Эхолокация , также называемая биосонаром , является биологически активным сонаром , используемым несколькими группами животных как в воздухе, так и под водой. Эхолокирующие животные издают звуки и слушают эхо этих звуков, которые возвращаются от различных объектов рядом с ними. Они используют эти эхо для определения местоположения и идентификации объектов. Эхолокация используется для навигации , поиска пищи и охоты на добычу .
Эхолокационные сигналы могут быть частотно-модулированными (ЧМ, с переменной высотой тона во время сигнала) или постоянной частоты (ПЧ). ЧМ обеспечивает точную дискриминацию по дальности для локализации добычи за счет уменьшения рабочего диапазона. ПЧ позволяет определять как скорость добычи, так и ее движения с помощью эффекта Доплера . ПЧ может быть лучшим вариантом для тесных, загроможденных сред, в то время как ПЧ может быть лучшим вариантом для открытых сред или для охоты из положения сидя.
Эхолокационные животные включают млекопитающих, особенно зубатых китов и некоторые виды летучих мышей , а также, используя более простые формы, виды других групп, такие как землеройки . Несколько видов птиц из двух групп пещерных птиц используют эхолокацию, а именно пещерные саланганы и масляная птица .
Некоторые животные-жертвы, на которых охотятся эхолокационные летучие мыши, принимают активные контрмеры , чтобы избежать захвата. К ним относятся избегание хищников, отклонение атаки и использование ультразвуковых щелчков, которые развили множество функций, включая апосематизм, мимикрию химически защищенных видов и глушение эхолокации.
Термин «эхолокация» был придуман в 1944 году американским зоологом Дональдом Гриффином , который вместе с Робертом Галамбосом впервые продемонстрировал это явление у летучих мышей. [1] [2] Как описал Гриффин в своей книге, [3] итальянский ученый XVIII века Ладзаро Спалланцани с помощью серии сложных экспериментов пришел к выводу, что когда летучие мыши летают ночью, они полагаются на какое-то чувство, помимо зрения, но он не обнаружил, что другим чувством является слух. [4] [5] Швейцарский врач и натуралист Луи Жюрин повторил эксперименты Спалланцани (используя разные виды летучих мышей) и пришел к выводу, что когда летучие мыши охотятся ночью, они полагаются на слух. [6] [7] [8] В 1908 году Вальтер Луис Хан подтвердил выводы Спалланцани и Жюрина. [9]
В 1912 году изобретатель Хирам Максим независимо предположил, что летучие мыши используют звук ниже диапазона человеческого слуха , чтобы избегать препятствий. [10] В 1920 году английский физиолог Гамильтон Хартридж правильно предположил, что летучие мыши вместо этого используют частоты выше диапазона человеческого слуха . [11] [12]
Эхолокация у зубатых китов ( odontocete ) была должным образом описана лишь спустя два десятилетия после работы Гриффина и Галамбоса, Шевиллом и Макбрайдом в 1956 году. [13] Однако в 1953 году Жак Ив Кусто в своей первой книге «Безмолвный мир» предположил , что у морских свиней есть что-то вроде сонара , судя по их навигационным способностям. [14]
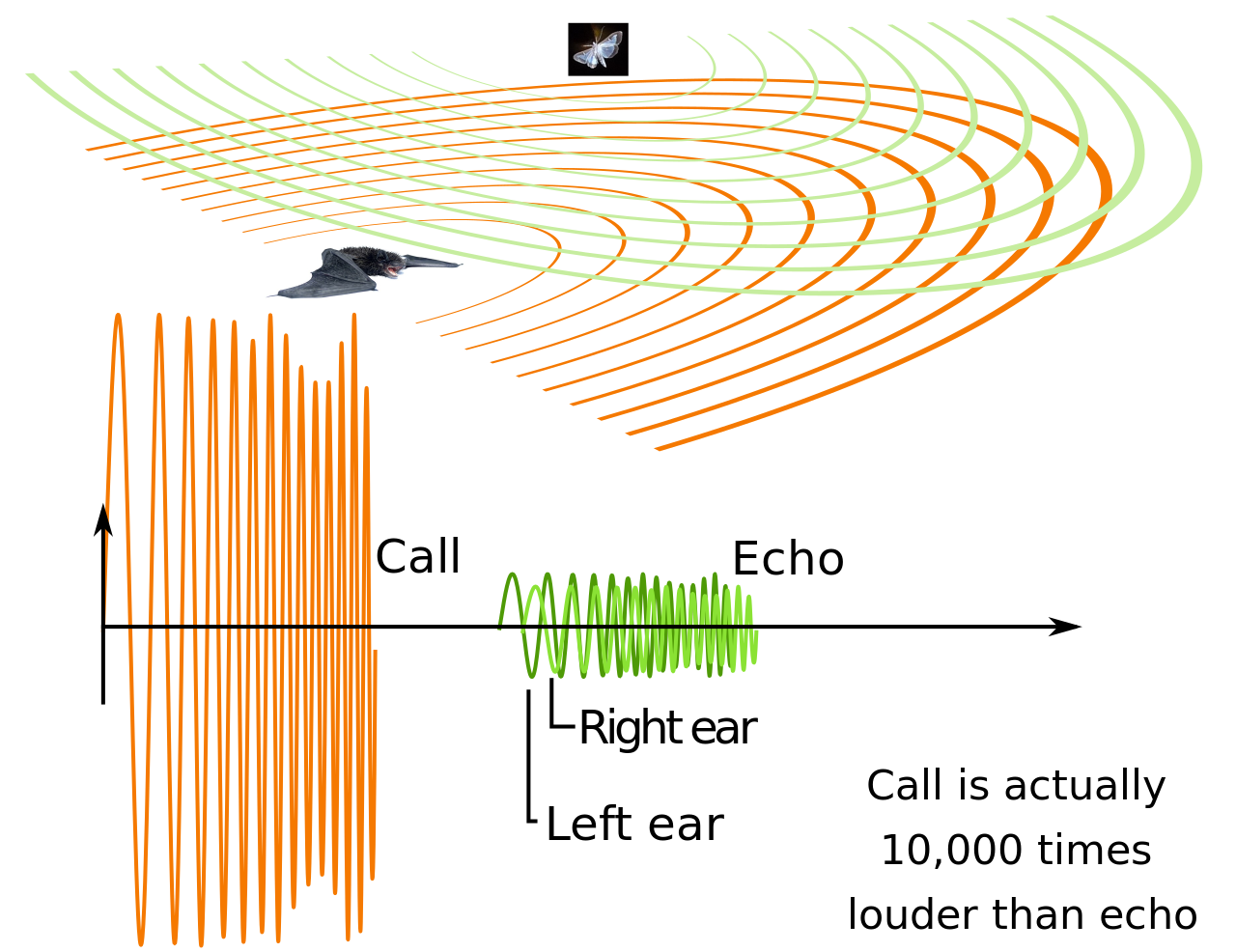
Эхолокация — это активный сонар , использующий звуки, издаваемые самим животным. Определение дальности достигается путем измерения временной задержки между собственным звуковым излучением животного и любыми эхами, которые возвращаются из окружающей среды. Относительная интенсивность звука, полученного каждым ухом, а также временная задержка между прибытием в два уха, предоставляют информацию о горизонтальном угле (азимуте), с которого приходят отраженные звуковые волны. [15]
В отличие от некоторых созданных человеком сонаров, которые полагаются на множество чрезвычайно узких лучей и множество приемников для локализации цели ( многолучевой сонар ), эхолокация животных имеет только один передатчик и два приемника (уши), расположенных немного отдельно. Эхо, возвращающееся в уши, приходит в разное время и с разной интенсивностью, в зависимости от положения объекта, генерирующего эхо. Различия во времени и громкости используются животными для восприятия расстояния и направления. С помощью эхолокации летучая мышь или другое животное может определить не только, куда оно идет, но и насколько велико другое животное, что это за животное и другие характеристики. [16] [17]
Описание разнообразия эхолокационных сигналов требует изучения частотных и временных характеристик сигналов. Именно вариации в этих аспектах создают эхолокационные сигналы, подходящие для различных акустических сред и охотничьего поведения. Сигналы летучих мышей были наиболее интенсивно исследованы, но принципы применимы ко всем эхолокационным сигналам. [18] [19]
Частоты криков летучих мышей варьируются от 11 кГц до 212 кГц. [20] Насекомоядные летучие мыши-ястребки, которые преследуют добычу на открытом воздухе, имеют частоту крика от 20 кГц до 60 кГц, поскольку именно эта частота обеспечивает наилучший диапазон и четкость изображения, а также делает их менее заметными для насекомых. [21] Однако низкие частоты являются адаптивными для некоторых видов с различной добычей и средой обитания. Euderma maculatum , вид летучих мышей, питающийся молью , использует особенно низкую частоту 12,7 кГц, которую моль не слышит. [22]
Эхолокационные сигналы могут состоять из двух различных типов частотной структуры: частотно-модулированные (FM) свипы и тоны постоянной частоты (CF). Конкретный сигнал может состоять из одной, другой или обеих структур. FM-свип представляет собой широкополосный сигнал, то есть он содержит нисходящую свип-волну через диапазон частот. CF-тон представляет собой узкополосный сигнал: звук остается постоянным на одной частоте на протяжении всей своей продолжительности. [23]
Эхолокационные сигналы летучих мышей измерялись при интенсивности от 60 до 140 децибел . [24] Некоторые виды летучих мышей могут изменять интенсивность своего сигнала во время сигнала, снижая ее по мере приближения к объектам, которые сильно отражают звук. Это не позволяет возвращающемуся эху оглушить летучую мышь. [19] Высокоинтенсивные сигналы, такие как сигналы летучих мышей-ястребов (133 дБ), являются адаптивными для охоты в открытом небе. Их высокоинтенсивные сигналы необходимы даже для умеренного обнаружения окружающей среды, поскольку воздух обладает высокой поглощающей способностью ультразвука, а размер насекомых обеспечивает лишь небольшую цель для отражения звука. [25] Кроме того, так называемые «шепчущие летучие мыши» адаптировали низкоамплитудную эхолокацию, так что их добыча, мотыльки, которые способны слышать эхолокационные сигналы, менее способны обнаружить и избежать приближающейся летучей мыши. [22] [26]
Одиночный эхолокационный сигнал (сигнал, представляющий собой один непрерывный след на звуковой спектрограмме , и серия сигналов, включающая последовательность или проход) может длиться от менее 3 до более 50 миллисекунд. Длительность импульса составляет около 3 миллисекунд у летучих мышей FM, таких как Phyllostomidae и некоторые Vespertilionidae; от 7 до 16 миллисекунд у летучих мышей с квазипостоянной частотой (QCF), таких как другие Vespertilionidae, Emballonuridae и Molossidae; и от 11 миллисекунд (Hipposideridae) до 52 миллисекунд (Rhinolophidae) у летучих мышей CF. [27] Длительность также зависит от стадии поведения по поимке добычи, в которой участвует летучая мышь, и обычно уменьшается, когда летучая мышь находится на заключительных стадиях поимки добычи — это позволяет летучей мыши кричать быстрее, не накладываясь друг на друга. Сокращение продолжительности достигается за счет уменьшения общего количества звука, который может отражаться от объектов и быть услышанным летучей мышью. [20]
Временной интервал между последовательными эхолокационными сигналами (или импульсами) определяет два аспекта восприятия летучей мыши. Во-первых, он устанавливает, как быстро обновляется слуховая информация летучей мыши. Например, летучие мыши увеличивают частоту повторения своих сигналов (то есть уменьшают интервал импульсов), когда они наводятся на цель. Это позволяет летучей мыши получать новую информацию о местоположении цели с большей скоростью, когда она больше всего в этом нуждается. Во-вторых, интервал импульсов определяет максимальный диапазон, на котором летучие мыши могут обнаруживать объекты. Это связано с тем, что летучие мыши могут отслеживать только эхо от одного сигнала за раз; как только они делают другой сигнал, они перестают слушать эхо от предыдущего сигнала. Например, интервал импульсов в 100 мс (типичный для летучей мыши, ищущей насекомых) позволяет звуку распространяться по воздуху примерно на 34 метра, поэтому летучая мышь может обнаруживать объекты только на расстоянии до 17 метров (звук должен распространяться туда и обратно). При интервале импульса 5 мс (типичном для летучей мыши в последние моменты попытки захвата) летучая мышь может обнаруживать объекты только на расстоянии до 85 см. Поэтому летучей мыши постоянно приходится делать выбор между быстрым получением новой информации и обнаружением объектов на большом расстоянии. [28]
Главное преимущество, предоставляемое FM-сигналом, — это чрезвычайно точное различение диапазона или локализация цели. JA Simmons продемонстрировал этот эффект с помощью серии экспериментов, которые показали, как летучие мыши, использующие FM-сигналы, могут различать две отдельные цели, даже если цели находятся на расстоянии менее половины миллиметра друг от друга. Эта способность обусловлена широкополосным распространением сигнала, что позволяет лучше разрешать временную задержку между вызовом и возвращающимся эхом, тем самым улучшая их взаимную корреляцию. Если к FM-сигналу добавляются гармонические частоты, то эта локализация становится еще более точной. [29] [30] [31]
Одним из возможных недостатков FM-сигнала является уменьшенный рабочий диапазон вызова. Поскольку энергия вызова распределена по многим частотам, расстояние, на котором FM-летучая мышь может обнаруживать цели, ограничено. [32] Это отчасти связано с тем, что любое эхо, возвращающееся на определенной частоте, может быть оценено только в течение короткой доли миллисекунды, поскольку быстрое нисходящее движение вызова не остается на какой-либо одной частоте долгое время. [30]
Структура сигнала CF адаптивна, поскольку она позволяет летучей мыши CF-bat обнаруживать как скорость цели, так и трепетание крыльев цели, как смещенные частоты Доплера. Смещение Доплера представляет собой изменение частоты звуковой волны и возникает в двух соответствующих ситуациях: когда летучая мышь и ее цель движутся относительно друг друга, и когда крылья цели колеблются вперед и назад. Летучие мыши CF-bat должны компенсировать сдвиги Доплера, понижая частоту своего сигнала в ответ на эхо повышенной частоты – это гарантирует, что возвращающееся эхо останется на частоте, на которую уши летучей мыши наиболее точно настроены. Колебание крыльев цели также вызывает сдвиги амплитуды, что дает летучей мыши CF-bat дополнительную помощь в различении летящей цели от неподвижной. [33] [29] Подковоносые летучие мыши охотятся таким образом. [34]
Кроме того, поскольку энергия сигнала вызова CF сосредоточена в узкой полосе частот, рабочий диапазон вызова намного больше, чем у сигнала FM. Это основано на том факте, что эхо, возвращающееся в пределах узкой полосы частот, может быть суммировано по всей длине вызова, что поддерживает постоянную частоту до 100 миллисекунд. [30] [32]
Компонент FM отлично подходит для охоты на добычу во время полета в тесной, загроможденной среде. Два аспекта сигнала FM объясняют этот факт: точная локализация цели, обеспечиваемая широкополосным сигналом, и короткая продолжительность сигнала. Первый из них важен, поскольку в загроможденной среде летучие мыши должны уметь различать свою добычу среди большого количества фонового шума. Возможности трехмерной локализации широкополосного сигнала позволяют летучей мыши делать именно это, предоставляя ей то, что Симмонс и Стайн (1980) называют «стратегией отсечения помех». [31] Эта стратегия дополнительно улучшается за счет использования гармоник, которые, как было сказано ранее, улучшают локализационные свойства сигнала. Короткая продолжительность сигнала FM также лучше всего подходит для тесной, загроможденной среды, поскольку она позволяет летучей мыши чрезвычайно быстро издавать множество сигналов без перекрытия. Это означает, что летучая мышь может получать почти непрерывный поток информации — важный, когда объекты находятся близко, потому что они быстро пролетят мимо — не путая, какое эхо соответствует какому сигналу. [33] [29]
Компонент CF часто используется летучими мышами, охотящимися за добычей во время полета в открытой, свободной от помех среде, или летучими мышами, которые ждут на насестах, пока их добыча не появится. Успех первой стратегии обусловлен двумя аспектами сигнала CF, оба из которых обеспечивают превосходные способности обнаружения добычи. Во-первых, больший рабочий диапазон сигнала позволяет летучим мышам обнаруживать цели, находящиеся на больших расстояниях — обычная ситуация в открытых средах. Во-вторых, длина сигнала также подходит для целей на больших расстояниях: в этом случае снижается вероятность того, что длинный сигнал будет перекрываться возвращающимся эхом. Последняя стратегия становится возможной благодаря тому факту, что длинный, узкополосный сигнал позволяет летучей мыши обнаруживать доплеровские сдвиги, которые могут быть вызваны насекомым, движущимся либо к сидящей летучей мыши, либо от нее. [33] [31] [29]
Эхолокация встречается у различных млекопитающих и птиц, как описано ниже. [35] Она развивалась неоднократно, что является примером конвергентной эволюции . [29] [36]

Эхолокационные летучие мыши используют эхолокацию для навигации и добычи пищи, часто в полной темноте. Обычно они выходят из своих убежищ в пещерах, на чердаках или на деревьях в сумерках и охотятся на насекомых ночью. Используя эхолокацию, летучие мыши могут определить, насколько далеко находится объект, его размер, форму и плотность, а также направление (если таковое имеется), в котором движется объект. Использование ими эхолокации, наряду с мощным полетом, позволяет им занимать нишу, где часто много насекомых (которые выходят ночью, так как тогда меньше хищников), меньше конкуренции за пищу и меньше видов, которые могут охотиться на самих летучих мышей. [37]
Эхолокационные летучие мыши генерируют ультразвук через гортань и издают звук через открытый рот или, гораздо реже, через нос. [38] Последнее наиболее выражено у подковоносов ( Rhinolophus spp. ). Эхолокационные сигналы летучих мышей варьируются по частоте от 14 000 до более чем 100 000 Гц, в основном за пределами диапазона человеческого уха (типичный диапазон человеческого слуха считается от 20 Гц до 20 000 Гц). Летучие мыши могут оценивать высоту целей, интерпретируя интерференционные картины, вызванные эхом, отражающимся от козелка , лоскута кожи во внешнем ухе. [39]
Отдельные виды летучих мышей эхолокируют в определенных диапазонах частот, которые подходят их среде обитания и типам добычи. Иногда это использовалось исследователями для идентификации летучих мышей, летающих в определенной области, просто записывая их крики с помощью ультразвуковых регистраторов, известных как «детекторы летучих мышей». Однако крики эхолокации не всегда являются видоспецифичными, и некоторые летучие мыши перекрываются по типу используемых ими криков, поэтому записи криков эхолокации не могут быть использованы для идентификации всех летучих мышей. Исследователи в нескольких странах разработали «библиотеки криков летучих мышей», которые содержат записи «эталонных криков» местных видов летучих мышей для помощи в идентификации. [40] [41] [42]
При поиске добычи они издают звуки с низкой частотой (10–20 щелчков в секунду). Во время фазы поиска эмиссия звука связана с дыханием, которое, в свою очередь, связано с взмахами крыльев. Эта связь, по-видимому, значительно экономит энергию, поскольку для летающих летучих мышей эхолокация не требует дополнительных энергетических затрат. [43] После обнаружения потенциальной добычи летучие мыши, использующие эхолокацию, увеличивают частоту импульсов, заканчивая конечным жужжанием , со скоростью до 200 щелчков в секунду. Во время приближения к обнаруженной цели продолжительность звуков постепенно уменьшается, как и энергия звука. [44]
Летучие мыши появились в начале эпохи эоцена , около 64 млн лет назад . Yangochiroptera появились около 55 млн лет назад, а Rhinolophoidea около 52 млн лет назад. [45]
Существует две гипотезы об эволюции эхолокации у летучих мышей. Первая предполагает, что ларингеальная эхолокация развивалась дважды или более у рукокрылых, по крайней мере один раз у Yangochiroptera и по крайней мере один раз у подковоносов (Rhinolophidae): [46]
Вторая теория предполагает, что гортанная эхолокация имела единое происхождение у Chiroptera, то есть была базовой для этой группы и впоследствии была утрачена в семействе Pteropodidae . [47] Позднее род Rousettus в семействе Pteropodidae развил другой механизм эхолокации, использующий систему щелчков языком: [48]
Эхолокационные летучие мыши занимают разнообразные экологические условия; их можно встретить живущими в таких разных средах, как Европа и Мадагаскар , и охотящимися за такими разными источниками пищи, как насекомые, лягушки, нектар, фрукты и кровь. Характеристики эхолокационного сигнала адаптированы к конкретной среде, охотничьему поведению и источнику пищи конкретной летучей мыши. Адаптация эхолокационных сигналов к экологическим факторам ограничена филогенетическими отношениями летучих мышей, что приводит к процессу, известному как спуск с модификацией, и приводит к разнообразию рукокрылых сегодня. [29] [32] [31] Летучие мыши могут непреднамеренно глушить друг друга, и в некоторых ситуациях они могут прекратить кричать, чтобы избежать глушения. [49]
Летающие насекомые являются обычным источником пищи для летучих мышей, использующих эхолокацию, а некоторые насекомые (в частности, мотыльки) могут слышать крики хищных летучих мышей. Однако эволюция органов слуха у мотыльков предшествовала появлению летучих мышей, поэтому, хотя многие мотыльки и слушают приближающуюся эхолокацию летучих мышей, их уши изначально не развивались в ответ на селективное давление со стороны летучих мышей. [50] Эти адаптации мотыльков обеспечивают селективное давление на летучих мышей, чтобы улучшить их системы охоты на насекомых, и этот цикл достигает кульминации в « эволюционной гонке вооружений » между мотыльками и летучими мышами. [51] [52]
Поскольку летучие мыши используют эхолокацию для ориентации и обнаружения объектов, их слуховые системы адаптированы для этой цели, высокоспециализированные для восприятия и интерпретации стереотипных эхолокационных сигналов, характерных для их собственного вида. Эта специализация очевидна от внутреннего уха до самых высоких уровней обработки информации в слуховой коре. [53]
У летучих мышей CF и FM есть специализированные внутренние уши, которые позволяют им слышать звуки в ультразвуковом диапазоне, далеко за пределами диапазона человеческого слуха. Хотя в большинстве других аспектов слуховые органы летучих мышей похожи на органы большинства других млекопитающих, некоторые летучие мыши ( подковоносы , Rhinolophus spp. и усатые летучие мыши , Pteronotus parnelii ) с компонентом постоянной частоты (CF) в их крике (известные как летучие мыши с высоким рабочим циклом) имеют несколько дополнительных адаптаций для обнаружения преобладающей частоты (и гармоник) вокализации CF. Они включают узкую частотную «настройку» органов внутреннего уха, с особенно большой областью, реагирующей на частоту возвращающихся эхо-сигналов летучей мыши. [33]
Базилярная мембрана внутри улитки содержит первую из этих специализаций для обработки эхо-информации. У летучих мышей, которые используют сигналы CF, участок мембраны, который реагирует на частоту возвращающихся эхо-сигналов, намного больше, чем область ответа для любой другой частоты. Например, у большого подковоноса, Rhinolophus ferrumequinum , есть непропорционально удлиненный и утолщенный участок мембраны, который реагирует на звуки около 83 кГц, постоянной частоты эха, производимого криком летучей мыши. Эта область высокой чувствительности к определенному узкому диапазону частот известна как « акустическая ямка ». [54]
Эхолокирующие летучие мыши имеют кохлеарные волоски, которые особенно устойчивы к интенсивному шуму. Кохлеарные волосковые клетки необходимы для чувствительности слуха и могут быть повреждены интенсивным шумом. Поскольку летучие мыши регулярно подвергаются воздействию интенсивного шума посредством эхолокации, устойчивость к деградации под воздействием интенсивного шума необходима. [55]
Далее по слуховому пути движение базилярной мембраны приводит к стимуляции первичных слуховых нейронов. Многие из этих нейронов специально «настроены» (реагируют наиболее сильно) на узкий диапазон частот возвращающихся эхо-сигналов CF-звонков. Из-за большого размера акустической ямки число нейронов, реагирующих на эту область, и, следовательно, на частоту эха, особенно велико. [56]
В нижнем холмике , структуре в среднем мозге летучей мыши, информация из нижних отделов слухового пути обработки интегрируется и отправляется в слуховую кору. Как показали Джордж Поллак и другие в серии статей в 1977 году, интернейроны в этой области имеют очень высокий уровень чувствительности к разнице во времени, поскольку задержка во времени между призывом и возвращающимся эхом сообщает летучей мыши ее расстояние от целевого объекта. В то время как большинство нейронов быстрее реагируют на более сильные стимулы, нейроны холмика сохраняют точность синхронизации даже при изменении интенсивности сигнала. [57] Эти интернейроны специализируются на чувствительности ко времени несколькими способами. Во-первых, при активации они обычно реагируют только одним или двумя потенциалами действия . Эта короткая продолжительность ответа позволяет их потенциалам действия давать конкретное указание на момент поступления стимула и точно реагировать на стимулы, которые происходят близко друг к другу по времени. Нейроны имеют очень низкий порог активации — они быстро реагируют даже на слабые стимулы. Наконец, для сигналов FM каждый интернейрон настроен на определенную частоту в пределах развертки, а также на ту же частоту в последующем эхе. На этом уровне также существует специализация для компонента CF вызова. Высокая доля нейронов, реагирующих на частоту акустической ямки, фактически увеличивается на этом уровне. [57]
Слуховая кора у летучих мышей довольно большая по сравнению с другими млекопитающими. [58] Различные характеристики звука обрабатываются различными областями коры, каждая из которых предоставляет различную информацию о местоположении или движении целевого объекта. Большинство существующих исследований по обработке информации в слуховой коре летучих мышей были проведены Нобуо Суга на усатой летучей мыши, Pteronotus parnellii . Этот крик летучей мыши имеет как тон CF, так и компоненты FM-развертки. [59] [60]
Суга и его коллеги показали, что кора содержит ряд «карт» слуховой информации, каждая из которых организована систематически на основе характеристик звука, таких как частота и амплитуда . Нейроны в этих областях реагируют только на определенную комбинацию частоты и времени (задержка звука-эха) и известны как нейроны, чувствительные к комбинации. [59] [60]
Систематически организованные карты в слуховой коре реагируют на различные аспекты эхо-сигнала, такие как его задержка и скорость. Эти области состоят из «комбинационно-чувствительных» нейронов, которым требуется по крайней мере два определенных стимула для получения ответа. Нейроны систематически различаются по картам, которые организованы по акустическим характеристикам звука и могут быть двумерными. Различные характеристики вызова и его эха используются летучей мышью для определения важных характеристик своей добычи. Карты включают: [59] [60]


Биосонар ценен как для зубатых китов (подотряд Odontoceti ), включая дельфинов , морских свиней , речных дельфинов , косаток и кашалотов , так и для усатых китов (подотряд Mysticeti ), включая гладких китов , гренландских китов , карликовых китов , а также серых китов и полосатиков , поскольку они живут в подводной среде обитания, которая имеет благоприятные акустические характеристики и где зрение часто крайне ограничено в диапазоне из-за поглощения или мутности . [62] Зубатые киты, как правило, способны слышать звуки на ультразвуковых частотах, в то время как усатые киты слышат звуки в инфразвуковом частотном режиме. [63]
Эволюция китообразных состояла из трех основных радиаций . На протяжении среднего и позднего эоцена (49–31,5 млн лет назад) археоцеты , примитивные зубатые китообразные, которые произошли от наземных млекопитающих, были единственными китообразными. [64] [65] Они не обладали эхолокацией, но имели слегка адаптированный подводный слух. [66] К концу среднего эоцена акустически изолированные слуховые косточки эволюционировали, чтобы дать базилозавридным археоцетам направленный подводный слух на низких и средних частотах. [67] С вымиранием археоцетов в начале олигоцена ( 33,9–23 млн лет назад) во второй радиации развились две новые линии. Ранние усатые киты (усатые киты) и зубатые киты появились в среднем олигоцене в Новой Зеландии. [65] Современные зубатые киты являются монофилетическими (единая эволюционная группа), но эхолокация развивалась дважды, конвергентно: один раз у Xenorophus , олигоценового стволового зубата, и один раз у корончатых зубатов. [36]
Физическая перестройка океанов сыграла свою роль в эволюции эхолокации. Глобальное похолодание на границе эоцена и олигоцена вызвало изменение от парникового к ледниковому миру . Тектонические отверстия создали Южный океан со свободно текущим Антарктическим циркумполярным течением . [66] [67] [68] Эти события способствовали отбору на способность находить и захватывать добычу в мутных речных водах, что позволило зубатым китам вторгаться и кормиться на глубинах ниже фотической зоны . В частности, эхолокация ниже фотической зоны могла быть адаптацией к хищничеству у дневных мигрирующих головоногих моллюсков . [67] [69] Семейство Delphinidae (дельфины) разнообразилось в неогене (23–2,6 миллиона лет назад), развив чрезвычайно специализированную эхолокацию. [70] [66]
Четыре белка играют важную роль в эхолокации зубатых китов. Престин , двигательный белок наружных волосковых клеток внутреннего уха улитки млекопитающих , связан с чувствительностью слуха. [71] Он претерпел два четких эпизода ускоренной эволюции у китообразных. [71] Первый связан с дивергенцией зубатых китов, когда впервые развилась эхолокация, а второй — с увеличением частоты эхолокации среди дельфинов. Tmc1 и Pjvk — это белки, связанные с чувствительностью слуха: Tmc1 связан с развитием волосковых клеток и высокочастотным слухом, а Pjvk — с функцией волосковых клеток. [72] Молекулярная эволюция Tmc1 и Pjvk указывает на положительный отбор для эхолокации у зубатых китов. [72] Cldn14 , член белков плотных контактов, которые образуют барьеры между клетками внутреннего уха, демонстрирует ту же эволюционную модель, что и Престин. [73] Два события эволюции белков, для Prestin и Cldn14, произошли в то же время, что и тектоническое открытие пролива Дрейка (34–31 млн лет назад) и рост антарктического льда в период климатического перехода в среднем миоцене (14 млн лет назад), при этом расхождение зубатых китов и усатых китов произошло в первом случае, а видообразование Delphinidae — во втором. [68]
Эволюция двух черепных структур может быть связана с эхолокацией. Телескопирование черепа (перекрытие лобных и верхнечелюстных костей и смещение ноздрей назад [74] ) впервые развилось у ксенорофид . Оно далее эволюционировало у стволовых зубатых китов, достигнув полного телескопирования черепа у коронных зубатых китов. [75] Движение ноздрей могло обеспечить более крупный носовой аппарат и дыню для эхолокации. [75] Это изменение произошло после расхождения неокетов от базилозавридов. [76] Первый сдвиг в сторону асимметрии черепа произошел в раннем олигоцене, до появления ксенорофидов. [76] Ископаемое ксенорофида ( Cotylocara macei ) имеет асимметрию черепа и показывает другие показатели эхолокации. [77] Однако у базальных ксенорофидов асимметрия черепа отсутствует, что указывает на то, что она, вероятно, развивалась дважды. [76] Современные зубатые киты имеют асимметричные носолицевые области; как правило, срединная плоскость смещена влево, а структуры справа больше. [77] Как краниальное телескопирование, так и асимметрия, вероятно, связаны с производством звука для эхолокации. [75]
Тринадцать видов современных зубатых китов конвергентно развили узкополосную высокочастотную (NBHF) эхолокацию в четырех отдельных событиях. Эти виды включают семейства Kogiidae (карликовые кашалоты) и Phocoenidae (морские свиньи), а также некоторые виды рода Lagenorhynchus , все Cephalorhynchus и дельфина Ла-Платы . Считается, что NBHF развился как средство уклонения от хищников; виды, продуцирующие NBHF, малы по сравнению с другими зубатыми китами, что делает их пригодной добычей для крупных видов, таких как косатка . Однако, поскольку три из этих групп развили NBHF до появления косатки, движущей силой развития NBHF должно было быть хищничество со стороны других древних хищных зубатых китов, а не хищничество со стороны косатки. Косатки и, предположительно, древние хищные зубатые киты, такие как акрофизетер , не способны слышать частоты выше 100 кГц. [78]
Другой причиной вариации эхолокации является среда обитания. Для всех систем сонара ограничивающим фактором, определяющим, будет ли обнаружено возвращающееся эхо, является отношение эха к шуму (ENR). ENR определяется уровнем излучаемого источника (SL) плюс сила цели, за вычетом потерь при двусторонней передаче (поглощение и распространение) и полученного шума. [79] Животные будут адаптироваться либо к максимальному увеличению дальности в условиях ограниченного шума (увеличение уровня источника), либо к уменьшению шумового шума в мелководной и/или замусоренной среде обитания (уменьшение уровня источника). В замусоренных средах обитания, таких как прибрежные районы, диапазоны добычи меньше, и такие виды, как дельфин Коммерсона ( Cephalorhynchus commersonii ), имеют пониженные уровни источника, чтобы лучше соответствовать своей среде. [79]
Зубатые киты издают сфокусированный луч высокочастотных щелчков в направлении, куда указывает их голова. Звуки генерируются путем прохождения воздуха из костных ноздрей через фонические губы . Эти звуки отражаются плотной вогнутой костью черепа и воздушным мешком у его основания. Сфокусированный луч модулируется большим жировым органом, известным как дыня. Он действует как акустическая линза, поскольку состоит из липидов различной плотности. Большинство зубатых китов используют щелчки в серии или последовательности щелчков для эхолокации, в то время как кашалот может производить щелчки по отдельности. Свистки зубатых китов, по-видимому, не используются для эхолокации. Различные скорости производства щелчков в последовательности щелчков вызывают знакомые лай, визги и рычание дельфина -афалины . Последовательность щелчков с частотой повторения более 600 в секунду называется пачкой импульсов. У дельфинов-афалин слуховая реакция мозга различает отдельные щелчки с частотой до 600 в секунду, но дает ступенчатый ответ при более высокой частоте повторения. [80]
Было высказано предположение, что расположение зубов некоторых более мелких зубатых китов может быть адаптацией для эхолокации. [81] Например, зубы дельфина-бутылки не расположены симметрично, если смотреть с вертикальной плоскости. Эта асимметрия, возможно, может помочь в определении того, исходят ли эхо от его биосонара с одной или другой стороны; но это не было проверено экспериментально. [82]
Эхо принимается с использованием сложных жировых структур вокруг нижней челюсти в качестве основного пути приема, откуда они передаются в среднее ухо через непрерывное жировое тело. Боковой звук может быть получен через жировые лопасти, окружающие уши с плотностью, аналогичной плотности воды. Некоторые исследователи полагают, что когда они приближаются к интересующему объекту, они защищают себя от более громкого эха, приглушая издаваемый звук. Известно, что это происходит у летучих мышей, но здесь также снижается слуховая чувствительность вблизи цели. [83] [84]
_hunting_by_echolocation.JPG/1280px-Palawan_swiftlet_(Aerodramus_palawanensis)_hunting_by_echolocation.JPG)
Известно, что масляные птицы и некоторые виды саланганы используют относительно грубую форму эхолокации по сравнению с летучими мышами и дельфинами. Эти ночные птицы издают крики во время полета и используют их для навигации по деревьям и пещерам, где они живут. [85] [86]
Наземные млекопитающие , кроме летучих мышей, известные или предположительно обладающие эхолокацией, включают землеройок, [87] [88] [89] тенреков Мадагаскара , [ 90 ] китайских карликовых сонь , [ 91 ] и щелезубов . [92] Звуки землероек, в отличие от звуков летучих мышей, имеют низкую амплитуду, широкополосные, многогармонические и частотно-модулированные. [89] Они не содержат эхолокационных щелчков с реверберациями и, по-видимому, используются для простой пространственной ориентации на близком расстоянии. В отличие от летучих мышей, землеройки используют эхолокацию только для исследования своей среды обитания, а не для точного определения пищи. [89] Есть доказательства того, что ослепленные лабораторные крысы могут использовать эхолокацию для навигации в лабиринтах. [93]

Некоторые насекомые, на которых охотятся летучие мыши, обладают защитными приспособлениями от хищников , включая избегание хищников, [95] отклонение атаки, [94] и ультразвуковые щелчки, которые, по-видимому, выполняют функцию предупреждения, а не глушения эхолокации . [49] [96]
Тигровые моли ( Arctiidae ) разных видов (две трети протестированных видов) реагируют на имитацию атаки эхолокирующими летучими мышами, производя ускоряющуюся серию щелчков. Было показано, что вид Bertholdia trigona подавляет эхолокацию летучих мышей: при столкновении с наивными большими коричневыми летучими мышами ультразвук был немедленно и последовательно эффективен для предотвращения нападения летучих мышей. Летучие мыши контактировали с молчаливыми контрольными молями на 400% чаще, чем с B. trigona . [97]
Ультразвук мотылька также может функционировать, чтобы напугать летучую мышь (тактика блефа), предупредить летучую мышь, что моль неприятна (честная сигнализация, апосематизм ), или имитировать химически защищенные виды. Было показано, что и апосематизм, и мимикрия дают преимущество в выживании против нападения летучих мышей. [98] [99]
Большая восковая моль ( Galleria mellonella ) предпринимает действия по избеганию хищников, такие как падение, петляние и замирание, когда обнаруживает ультразвуковые волны, что указывает на то, что она может как обнаруживать, так и различать ультразвуковые частоты, используемые хищниками, и сигналы от других членов их вида. [95] Некоторые члены семейства моли Saturniidae , в которое входят гигантские шелкопряды, имеют длинные хвосты на задних крыльях, особенно те, что относятся к подгруппам Attacini и Arsenurinae . Хвосты колеблются в полете, создавая эхо, которое отклоняет атаку охотящейся летучей мыши от тела моли к хвостам. Вид Argema mimosae (африканская лунная моль), у которого особенно длинные хвосты, был наиболее склонен избегать захвата. [94]
Со стр. 140: Из этих экспериментов автор делает вывод: … что орган слуха, по-видимому, дополняет орган зрения при обнаружении тел и снабжает этих животных различными ощущениями для управления их полетом и позволяет им избегать тех препятствий, которые могут возникнуть.
Летучие мыши и дельфины известны своей способностью использовать эхолокацию. ... некоторые слепые люди научились делать то же самое ...
