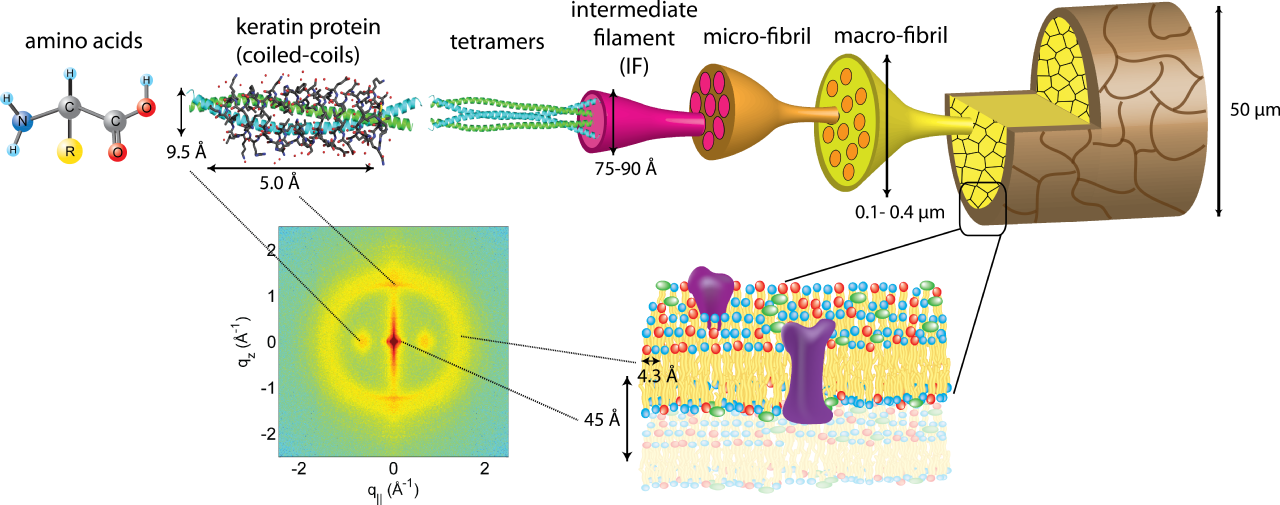
Кератин — это структурный белок, который в основном содержится в волосах, ногтях, копытах, рогах, иглах. [14] В основном кератин образован полипептидными цепями, которые скручены в α-спирали с серными поперечными связями или связаны в β-слои, соединенные водородными связями. β-кератин, который прочнее α-конформации, чаще встречается у птиц и рептилий.
Резилин — это эластомерный белок насекомых, состоящий из структуры α-спиралей и β-слоев. [15] Это один из самых упругих белков в природе. Он имеет низкую жесткость ~0,6 МПа, но высокий процент восстановления энергии ~98%, и эффективно помогает летающим насекомым хлопать крыльями или блохам прыгать.
Фибрилла паучьего шелка состоит из жесткой кристаллизованной структуры β-листов, отвечающей за прочность, и аморфной матрицы, окружающей ее, что улучшает прочность и способность к удлинению. [16] Она обладает исключительно высокой прочностью на разрыв и пластичностью при относительно низкой плотности по сравнению с другими натуральными фибриллами. Ее характеристики различаются у разных видов пауков для разных целей.
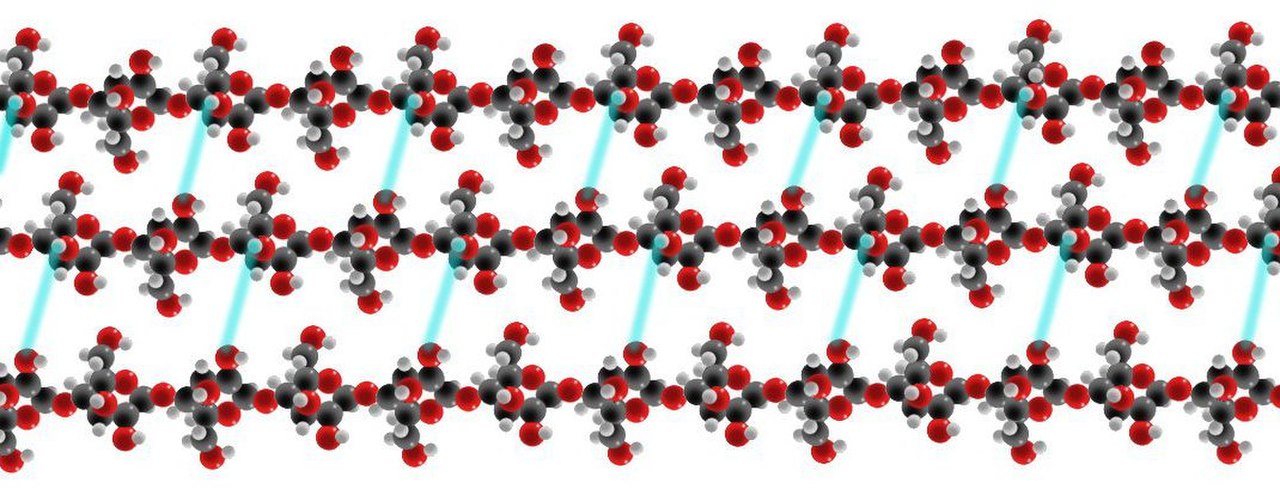
Первичная клеточная стенка получает свою заметную прочность на разрыв от молекул целлюлозы или длинных цепей остатков глюкозы, стабилизированных водородными связями . [17] Наблюдается, что цепи целлюлозы выстраиваются в перекрывающиеся параллельные массивы с аналогичной полярностью, образуя микрофибриллу целлюлозы. В растениях эти микрофибриллы целлюлозы выстраиваются в слои, формально известные как ламеллы, и стабилизируются в клеточной стенке поверхностными, длинными поперечно-сшитыми молекулами гликанов. Молекулы гликанов увеличивают сложность потенциальных сетей, в которые может конфигурироваться растительная целлюлоза. Соразмерный в первичной клеточной стенке как микрофибриллам целлюлозы, так и комплементарным сетям гликанов, пектин , который является полисахаридом, содержащим множество отрицательно заряженных единиц галактуроновой кислоты. [17] Кроме того, микрофибриллы целлюлозы также вносят вклад в форму растения посредством контролируемого расширения клеток. Стереоскопическое расположение микрофибрилл в клеточной стенке создает системы тургорного давления , что в конечном итоге приводит к клеточному росту и расширению. Микрофибриллы целлюлозы являются уникальными матричными макромолекулами, поскольку они собираются ферментами целлюлозосинтазы, расположенными на внеклеточной поверхности плазматической мембраны. [17] Считается, что растение может «предвидеть свою будущую морфологию, контролируя ориентацию микрофибрилл» с помощью механизма, при котором микрофибриллы целлюлозы располагаются поверх кортикального массива микротрубочек.
Говорят, что перемешивание данного образца амилозы приводит к образованию фибриллярных кристаллов, которые, как говорят, выпадают в осадок из маточного раствора. Эти длинные фибриллы можно визуализировать с помощью электронной микроскопии, выявляющей поперечные полосы, напоминающие шашлык . [ необходима цитата ] Фибриллы амилозы классифицируются по наличию одной из двух морфологий: одни с небольшими стержневидными фибриллами, а другие с кристаллами в форме реек.
Говорят, что фибриллярная структура древесины играет важную роль как в механической стабильности, так и в способности древесины иметь каналы для транспортировки минералов и воды. Сообщается, что ель (Picea abies), среди прочих, обладает целлюлозными фибриллами с нормализованным диаметром 2,5 нм. Также сообщается о связи между возрастом древесины и спиральным углом фибрилл по отношению к продольному направлению. Говорят, что ранняя древесина имеет постоянный угол покоя 4,6 ± 0,6°, тогда как поздняя древесина, как говорят, имеет переходную область от 4,6° до 19,8 ± 0,7°. [18] В поздней древесине две области спирального угла целлюлозных фибрилл не являются непрерывными, что означает, что в «старых» деревьях есть две независимые структуры трахеид, отвечающие различным механическим требованиям. Более того, продольно ориентированные фибриллы улучшают прочность на растяжение, тогда как добавление наклоненных на 20° фибрилл, исключительно для трахеид поздней древесины, обеспечивает устойчивость к сжатию. [18]
Чтобы имитировать прочную адгезию, легкое отсоединение и самоочищающиеся свойства подушечки пальца геккона, можно создать фибриллярный клей. Эти эксплуатационные характеристики вытекают из базовой иерархической структуры, которая состоит из миллиона микрофибрилл, называемых щетинками , которые, в свою очередь, состоят из миллиардов наноразмерных ветвей, называемых лопаточками .
Имитация этого явления включает четыре отдельных этапа проектирования: [19]
Для имитации зрелой костной матрицы можно использовать самоорганизующиеся фибриллы для выравнивания заданной минеральной матрицы. Это достигается с помощью самоорганизующейся молекулы с гидрофобным алкильным хвостом и гидрофильной олигопептидной головкой. Эти молекулы образуют мицеллярные структуры in situ и дисульфидные мостики при низком pH, что приводит к образованию и кристаллизации полимерных нанофибрилл массой 200 кДа. [ необходима цитата ] Минеральная матрица в конечном итоге взаимодействует с синтетической фибриллой через остаток фосфосерина, что приводит к зародышеобразованию и росту минерала.
{{cite journal}}: Цитировать журнал требует |journal=( помощь )