
Пелагические рыбы обитают в пелагической зоне океанских или озерных вод, не находясь ни у дна, ни у берега, в отличие от демерсальных рыб , обитающих на дне или вблизи него, и рифовых рыб , связанных с коралловыми рифами . [1]
Морская пелагическая среда является крупнейшей водной средой обитания на Земле, занимая 1370 миллионов кубических километров (330 миллионов кубических миль), и является средой обитания для 11% известных видов рыб . Средняя глубина океанов составляет 4000 метров (2,5 мили) . Около 98% общего объема воды находится на глубине ниже 100 метров (330 футов), а 75% - ниже 1000 метров (3300 футов). [2]
Морских пелагических рыб можно разделить на прибрежных (прибрежных) и океанических (шельфовых) рыб. Прибрежные пелагические рыбы обитают в относительно мелких и освещенных солнцем водах над континентальным шельфом , в то время как океанические пелагические рыбы обитают в обширных и глубоких водах за пределами континентального шельфа (хотя они также могут плавать вблизи берега). [3] [4]
Размер пелагических рыб варьируется от мелких прибрежных кормовых рыб , таких как сельдь и сардины , до крупных океанических рыб-хищников , таких как голубой тунец и океанические акулы . [1] Обычно это ловкие пловцы с обтекаемым телом, способные совершать длительные миграции на большие расстояния . Многие пелагические рыбы плавают стаями весом в сотни тонн. Другие, такие как большая океанская рыба-луна , ведут одиночный образ жизни. [1] Есть также пресноводные пелагические рыбы в некоторых крупных озерах, таких как сардины в озере Танганьика . [5]
Эпипелагические рыбы населяют эпипелагическую зону , самый верхний слой водной толщи , простирающийся от уровня моря до 200 м (660 футов). Ее также называют поверхностными водами или солнечной зоной и включают в себя фотозону . Фотическая зона определяется как поверхностные воды до глубины, где солнечный свет ослабляется до 1% от поверхностного значения. Эта глубина зависит от того, насколько мутна вода, но в чистой воде может достигать 200 м (660 футов), что соответствует эпипелагиали. Фотическая зона обеспечивает достаточно света для фотосинтеза фитопланктона . [6]
Обширная среда обитания для большинства пелагических рыб, эпипелагическая зона хорошо освещена, поэтому хищники-зрители могут использовать свое зрение, обычно хорошо перемешивается и насыщается кислородом благодаря воздействию волн и может быть хорошей средой обитания для роста водорослей . Однако это почти безликая среда обитания. Отсутствие разнообразия среды обитания приводит к отсутствию видового разнообразия , поэтому в этой зоне обитает менее 2% известных в мире видов рыб. На большей части зоны не хватает питательных веществ для поддержания рыб, поэтому эпипелагические рыбы, как правило, встречаются в прибрежных водах над континентальными шельфами , где поверхностный сток может обеспечить питательные вещества, или в тех частях океана, где апвеллинг перемещает питательные вещества в этот район. [6]
Эпипелагических рыб можно условно разделить на мелких кормовых рыб и более крупных хищных рыб , питающихся ими. Кормовой косяк рыб и фильтрационный корм планктона . Большинство эпипелагических рыб имеют обтекаемые тела, способные к длительным миграциям . В целом хищные и кормовые рыбы имеют одинаковые морфологические особенности. Рыбы-хищники обычно веретенообразные, с большой пастью, гладким телом и глубоко раздвоенным хвостом. Многие используют зрение, чтобы охотиться на зоопланктон или более мелкую рыбу, в то время как другие фильтруют корм планктоном.

Большинство эпипелагических хищных рыб и их более мелкая добыча затенены серебристыми цветами, которые уменьшают видимость из-за рассеивания падающего света. [6] Серебрение достигается с помощью отражающей рыбьей чешуи , которая действует как маленькое зеркало. Это может дать эффект прозрачности. На средних глубинах в море свет падает сверху, поэтому зеркало, ориентированное вертикально, делает животных, например рыб, невидимыми сбоку. [7]
В более мелких эпипелагических водах зеркала должны отражать смесь длин волн, и рыба, соответственно, имеет стопки кристаллов с разным расстоянием между ними. Еще одна сложность для рыб с закругленным в поперечном сечении телом заключается в том, что зеркала будут неэффективны, если положить их ровно на кожу, поскольку они не смогут отражать горизонтально. Общий зеркальный эффект достигается за счет множества небольших отражателей, ориентированных вертикально. [7]
Хотя число видов ограничено, эпипелагические рыбы многочисленны. Недостаток разнообразия они компенсируют численностью. Кормовая рыба встречается в огромных количествах, и крупная рыба, которая охотится на нее, часто считается основной пищевой рыбой . Как группа эпипелагических рыб образуют наиболее ценные промысловые запасы в мире. [6]
Многие кормовые рыбы являются факультативными хищниками, которые могут собирать отдельных копепод или личинок рыб из толщи воды, а затем переходить на фильтрующее питание фитопланктоном , когда это дает лучшие энергетические результаты. Рыбы, питающиеся фильтратором, обычно используют длинные тонкие жаберные тычинки для выделения мелких организмов из толщи воды. Некоторые из крупнейших эпипелагических рыб, такие как гигантская акула и китовая акула , являются фильтраторами, как и некоторые из самых мелких, например взрослые кильки и анчоусы . [8]
Исключительно прозрачные океанские воды содержат мало пищи. Области с высокой продуктивностью имеют тенденцию быть несколько мутными из-за цветения планктона . Они привлекают фильтраторов, питающихся планктоном, которые, в свою очередь, привлекают высших хищников. Ловля тунца оптимальна, когда мутность воды, измеряемая по максимальной глубине, на которой можно увидеть диск секки в солнечный день, составляет от 15 до 35 метров. [9]
Эпипелагические рыбы очарованы плавающими объектами. Они собираются в значительных количествах вокруг таких объектов, как дрейфующие обломки, плоты, медузы и плавающие водоросли. Кажется, что объекты обеспечивают «визуальный стимул в оптической пустоте». [10] Плавающие объекты могут служить убежищем молоди рыб от хищников. Обилие дрейфующих водорослей или медуз может привести к значительному увеличению выживаемости некоторых молодых видов. [11]
Многие прибрежные молодые особи используют морские водоросли в качестве убежища и пищи, которую можно получить от беспозвоночных и других связанных с ними рыб. Дрейфующие морские водоросли, особенно пелагические саргассумы , обеспечивают нишу среды обитания с собственным укрытием и пищей и даже поддерживают собственную уникальную фауну, такую как рыба саргассум . [8] Одно исследование, проведенное у берегов Флориды, выявило 54 вида из 23 семей, живущих в обломках циновок Саргассума . [12] Молодь рыб также использует медуз в качестве убежища и еды, хотя медузы могут охотиться на мелкую рыбу. [13]
Подвижные океанические виды, такие как тунец, можно поймать, путешествуя на большие расстояния на больших рыболовных судах . Более простая альтернатива — использовать пристрастие рыб к плавающим объектам. Когда рыбаки используют такие предметы, их называют устройствами для сбора рыбы (УАУ). FAD — это закрепленные на якоре плоты или объекты любого типа, плавающие на поверхности или чуть ниже нее. Рыбаки Тихого и Индийского океанов устанавливают вокруг тропических островов плавучие ФАДы, собранные из разного мусора, а затем с помощью кошельковых неводов ловят привлеченную ими рыбу. [14]
Исследование с использованием сонара во Французской Полинезии показало, что большие косяки молоди большеглазого и желтоперого тунца скопились ближе всего к устройствам, на расстоянии от 10 до 50 метров. Дальше, на расстоянии 50–150 м, обитала менее плотная группа более крупных желтоперых и длинноперых тунцов . Еще дальше, на высоте 500 м, находилась рассредоточенная группа разных крупных взрослых тунцов. Распределение и плотность этих групп были переменными и перекрывались. FAD также использовались другими рыбами, и скопления рассеивались в темноте. [15]
Более крупные рыбы, даже хищные рыбы, такие как большая барракуда , часто привлекают свиту мелких рыб, которые стратегически безопасно сопровождают их. Скиндайверы , которые остаются в воде в течение длительного времени, также часто привлекают стайку рыб: более мелкие рыбы подходят близко, а более крупные наблюдают с большего расстояния. Морские черепахи , служащие мобильным укрытием для мелкой рыбы, могут быть случайно пронзены рыбой-меч, пытающейся поймать рыбу. [16]

Прибрежные рыбы (также называемые неритическими или прибрежными рыбами) обитают в водах у побережья и над континентальным шельфом . Поскольку глубина континентального шельфа обычно составляет менее 200 метров, из этого следует, что прибрежные рыбы, не являющиеся демерсальными рыбами, обычно являются эпипелагическими рыбами, населяющими освещенную солнцем эпипелагическую зону. [2]
Прибрежные эпипелагические рыбы являются одними из самых многочисленных в мире. К ним относятся кормовая рыба, а также хищные рыбы, которые ими питаются. Кормовая рыба процветает в тех прибрежных водах, где высокая продуктивность обусловлена апвеллингом и стоком питательных веществ с береговой линии. Некоторые из них являются частичными обитателями и нерестятся в ручьях, устьях рек и заливах, но большинство завершает свой жизненный цикл в этой зоне. [8]
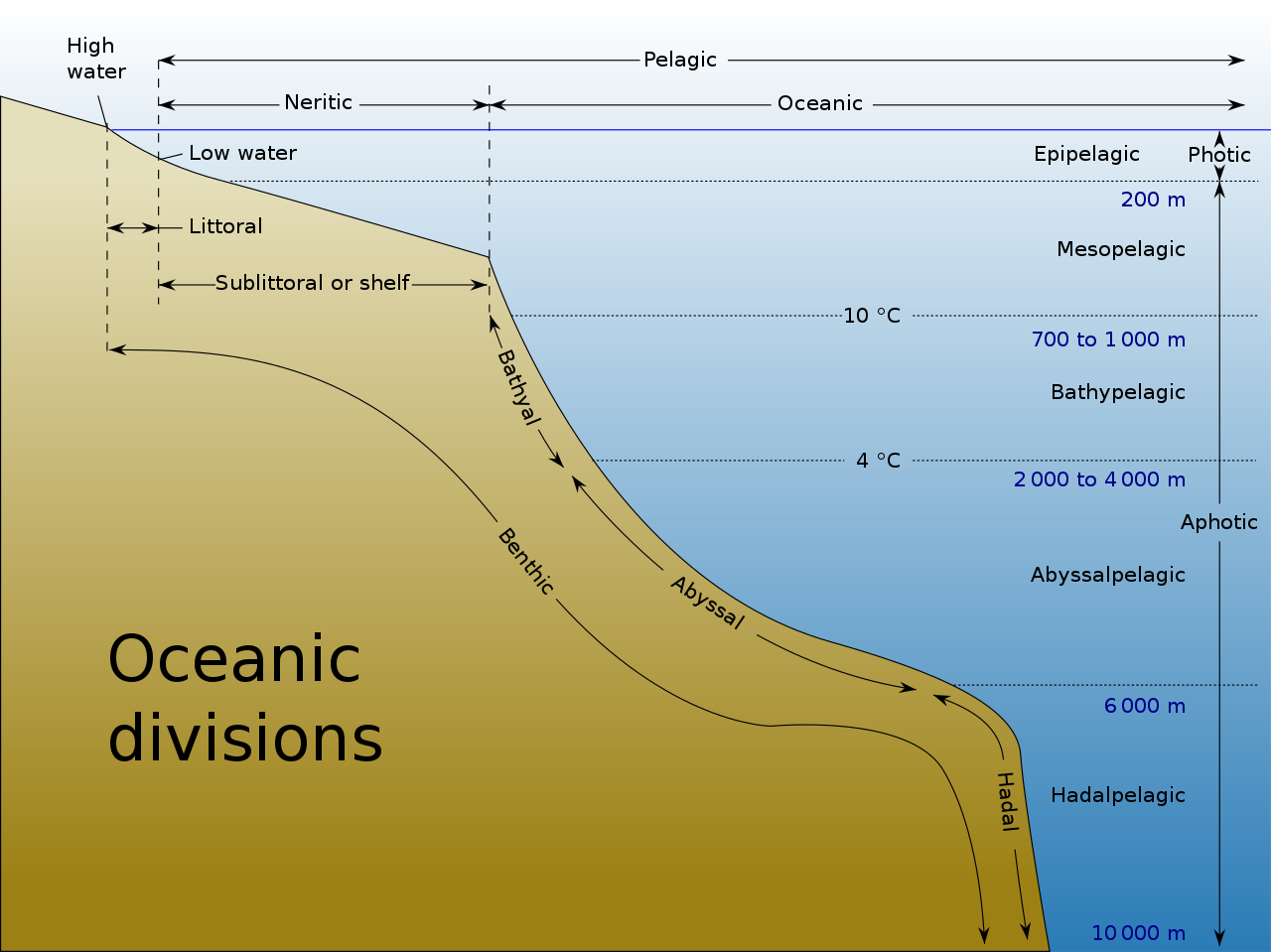
Океанические рыбы (также называемые рыбами открытого океана или прибрежными рыбами) обитают в водах, не расположенных над континентальным шельфом. Океаническую рыбу можно противопоставить прибрежным рыбам , которые обитают над континентальным шельфом. Однако эти два типа не являются взаимоисключающими, поскольку между прибрежными и океаническими районами нет четких границ, и многие эпипелагические рыбы перемещаются между прибрежными и океаническими водами, особенно на разных стадиях своего жизненного цикла. [8]
Океанические эпипелагические рыбы могут быть настоящими, частичными и случайными обитателями. Настоящие жители всю свою жизнь проживают в открытом океане. Лишь немногие виды являются настоящими обитателями, например, тунец , марлиновые , летучие рыбы , сайры , лоцманы , реморы , дельфины , океанские акулы и океанские солнечные рыбы . Большинство этих видов мигрируют взад и вперед через открытые океаны, редко пересекая континентальные шельфы. У некоторых истинных жителей это место ассоциируется с дрейфующими медузами или водорослями. [8]
Частичные обитатели встречаются в трех группах: виды, обитающие в зоне только в молодом возрасте (дрейфующие вместе с медузами и водорослями); виды, обитающие в зоне только во взрослом возрасте (лосось, летучая рыба, дельфины и китовые акулы); и глубоководные виды, которые совершают ночные миграции в поверхностные воды (например, рыба-фонарь ). [8] Случайные обитатели происходят время от времени, когда взрослые и молодые особи видов из других сред случайно заносятся в зону течениями. [8]
.gif/1280px-Protomyctophum_subparallelum_(no_common_name).gif)

В глубоком океане воды простираются намного ниже эпипелагиали и служат средой обитания самых разных типов пелагических рыб, приспособленных к жизни в этих более глубоких зонах. [2]
На глубокой воде морской снег представляет собой непрерывный дождь преимущественно органического детрита , выпадающего из верхних слоев водной толщи. Его происхождение лежит в деятельности продуктивной фотозоны . Морской снег включает мертвый или умирающий планктон , простейших ( диатомовые водоросли ), фекалии, песок, сажу и другую неорганическую пыль. «Снежинки» со временем растут и могут достигать нескольких сантиметров в диаметре, путешествуя неделями, прежде чем достичь дна океана. Однако большинство органических компонентов морского снега потребляются микробами , зоопланктоном и другими животными-фильтраторами в течение первых 1000 метров их пути, то есть в пределах эпипелагиали. Таким образом, морской снег можно считать основой глубоководных мезопелагических и донных экосистем : поскольку солнечный свет не может достичь их, глубоководные организмы в значительной степени полагаются на морской снег как на источник энергии.
Некоторые глубоководные пелагические группы, такие как семейства фонарей , риджхедов , морских топориков и легких рыб , иногда называют псевдокеаническими , потому что вместо равномерного распределения в открытой воде они встречаются в значительно более высоких количествах вокруг структурных оазисов, особенно подводных гор , и над материковыми склонами . Это явление объясняется также обилием видов добычи, которых также привлекают структуры.
Рыбы в различных пелагических и глубоководных бентических зонах имеют физическое строение и поведение, заметно отличающееся друг от друга. Группы сосуществующих видов в каждой зоне, по-видимому, действуют схожим образом, например, небольшие мезопелагические вертикально мигрирующие питающиеся планктоном, батипелагические удильщики и глубоководные донные гремуучие хвосты . [17]
Виды с лучами и колючими плавниками редки среди глубоководных рыб, что позволяет предположить, что глубоководные рыбы являются древними и настолько хорошо адаптированы к окружающей среде, что вторжения более современных рыб оказались безуспешными. [18] Несколько лучевых плавников, которые существуют, в основном встречаются у Beryciformes и Lampriformes , которые также являются древними формами. Большинство глубоководных пелагических рыб принадлежат к своим отрядам, что позволяет предположить длительную эволюцию в глубоководной среде. Напротив, глубоководные донные виды относятся к отрядам, которые включают множество родственных мелководных рыб. [19]
Многие виды ежедневно перемещаются между зонами в ходе вертикальных миграций. В следующей таблице они перечислены в средней или более глубокой зоне, где они регулярно встречаются.


Ниже эпипелагиали условия быстро меняются. На высоте от 200 до примерно 1000 метров свет продолжает тускнеть, пока темнота не становится почти полной. Температура падает через термоклин до температуры от 4 ° C (39 ° F) до 8 ° C (46 ° F). Это сумеречная или мезопелагическая зона. Давление продолжает расти со скоростью одна атмосфера каждые 10 метров, в то время как концентрация питательных веществ падает вместе с растворенным кислородом и скоростью циркуляции воды. [2] [21]
Операторы гидролокаторов, использовавшие гидроакустические технологии, разработанные во время Второй мировой войны, были озадачены тем, что выглядело ложным морским дном глубиной 300–500 метров днем и меньшей глубиной ночью. Оказалось, что это произошло благодаря миллионам морских организмов, особенно мелких мезопелагических рыб, плавательные пузыри которых отражали сигналы сонара.
Мезопелагические организмы мигрируют на мелководье в сумерках, чтобы питаться планктоном. Слой глубже, когда луна светит, и может подниматься выше, когда небо темное. Это явление стало известно как глубокий слой рассеяния . [22]
Большинство мезопелагических рыб совершают ежедневные вертикальные миграции , перемещаясь каждую ночь в эпипелагическую зону, часто следуя за подобными миграциями зоопланктона, и возвращаясь на глубину в целях безопасности в течение дня. [21] [2] [23] Эти вертикальные миграции происходят на расстояние в сотни метров.
У этих рыб мускулистое тело, окостеневшие кости, чешуя, хорошо развитые жабры и центральная нервная система, а также большие сердце и почки. Мезопелагические питатели планктона имеют маленькие рты с тонкими жаберными тычинками , тогда как у рыбоядных есть рот большего размера и более грубые жаберные тычинки. [21] [2]
Вертикально мигрирующие рыбы имеют плавательный пузырь . [18] Рыба надувает плавательный пузырь, чтобы двигаться вверх. Учитывая высокие давления в мезопелагической зоне, это требует значительных энергетических затрат. По мере подъема рыбы воздух в плавательном пузыре должен уменьшаться, чтобы предотвратить разрыв плавательного пузыря. Для возвращения на глубину плавательный пузырь сдувается. [24] Миграция проходит через термоклин , где температура колеблется от 10 до 20 ° C, что демонстрирует значительную устойчивость к температуре. [25]
Мезопелагические рыбы приспособлены к активной жизни в условиях недостаточной освещенности. Большинство из них — зрительные хищники с большими глазами. У некоторых глубоководных рыб, таких как рыба-телескоп , глаза трубчатые с большими линзами и только палочковые клетки смотрят вверх. Они обеспечивают бинокулярное зрение и высокую чувствительность к слабым световым сигналам. [2] Эта адаптация обеспечивает улучшенное конечное зрение за счет бокового зрения и позволяет хищнику различать кальмаров , каракатиц и более мелких рыб, силуэты которых вырисовываются над ними. [21]
Мезопелагические рыбы обычно не имеют защитных шипов и используют цвет для маскировки . [21] Хищники, устраивающие засаду, бывают темными, черными или красными. Поскольку более длинные красные длины волн света не достигают морских глубин, красный цвет эффективно действует так же, как черный. Мигрирующие формы используют серебристые цвета с контрастными оттенками . На животе у них часто имеются фотофоры , излучающие слабый свет. Для хищника снизу, смотрящего вверх, эта биолюминесценция маскирует силуэт рыбы. Однако у некоторых из этих хищников есть желтые линзы, которые фильтруют окружающий свет (с недостатком красного), оставляя видимой биолюминесценцию. [26]

![Светоотражатель также является одной из немногих рыб, которые производят красную биолюминесценцию. Поскольку большая часть их жертв не воспринимает красный свет, это позволяет им охотиться с помощью практически невидимого луча света.[29]](http://upload.wikimedia.org/wikipedia/commons/thumb/b/b2/Malacosteus.JPG/1280px-Malacosteus.JPG)
Коричневорылая рыба- призрак — это разновидность бочкоглаза , и единственное известное позвоночное животное, которое использует зеркало, а не линзу, чтобы фокусировать изображение в глазах. [31] [32]
Отбор проб с помощью глубоководного траления показывает, что на долю фонарей приходится до 65% всей биомассы глубоководных рыб . [33] Действительно, рыбы-фонарики являются одними из наиболее широко распространенных, густонаселенных и разнообразных среди всех позвоночных животных , играя важную экологическую роль в качестве добычи для более крупных организмов. По оценкам, глобальная биомасса фонарей составляет 550–660 миллионов тонн , что в несколько раз превышает весь мировой вылов рыбы. Рыбы-фонарики также составляют большую часть биомассы, ответственной за глубокий рассеивающий слой мирового океана. Сонар отражается от миллионов плавательных пузырей рыб-фонарей , создавая впечатление ложного дна. [34]
В 2010 году кругосветная экспедиция Маласпины преодолела 60 000 км, проведя акустические наблюдения. В нем сообщается, что мезопелагическая биомасса составляет 10 миллиардов тонн и более (в 10 раз больше предыдущих оценок), что составляет около 90 процентов всей биомассы океанских рыб. [35] По состоянию на 2024 год оценки того, сколько углерода связывают эти рыбы, оставались весьма неопределенными. [36]
По состоянию на 2024 год мезопелагическая рыба не является основным промыслом. Первоначальные усилия в Исландии, Норвегии и Советском Союзе не привели к созданию коммерческой отрасли. Европейский Союз профинансировал проект MEESO по изучению численности и технологий рыболовства ключевых мезопелагических видов. На сегодняшний день не обнаружено рыб, которые бы понравились человеческому вкусу, поэтому промысловики вместо этого сосредоточились на рынках кормов для животных. [36]
Большеглазый тунец — это эпипелагический/мезопелагический вид, плотоядный и поедающий другую рыбу. Спутниковое мечение показало, что большеглазый тунец в дневное время часто проводит длительные периоды времени, путешествуя глубоко под поверхностью воды, иногда ныряя на глубину до 500 метров (1600 футов). Считается, что эти движения являются реакцией на вертикальные миграции хищных организмов в глубоком слое рассеяния .




Ниже мезопелагической зоны темно. Это полуночная или батипелагическая зона , простирающаяся от 1000 м до дна глубоководной донной зоны . Если вода исключительно глубокая, пелагическую зону ниже 4000 метров (2,5 мили) иногда называют нижней полуночной или абиссопелагической зоной .
Условия во всех этих зонах в некоторой степени однородны, темнота полная, давление сокрушительное, а температура, питательные вещества и уровни растворенного кислорода низкие. [2]
Батипелагические рыбы обладают особыми приспособлениями , позволяющими справляться с этими условиями: у них медленный метаболизм и неспециализированная диета, они готовы есть все, что попадается под руку. Они предпочитают сидеть и ждать еды, а не тратить энергию на ее поиски. Поведение батипелагических рыб можно противопоставить поведению мезопелагических рыб. Мезопелагические рыбы часто очень подвижны, тогда как батипелагические рыбы почти все являются хищниками, затаившимися в засаде, обычно затрачивающими мало энергии на движение. [44]
Доминирующими рыбами батипелагии являются щетинорот и удильщик ; Также распространены клыказуб , рыба-гадюка , кинжалозуб и барракудина . Эти рыбы маленькие, многие длиной около 10 см (3,9 дюйма), но немногие длиннее 25 см (9,8 дюйма). Большую часть времени они терпеливо ждут в толще воды, пока не появится добыча или ее не заманит их люминофор. То немногое энергии, которое имеется в батипелагической зоне, фильтруется сверху в виде детрита, фекалий и редких беспозвоночных или мезопелагических рыб. [44] Около 20% пищи, происходящей из эпипелагиали, попадает в мезопелагическую зону, [22] но только около 5% фильтруется в батипелагическую зону. [40]
Батипелагические рыбы ведут оседлый образ жизни и приспособлены к выработке минимума энергии в среде обитания с очень небольшим количеством пищи или доступной энергии, даже не солнечного света, а только биолюминесценции. Их тела удлиненные, со слабыми водянистыми мышцами и скелетными структурами. Поскольку большая часть рыбы состоит из воды, на этих глубинах они не сжимаются большим давлением. У них часто есть выдвижные шарнирные челюсти с загнутыми зубами. Они склизкие, без чешуи . Центральная нервная система ограничена боковой линией и обонятельной системой, глаза маленькие и могут не функционировать, а жабры , почки и сердце, а также плавательный пузырь маленькие или отсутствуют. [40] [45]
Это те же особенности, которые обнаружены у личинок рыб , что позволяет предположить, что в ходе своей эволюции батипелагические рыбы приобрели эти особенности посредством неотении . Как и в случае с личинками, эти особенности позволяют рыбе оставаться в воде с небольшими затратами энергии. [46]
Несмотря на свой свирепый вид, эти подводные твари в основном представляют собой миниатюрных рыбок со слабыми мышцами и слишком малы, чтобы представлять какую-либо угрозу для людей.
Плавательные пузыри глубоководных рыб либо отсутствуют, либо почти не функционируют, а батипелагические рыбы обычно не совершают вертикальных миграций. Наполнение мочевых пузырей при таком высоком давлении требует огромных затрат энергии. У некоторых глубоководных рыб есть плавательные пузыри, которые функционируют, пока они молоды и обитают в верхней эпипелагиали, но они увядают или наполняются жиром, когда рыбы спускаются к своей взрослой среде обитания. [47]
Наиболее важными сенсорными системами обычно являются внутреннее ухо , реагирующее на звук, и боковая линия , реагирующая на изменения давления воды. Обонятельная система также может быть важна для самцов, которые находят самок по запаху . [48] Батипелагические рыбы черные, а иногда и красные, с небольшим количеством фотофоров . Фотофоры обычно используются для соблазнения добычи или привлечения партнера. Поскольку еды очень мало, батипелагические хищники не избирательны в своих пищевых привычках, а хватают все, что оказывается достаточно близко. Они достигают этого за счет большого рта с острыми зубами для захвата крупной добычи и перекрывающихся жаберных тычинок , которые не позволяют проглоченной мелкой добыче ускользнуть. [45]
В этой зоне нелегко найти себе пару. Некоторые виды зависят от биолюминесценции . Другие являются гермафродитами , что удваивает их шансы на производство яйцеклеток и сперматозоидов при столкновении. [40] Самка удильщика выделяет феромоны , чтобы привлечь крошечных самцов. Когда самец находит ее, он кусает ее и уже не отпускает. Когда самец вида удильщиков Haplophryne mollis кусает кожу самки, он выделяет фермент , который переваривает кожу его рта и ее тела, соединяя пару до точки, где соединяются две системы кровообращения. Затем самец атрофируется, превращаясь в пару гонад . Этот крайний половой диморфизм гарантирует, что, когда самка будет готова к нересту, у нее сразу же появится партнер. [49]
В батипелагиали обитают многие формы животных, кроме рыб, такие как кальмары, крупные киты, осьминоги, губки, брахиоподы , морские звезды и морские ежи , но для рыб в этой зоне жить трудно.
.gif/1280px-Haplophryne_mollis_(female,_with_atrophied_male_attached).gif)

Демерсальные рыбы живут на дне моря или вблизи него. [54] Демерсальные рыбы обитают на морском дне в прибрежных районах континентального шельфа , а в открытом океане они встречаются вдоль внешней окраины континента на континентальном склоне и континентальном поднятии. Они обычно не встречаются на абиссопелагических или гадопелагических глубинах или на абиссальной равнине . Они занимают ряд морских участков, состоящих из ила, песка, гравия или камней. [54]
В глубоких водах рыбы демерсальной зоны активны и относительно многочисленны по сравнению с рыбами батипелагиали . [44]
Распространены крысохвосты и бротулы , а другие хорошо зарекомендовавшие себя семейства — это угри , бельдюги , миксины , зеленоглазки , нетопыри и пинагоры . [45]
Тело глубоководных донных рыб мускулистое, с хорошо развитыми органами. В этом отношении они ближе к мезопелагическим рыбам, чем к батопелагическим. В остальном они более изменчивы. Фотофоры обычно отсутствуют, глаза и плавательные пузыри варьируются от отсутствующих до хорошо развитых. Они различаются по размеру, причем нередко встречаются более крупные виды размером более одного метра.
Глубоководные донные рыбы обычно длинные и узкие. Многие из них представляют собой угри или имеют форму угрей. Это может быть связано с тем, что длинные тела имеют длинные боковые линии . Боковые линии улавливают низкочастотные звуки, а у некоторых донных рыб есть мышцы, издающие такие звуки для привлечения самок. [18] Запах также важен, о чем свидетельствует скорость, с которой донная рыба находит ловушки с наживкой .
Основной рацион глубоководных донных рыб — беспозвоночные глубоководного бентоса и падаль . Обоняние, осязание и чувствительность боковой линии, по-видимому, являются основными сенсорными устройствами для их обнаружения. [55]
Глубоководных донных рыб можно разделить на строго донных и бентопелагических рыб. Обычно строго донные рыбы имеют отрицательную плавучесть, тогда как бентопелагические рыбы имеют нейтральную плавучесть. Строго донные рыбы находятся в постоянном контакте со дном. Они либо подстерегают хищников из засады , либо активно перемещаются по дну в поисках пищи. [55]


Бентопелагические рыбы обитают в воде чуть выше дна, питаясь бентосом и бентопелагическим зоопланктоном . [56] Большинство кожных рыб являются бентопелагическими. [54]
Их можно разделить на дряблые и крепкие типы телосложения. Вялые бентопелагические рыбы похожи на батопелагических рыб: у них уменьшенная масса тела и низкий уровень метаболизма, они расходуют минимум энергии, когда лежат и ждут, чтобы устроить засаду на добычу . [57] Примером дряблой рыбы является обыкновенный угорь Acanthonus Armatus , [58] хищник с огромной головой и телом, состоящим на 90% из воды. У этой рыбы самые большие уши ( отолиты ) и самый маленький мозг по отношению к размеру тела среди всех известных позвоночных. [59]
Крепкие бентопелагические рыбы — мускулистые пловцы, которые активно бороздят дно в поисках добычи. Они могут жить вокруг таких объектов, как подводные горы , где есть сильные течения. [59] Примерами являются оранжевый большеголов и патагонский клыкач . Поскольку когда-то этих рыб было много, а их крепкие тела очень вкусны, эту рыбу стали ловить в коммерческих целях. [60] [61]
Бентические рыбы не являются пелагическими рыбами, но здесь они обсуждаются кратко, для полноты и контраста.
Некоторые рыбы не вписываются в приведенную выше классификацию. Например, семейство почти слепых рыб-пауков , обычное и широко распространенное, питается бентопелагическим зоопланктоном. Тем не менее, это строго донные рыбы, поскольку они остаются в контакте со дном. В их плавниках есть длинные лучи, которые они используют, чтобы «стоять» на дне, стоя лицом к течению, и захватывать проходящий мимо зоопланктон. [62]
Самая глубоководная из известных рыб, строго донная Abyssobrotula galatheae , похожая на угря и слепая, питается донными беспозвоночными. [63] [64]

На больших глубинах нехватка пищи и экстремальное давление ограничивают выживаемость рыб. Самая глубокая точка океана составляет около 11 000 метров (6,8 миль). Батипелагические рыбы обычно не встречаются на глубине ниже 3000 метров (1,9 мили). Наибольшая зарегистрированная глубина для донных рыб составляет 8370 м (5,20 миль). [67] Возможно, экстремальное давление мешает основным функциям ферментов. [40]
Бентические рыбы более разнообразны, и их, скорее всего, можно встретить на континентальном склоне , где есть разнообразие среды обитания и, зачастую, запасы пищи. Примерно 40% дна океана составляют абиссальные равнины , но эти плоские, безликие области покрыты осадками и в значительной степени лишены донной жизни ( бентоса ). Глубоководные донные рыбы с большей вероятностью обитают в каньонах или обнажениях скал среди равнин, где созданы сообщества беспозвоночных. Подводные горы ( подводные горы ) могут перехватывать глубоководные морские течения и вызывать продуктивные апвеллинги, поддерживающие донную рыбу. Подводные горные хребты могут разделять подводные регионы на разные экосистемы. [17]
Мелкие пелагические рыбы обычно являются кормовой рыбой , на которую охотятся более крупные пелагические рыбы и другие хищники. Кормовые рыбы -фильтраторы питаются планктоном и обычно имеют длину менее 10 сантиметров (3,9 дюйма). Они часто держатся вместе стаями и могут мигрировать на большие расстояния между нерестилищами и местами нагула. Они встречаются особенно в регионах апвеллинга вокруг северо-восточной Атлантики, у берегов Японии, а также у западных побережий Африки и Америки. Кормовая рыба, как правило, недолговечна, и ее запасы заметно колеблются с годами. [68]
Сельдь водится в Северном море и Северной Атлантике на глубине до 200 метров (660 футов). Важные промыслы сельди существовали в этих районах на протяжении веков. Сельдь разных размеров и темпов роста принадлежит разным популяциям, каждая из которых имеет свои пути миграции. При нересте самка производит от 20 000 до 50 000 икринок. После нереста сельдь обедняется жиром и мигрирует обратно в места нагула, богатые планктоном. [69] Вокруг Исландии традиционно вылавливали три отдельные популяции сельди. Эти акции рухнули в конце 1960-х годов, хотя две из них с тех пор восстановились. После коллапса Исландия обратилась к мойве , на которую сейчас приходится около половины общего улова Исландии. [70]
Путассу обитают в открытом океане и над материковым склоном на глубине от 100 до 1000 метров. Они следуют за вертикальными миграциями зоопланктона, которым питаются, ко дну днем и к поверхности ночью. [69] [71]
Традиционный промысел анчоусов и сардин также ведется в Тихом океане, Средиземноморье и юго-восточной Атлантике. [72] Ежегодный мировой вылов кормовой рыбы в последние годы составлял примерно 22 миллиона тонн, или четверть мирового улова.


Пелагические рыбы среднего размера включают тревалли , барракуду , летучую рыбу , скумбрия , махи-махи и прибрежную скумбрию. [1] Многие из этих рыб охотятся на кормовую рыбу, но на них, в свою очередь, охотятся еще более крупные пелагические рыбы. Почти все рыбы в той или иной степени являются хищными, и, за исключением высших хищников, различие между хищной рыбой и добычей или кормовой рыбой несколько искусственно. [73]
По всей Европе существуют три популяции прибрежной скумбрии . Одна популяция мигрирует в Северное море, другая остаётся в водах Ирландского моря , а третья популяция мигрирует на юг вдоль западного побережья Шотландии и Ирландии. Крейсерская скорость скумбрии составляет впечатляющие 10 километров в час. [69] [74]
Многие крупные пелагические рыбы являются океаническими кочевыми видами, совершающими длительные миграции в море. Питаются мелкой пелагической кормовой рыбой, а также пелагической рыбой среднего размера. Иногда они следуют за своей стайной добычей, и многие виды сами образуют стаи.
Примерами более крупных пелагических рыб являются тунец , марлин , королевская макрель , акулы и большие скаты .
Тунец, в частности, имеет большое значение для коммерческого рыболовства. Хотя тунцы мигрируют через океаны, пытаться найти их там – не совсем обычный подход. Тунцы, как правило, собираются в районах, где много пищи, вдоль границ течений, вокруг островов, вблизи подводных гор и в некоторых районах апвеллинга вдоль континентальных склонов. Тунца ловят несколькими методами: кошельковые неводы обволакивают весь надводный косяк специальными сетями, суда с удочками и лесками , в которых в качестве наживки используются удочки с наживкой из других более мелких пелагических рыб , а также устанавливаются плоты, называемые устройствами для сбора рыбы , поскольку тунец также как и некоторые другие пелагические рыбы, они имеют тенденцию собираться под плавучими объектами. [1]
Другие крупные пелагические рыбы являются ведущими промысловыми рыбами , особенно марлин и рыба-меч .




Апвеллинг происходит как вдоль береговой линии, так и в середине океана, когда столкновение глубинных океанских течений выносит на поверхность холодную воду, богатую питательными веществами. Эти апвеллинги способствуют цветению фитопланктона, который, в свою очередь, производит зоопланктон и поддерживает многие основные рыболовные отрасли мира. Если апвеллинг прекратится, рыболовство в этом районе прекратится. [14]
В 1960-е годы промысел перуанского анчоуса был крупнейшим промыслом в мире. Популяция анчоусов значительно сократилась во время явления Эль-Ниньо в 1972 году , когда теплая вода перетекла через холодное течение Гумбольдта в рамках 50-летнего цикла, понизив глубину термоклина . Апвеллинг прекратился, и производство фитопланктона резко упало, как и популяция анчоуса, а миллионы морских птиц , зависящих от анчоуса, погибли. [75] С середины 1980-х годов апвеллинг возобновился, и уровни вылова перуанского анчоуса вернулись к уровням 1960-х годов.
У берегов Японии столкновение течения Оясио с течением Куросио приводит к возникновению апвеллингов, богатых питательными веществами. Циклические изменения этих течений привели к сокращению популяций сардины sardinops melanosticta . Уловы рыболовства упали с 5 миллионов тонн в 1988 году до 280 тысяч тонн в 1998 году. Как следствие, тихоокеанский голубой тунец перестал завозиться в регион в поисках корма. [76] [77]
Океанские течения могут влиять на распределение рыбы, как концентрируя, так и рассеивая ее. Соседние океанские течения могут определять четкие, хотя и смещающиеся, границы. Эти границы могут быть даже видимыми, но обычно их наличие отмечается быстрыми изменениями солености, температуры и мутности. [14]
Например, в азиатской северной части Тихого океана альбакоры обитают между двумя современными системами. Северная граница определяется холодным Северо-Тихоокеанским течением , а южная граница определяется Северным экваториальным течением . Ситуация усложняется тем, что их распределение в пределах территории, определяемой двумя текущими системами, дополнительно модифицируется другим течением, течением Куросио , потоки которого колеблются в зависимости от сезона. [78]
Эпипелагические рыбы часто нерестятся в местах, где икра и личинки сносятся вниз по течению в подходящие районы нагула и, в конечном итоге, попадают в районы нагула взрослых особей. [14]
Острова и банки могут взаимодействовать с течениями и апвеллингами таким образом, что возникают области с высокой продуктивностью океана. Большие водовороты могут образовываться вниз по течению или с подветренной стороны от островов, концентрируя планктон. [79] Банки и рифы могут перехватывать глубокие течения, идущие вверх. [14]

Эпипелагические рыбы обычно перемещаются на большие расстояния между районами нагула и нереста или в ответ на изменения в океане. Крупные океанские хищники, такие как лосось и тунец, могут мигрировать на тысячи километров, пересекая океаны. [81]
В исследовании 2001 года перемещения атлантического голубого тунца из района Северной Каролины изучались с помощью специальных всплывающих тегов. Прикрепленные к тунцу, эти метки отслеживали перемещения тунца около года, затем отделились и всплыли на поверхность, где передали информацию на спутник. Исследование показало, что у тунца было четыре различных модели миграции. Одна группа на год ограничилась западной Атлантикой. Другая группа также осталась преимущественно в западной Атлантике, но на нерест мигрировала в Мексиканский залив. Третья группа пересекла Атлантический океан и обратно. Четвертая группа переправилась в восточную Атлантику, а затем направилась на нерест в Средиземное море. Исследование показывает, что, хотя существует некоторая дифференциация по нерестилищам, по существу существует только одна популяция атлантического голубого тунца, смешанные группы, которые между собой используют всю северную часть Атлантического океана, Мексиканский залив и Средиземное море. [82]
Термин «далеко мигрирующие виды» (HMS) – это юридический термин, берущий свое начало в статье 64 Конвенции Организации Объединенных Наций по морскому праву (UNCLOS). [83]
К далеко мигрирующим видам относятся: тунец и тунцеподобные виды ( альбакор , атлантический синий плавник, большеглазый тунец , скипджек , желтоперый , черноперый , маленький тунец , тихоокеанский голубой плавник , южный голубой плавник и пуля ), помфрет , марлин , парусник , рыба -меч , сайра и океанские виды . акулы , а также млекопитающие, такие как дельфины и другие китообразные .
По сути, далеко мигрирующие виды совпадают с более крупными из «крупных пелагических рыб», обсуждавшихся в предыдущем разделе, если добавить китообразных и исключить некоторые коммерчески незначимые рыбы, такие как рыба-луна . Это виды с высоким трофическим уровнем , которые совершают миграции на значительные, но переменные расстояния через океаны для питания, часто кормовой рыбой, или размножения, а также имеют широкое географическое распространение. Таким образом, эти виды встречаются как внутри 200-мильных (370-километровых) исключительных экономических зон , так и в открытом море за пределами этих зон. Это пелагические виды, что означает, что они в основном живут в открытом океане и не живут вблизи морского дна, хотя могут проводить часть своего жизненного цикла в прибрежных водах . [84]
По данным Продовольственной и сельскохозяйственной организации (ФАО), мировой улов в 2005 году составил 93,2 миллиона тонн , выловленных в результате коммерческого рыболовства в диких промыслах . [85] Из этого количества около 45% составляли пелагические рыбы. В следующей таблице показано мировое производство улова в тоннах . [86]
В 2009 году Международный союз охраны природы (МСОП) составил первый красный список океанических акул и скатов, находящихся под угрозой исчезновения. Они утверждают, что около трети акул и скатов открытого океана находятся под угрозой исчезновения . [87] В списке 64 вида океанических акул и скатов, включая молотоголовых , гигантских дьявольских скатов и морских свиней . [88]
Океанические акулы случайно вылавливаются при промысле меч-рыбы и тунца в открытом море . В прошлом существовало мало рынков сбыта акул, которые считались бесполезным приловом . Теперь акулы все чаще становятся объектом поставок на развивающиеся азиатские рынки, особенно акульих плавников , которые используются в супе из акульих плавников . [88]
По оценкам, популяция акул в северо-западной части Атлантического океана сократилась на 50% с начала 1970-х годов. Океанические акулы уязвимы, потому что они не производят много детенышей, а на созревание детенышей могут уйти десятилетия. [88]

В некоторых частях мира численность зубчатой акулы-молота сократилась на 99% с конца 1970-х годов. Его статус в Красном списке означает, что он находится под угрозой исчезновения во всем мире, то есть находится на грани исчезновения. [88]
Примечания
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )Библиография




![Длинноносая ланцетная рыба. Ланцетные рыбы — хищники, устраивающие засады и часто посещающие мезопелагические районы. Это одни из крупнейших мезопелагических рыб (до 2 метров).[37]](http://upload.wikimedia.org/wikipedia/commons/thumb/0/0d/Longnoselancetfish.jpg/1280px-Longnoselancetfish.jpg)
.jpg/1280px-Lestidiops_affinis_(1).jpg)

![Черный глотатель с растяжимым желудком примечателен способностью заглатывать целиком костистых рыб, в десять раз превышающих его массу.[50][51]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1b/Chiasmodon_niger.jpg/1280px-Chiasmodon_niger.jpg)












