Tyrannosauridae (или тираннозавриды , что означает « ящеры -тираны ») — семейство целурозавровых тероподовых динозавров , включающее два подсемейства, включающих до пятнадцати родов , включая одноименный Tyrannosaurus . Точное число родов является спорным, некоторые эксперты признают всего три. Все эти животные жили ближе к концу мелового периода , и их ископаемые останки были найдены только в Северной Америке и Азии .
Хотя тираннозавриды произошли от более мелких предков , они почти всегда были крупнейшими хищниками в своих экосистемах , что ставило их на вершину пищевой цепи . Самым крупным видом был Tyrannosaurus rex , самый массивный известный наземный хищник, который достигал более 13 метров (43 фута) в длину [2] и, согласно большинству современных оценок, весил до 8,87 метрических тонн (9,78 коротких тонн). [3] [4] Тираннозавриды были двуногими плотоядными животными с массивными черепами, заполненными крупными зубами. Несмотря на свои большие размеры, их ноги были длинными и пропорциональными для быстрого передвижения. Напротив, их руки были очень маленькими, с двумя функциональными пальцами .
В отличие от большинства других групп динозавров, для большинства известных тираннозавридов были обнаружены очень полные останки. Это позволило провести ряд исследований их биологии . Научные исследования были сосредоточены на их онтогенезе , биомеханике и экологии , среди прочих предметов.

Первые останки тираннозавридов были обнаружены во время экспедиций, возглавляемых Геологической службой Канады , которые обнаружили многочисленные разбросанные зубы. Эти отличительные зубы динозавров были названы Deinodon («ужасный зуб») Джозефом Лейди в 1856 году. Первые хорошие образцы тираннозаврида были найдены в формации Horseshoe Canyon в Альберте и состояли из почти полных черепов с частичными скелетами. Эти останки были впервые изучены Эдвардом Дринкером Коупом в 1876 году, который считал их видом восточного тираннозавроида Dryptosaurus . В 1905 году Генри Фэрфилд Осборн признал, что останки из Альберты значительно отличаются от Dryptosaurus , и придумал для них новое название: Albertosaurus sarcophagus («плотоядный ящер Альберты»). [5] Коуп описал больше материала тираннозавра в 1892 году в виде изолированных позвонков и дал этому животному название Manospondylus gigas . Это открытие в основном игнорировалось более столетия и вызвало споры в начале 2000-х годов, когда было обнаружено, что этот материал на самом деле принадлежал Tyrannosaurus rex и имел приоритет над названием . [6]

В своей статье 1905 года, называющей Albertosaurus , Осборн описал два дополнительных образца тираннозавра, которые были собраны в Монтане и Вайоминге во время экспедиции Американского музея естественной истории 1902 года под руководством Барнума Брауна . Первоначально Осборн считал их отдельными видами. Первого он назвал Dynamosaurus imperiosus («ящерица императорской власти»), а второго — Tyrannosaurus rex («ящер-король-тиран»). Год спустя Осборн признал, что эти два образца на самом деле принадлежат к одному и тому же виду. Несмотря на то, что Dynamosaurus был найден первым, название Tyrannosaurus появилось на одну страницу раньше в его оригинальной статье, описывающей оба образца. Поэтому, согласно Международному кодексу зоологической номенклатуры (ICZN), было использовано название Tyrannosaurus . [7]
Барнум Браун продолжил собирать еще несколько образцов тираннозавридов из Альберты, включая первый, сохранивший укороченные, двупалые передние конечности, характерные для этой группы (которую Лоуренс Лэмб назвал Gorgosaurus libratus , «сбалансированный свирепый ящер», в 1914 году). Вторая значительная находка, приписываемая горгозавру , была сделана в 1942 году в виде хорошо сохранившегося, хотя и необычно маленького, полного черепа. Образец ждал до окончания Второй мировой войны , пока его не изучил Чарльз У. Гилмор , который назвал его Gorgosaurus lancesnis . [5] Этот череп был повторно изучен Робертом Т. Баккером , Филом Карри и Майклом Уильямсом в 1988 году и отнесен к новому роду Nanotyrannus . [8] Также в 1946 году палеонтологи из Советского Союза начали экспедиции в Монголию и обнаружили первые останки тираннозавра из Азии. Евгений Малеев описал новые монгольские виды тираннозавра и горгозавра в 1955 году, и один новый род: тарбозавр («ужасный ящер»). Последующие исследования, однако, показали, что все виды тираннозавров Малеева на самом деле были одним видом тарбозавра на разных стадиях роста. Второй вид монгольского тираннозаврида был найден позже, описан Сергеем Курзановым в 1976 году и получил название Alioramus remotus («отдаленная другая ветвь»), хотя его статус как настоящего тираннозаврида, а не более примитивного тираннозавра до сих пор остается спорным. [9] [5]

Все тираннозавриды были крупными животными, и все виды могли весить не менее 1 тонны. [10] Был обнаружен единственный экземпляр алиорама , длина которого оценивалась от 5 до 6 метров (от 16 до 20 футов), [9] хотя некоторые эксперты считают его молодой особью. [10] [11] Альбертозавр , горгозавр и дасплетозавр имели длину от 8 до 9 метров (от 26 до 30 футов), [12] в то время как тарбозавр достигал длины 11 метров (36 футов) от морды до хвоста. [13] Массивный тираннозавр достигал 13 метров (43 фута) в самом крупном экземпляре, RSM P2523.8 . [14]
Анатомия черепа тираннозавридов хорошо изучена, поскольку полные черепа известны для всех родов, кроме Alioramus , который известен только по частичным останкам черепа. [15] У тираннозавра , тарбозавра и дасплетозавра были черепа, длина которых превышала 1 м (3,3 фута). [12] У взрослых тираннозавридов были высокие, массивные черепа со многими сросшимися и укрепленными для прочности костями. В то же время полые камеры во многих костях черепа и большие отверстия ( фенестры ) между этими костями помогали уменьшить вес черепа. Многие черты черепов тираннозавридов были также обнаружены у их непосредственных предков, включая высокие предчелюстные кости и сросшиеся носовые кости . [10]
Черепа тираннозавридов имели много уникальных характеристик, включая сросшиеся теменные кости с выступающим сагиттальным гребнем , который шел продольно вдоль сагиттального шва и разделял два надвисочных окна на крыше черепа. За этими окнами тираннозавриды имели характерно высокий затылочный гребень, который также возникал из теменных костей, но проходил вдоль поперечной плоскости, а не продольно. Затылочный гребень был особенно хорошо развит у тираннозавра , тарбозавра и алиорама . Альбертозавр , дасплетозавр и горгозавр имели высокие гребни перед глазами на слезных костях , в то время как тарбозавр и тираннозавр имели чрезвычайно утолщенные заглазничные кости, образующие гребни в форме полумесяца позади глаз. У алиорама был ряд из шести костных гребней на вершине морды, возникающих из носовых костей; Более низкие гребни были зарегистрированы у некоторых образцов Daspletosaurus и Tarbosaurus , а также у более базального тираннозавроида Appalachiosaurus . [11] [16] Морда и другие части черепа также имели многочисленные отверстия . Согласно исследованию 2017 года, в котором описывался D. horneri , чешуйчатый покров, а также тактильная чувствительность коррелировали с несколькими рядами нейрососудистых отверстий, наблюдаемых у крокодилов и тираннозавридов. [17]
Тираннозавриды, как и их предки-тираннозавроиды, были гетеродонтами , с предчелюстными зубами D -образной формы в поперечном сечении и меньшими, чем у остальных. В отличие от более ранних тираннозавроидов и большинства других теропод, верхнечелюстные и нижнечелюстные зубы зрелых тираннозавридов не лезвиевидные, а чрезвычайно утолщенные и часто круглые в поперечном сечении, причем некоторые виды имеют уменьшенные зазубрины. [10] Количество зубов, как правило, одинаково в пределах вида, и более крупные виды, как правило, имеют меньшее количество зубов, чем более мелкие. Например, у алиорама было от 76 до 78 зубов в челюстях, в то время как у тираннозавра их было от 54 до 60. [18]
Уильям Эблер заметил в 2001 году, что зубные зазубрины альбертозавра напоминают трещину в зубе, заканчивающуюся круглой пустотой, называемой ампулой. [19] Зубы тираннозавров использовались в качестве фиксаторов для отрывания мяса от тела, поэтому, когда тираннозавр тянул кусок мяса назад, напряжение могло привести к распространению чисто трещиноподобной зазубрины по зубу. [19] Однако наличие ампулы распределило бы эти силы по большей площади поверхности и уменьшило бы риск повреждения зуба при нагрузке. [19] Наличие надрезов, заканчивающихся пустотами, имеет параллели в человеческой инженерии. Производители гитар используют надрезы, заканчивающиеся пустотами, чтобы, как описывает Эблер, «придать чередующиеся области гибкости и жесткости» древесине, с которой они работают. [19] Использование сверла для создания своего рода «ампулы» и предотвращения распространения трещин по материалу также используется для защиты поверхностей самолетов . [19] Эблер продемонстрировал, что плексигласовый брусок с надрезами, называемыми «прорезями», и просверленными отверстиями был более чем на 25% прочнее, чем брусок с только регулярно расположенными надрезами. [19] В отличие от тираннозавров и других тероподов, древние хищники, такие как фитозавры и диметродон, не имели никаких приспособлений, которые бы предотвращали распространение трещинообразных зазубрин их зубов под воздействием сил, возникающих при кормлении. [19]

Череп располагался на конце толстой S -образной шеи, а длинный, тяжелый хвост служил противовесом, уравновешивая голову и туловище, с центром массы над бедрами. Тираннозавриды известны своими пропорционально очень маленькими двупалыми передними конечностями, хотя иногда встречаются остатки рудиментарного третьего пальца. [10] [20] У тарбозавра были самые короткие передние конечности по сравнению с размером тела, в то время как у дасплетозавра были самые длинные.

Тираннозавриды ходили исключительно на своих задних конечностях, поэтому их кости ног были массивными. В отличие от передних конечностей, задние конечности были длиннее по сравнению с размером тела, чем почти у любых других тероподов. Молодь и даже некоторые более мелкие взрослые особи, как и более базальные тираннозавроиды, имели более длинные большеберцовые кости , чем бедренные , что характерно для бегающих (быстро бегающих) динозавров, таких как орнитомимиды . Более крупные взрослые особи имели пропорции ног, характерные для более медлительных животных, но не в такой степени, как у других крупных тероподов, таких как абелизавриды или карнозавры . Третьи плюсневые кости тираннозавридов были зажаты между второй и четвертой плюсневыми костями, образуя структуру, известную как арктометатарзус . [10] Тираннозавриды также имели большие сохранившиеся прикрепления мышц ног и низкую инерцию вращения относительно массы тела, что указывает на то, что они могли поворачиваться быстрее, чем другие крупные тероподы. [21] [22]
Неясно, когда арктометатарзус впервые появился; он не присутствовал у самых ранних тираннозавроидов, таких как Dilong , [23], но был обнаружен у более позднего Appalachiosaurus . [16] Эта структура также характерна для троодонтид , орнитомимид и ценагнатид , [24] но ее отсутствие у самых ранних тираннозавроидов указывает на то, что она была приобретена в результате конвергентной эволюции . [23]
Сравнительный анализ зубов показывает, что тираннозавриды, как и другие крупные тероподы, имели губы, которые защищали их зубы от внешних повреждений. Эта анатомическая особенность придавала этим животным больше визуального сходства с лепидозаврами, чем с близкородственными крокодилами. [25]
Название Deinodontidae было придумано Эдвардом Дринкером Коупом в 1866 году для этого семейства [26] и продолжало использоваться вместо нового названия Tyrannosauridae вплоть до 1960-х годов. [27] Типовым родом Deinodontidae является Deinodon , который был назван в честь изолированных зубов из Монтаны . [28] Однако в обзоре североамериканских тираннозавров 1970 года Дейл Рассел пришел к выводу, что Deinodon не является действительным таксоном, и использовал название Tyrannosauridae вместо Deinodontidae, заявив, что это соответствует правилам МКЗН . [12] Поэтому современные эксперты предпочитают Tyrannosauridae. [5]
Тираннозавр был назван Генри Фэрфилдом Осборном в 1905 году вместе с семейством Tyrannosauridae. [29] Название происходит от древнегреческих слов τυραννος ( tyrannos ) ('тиран') и σαυρος ( sauros ) ('ящерица'). Очень распространенный суффикс -idae обычно добавляется к зоологическим семейным именам и происходит от греческого суффикса -ιδαι - idai , который указывает на множественное число существительного. [30]

Tyrannosauridae — семейство в ранговой таксономии Линнея , входящее в надсемейство Tyrannosauroidea и подотряд Theropoda .
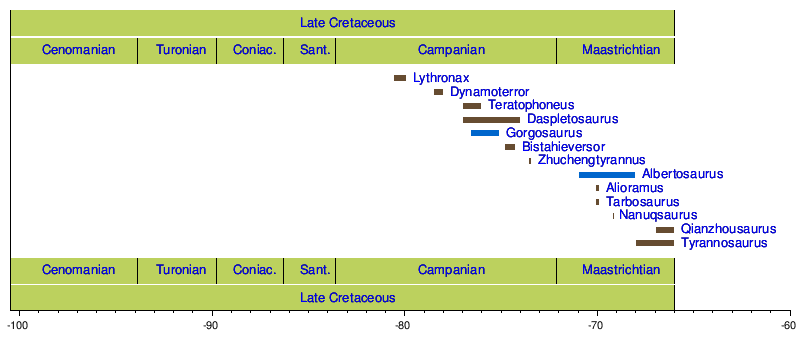
Tyrannosauridae бесспорно делится на два подсемейства. Albertosaurinae включает североамериканские роды Albertosaurus и Gorgosaurus , в то время как Tyrannosaurinae включает Daspletosaurus , Teratophoneus , Bistahieversor , Tarbosaurus , Nanuqsaurus , Zhuchengtyrannus и самого Tyrannosaurus . [31] Некоторые авторы включают вид Gorgosaurus libratus в род Albertosaurus , а Tarbosaurus bataar в род Tyrannosaurus , [16] [5] [32] в то время как другие предпочитают сохранять Gorgosaurus и Tarbosaurus как отдельные роды. [10] [11] Альбертозаврины характеризуются более стройным телосложением, более низким черепом и пропорционально более длинными большеберцовыми костями , чем тираннозаврины. [10] У тираннозавров сагиттальный гребень на теменных костях продолжается вперед на лобные кости. [11] В 2014 году Люй Цзюньчан и др. описали Alioramini как трибу в пределах Tyrannosauridae, содержащую роды Alioramus и Qianzhousaurus . Их филогенетический анализ показал, что триба располагалась у основания Tyrannosaurinae. [33] [34] Некоторые авторы, такие как Джордж Ольшевский и Трейси Форд, создали другие подразделения или трибы для различных комбинаций тираннозавридов в пределах подсемейств. [35] [36] Однако они не были филогенетически определены и обычно состояли из родов, которые теперь считаются синонимами других родов или видов. [18]
Дополнительные подсемейства были названы для более фрагментарных родов, включая Aublysodontinae и Deinodontinae . Однако роды Aublysodon и Deinodon обычно считаются nomina dubia , поэтому они и их одноименные подсемейства обычно исключаются из таксономий тираннозавридов. Дополнительный тираннозавр, Raptorex , изначально был описан как более примитивный тираннозавроид, но, вероятно, представляет собой молодого тираннозаврина, похожего на Tarbosaurus . Однако, поскольку он известен только по молодому образцу, в настоящее время он также считается nomen dubium . [37] Однако Томас Карр поддерживает его обоснованность и считает, что он похож на тираннозавринов. [38]
С появлением филогенетической таксономии в палеонтологии позвоночных, Tyrannosauridae получили несколько явных определений. Оригинал был создан Полом Серено в 1998 году и включал всех тираннозавроидов, более близких к Tyrannosaurus, чем к Alectrosaurus , Aublysodon или Nanotyrannus . [39] Однако Nanotyrannus часто считается молодой особью Tyrannosaurus rex , в то время как Aublysodon обычно рассматривается как nomen dubium, непригодный для использования в определении клады . [ 10] Определения с тех пор основывались на более устоявшихся родах.
В 2001 году Томас Р. Хольц -младший опубликовал кладистический анализ Tyrannosauridae. [40] Он пришел к выводу, что существует два подсемейства : более примитивное Aublysodontinae, характеризующееся не зазубренными предчелюстными зубами ; и Tyrannosaurinae . [40] Aublysodontinae включали Aublysodon , « Kirtland Aublysodon » и Alectrosaurus . [40] Хольц также обнаружил, что Siamotyrannus демонстрировал некоторые синапоморфии tyrannosauridae , но находился «вне [ семейства ] как такового». [40]
Позже в той же статье он предложил определить Tyrannosauridae как «всех потомков самого недавнего общего предка Tyrannosaurus и Aublysodon ». [40] Он также раскритиковал определения, ранее предложенные другими исследователями, например, одно, предложенное Полом Серено , что Tyrannosauridae были «всеми таксонами, более близкими к «Tyrannosaurus», чем к Alectrosaurus , Aublysodon и Nanotyrannus ». [40] Хольц заметил, что, поскольку Nanotyrannus , вероятно, был неправильно идентифицированной молодой особью T. rex , предложенное Серено определение будет включать семейство Tyrannosauridae в качестве подтаксона рода Tyrannosaurus . [40] Кроме того, предложенное им определение подсемейства Tyrannosaurinae также будет ограничено Tyrannosaurus . [40]
Попытка Кристофера Брошу 2003 года включить в определение Albertosaurus , Alectrosaurus , Alioramus , Daspletosaurus , Gorgosaurus , Tarbosaurus и Tyrannosaurus . [41] Хольц переопределил кладу в 2004 году, чтобы использовать все вышеперечисленное в качестве спецификаторов, за исключением Alioramus и Alectrosaurus , которые его анализ не мог разместить с уверенностью. Однако в той же статье Хольц также предоставил совершенно другое определение, включив всех тероподов, более тесно связанных с Tyrannosaurus, чем с Eotyrannus . [10] Самое последнее определение - это определение Серено в 2005 году, который определил Tyrannosauridae как наименее инклюзивную кладу, содержащую Albertosaurus , Gorgosaurus и Tyrannosaurus . [42]
Кладистический анализ филогении тираннозавров часто обнаруживает, что Tarbosaurus и Tyrannosaurus являются сестринскими таксонами , причем Daspletosaurus более базальный, чем любой из них. Тесная связь между Tarbosaurus и Tyrannosaurus подтверждается многочисленными особенностями черепа, включая рисунок швов между определенными костями, наличие серповидного гребня на заглазничной кости позади каждого глаза и очень глубокую верхнюю челюсть с заметным изгибом вниз на нижнем крае, среди прочего. [10] [16] Альтернативная гипотеза была представлена в исследовании 2003 года Филом Карри и коллегами, которые нашли слабую поддержку Daspletosaurus как базального члена клады, также включающей Tarbosaurus и Alioramus , оба из Азии, на основании отсутствия костного выступа, соединяющего носовую и слезную кости. [18] В этом исследовании было обнаружено, что алиорамус является ближайшим родственником тарбозавра , основываясь на схожей схеме распределения напряжений в черепе.
Связанное исследование также отметило запирающий механизм в нижней челюсти, общий для двух родов. [43] В отдельной статье Карри отметил возможность того, что Алиорамус может представлять собой молодую особь тарбозавра , но заявил, что гораздо большее количество зубов и более выступающие носовые гребни у Алиорама предполагают, что это отдельный род. Аналогичным образом Карри использует большое количество зубов у Нанотирануса, чтобы предположить, что это может быть отдельный род, [11] а не молодая особь тираннозавра , как считают большинство других экспертов. [10] [44] Однако открытие и описание Цяньчжоузавра показывает, что Алиорамус не является близким родственником тарбозавра , а принадлежит к недавно описанному трибе тираннозавридов; Алиорамини. Qianzhousaurus далее показывает, что подобные длинномордые тираннозавриды были широко распространены по всей Азии и могли бы жить в одной и той же среде, избегая при этом конкуренции с более крупными и крепкими тираннозаврами, охотясь на другую добычу. [45]
Палеонтолог Грегори Эриксон и его коллеги изучали рост и историю жизни тираннозавридов. Анализ гистологии костей может определить возраст образца, когда он умер. Темпы роста можно изучить, если нанести на график возраст различных особей против их размера. Эриксон показал, что после долгого пребывания в качестве ювенильных особей тираннозавры претерпевали огромные скачки роста в течение примерно четырех лет в середине своей жизни. После того, как фаза быстрого роста закончилась с наступлением половой зрелости , рост значительно замедлился у взрослых животных. Кривая роста тираннозавридов имеет S-образную форму, с максимальной скоростью роста особей около 14 лет. [49]

Самая маленькая известная особь Tyrannosaurus rex ( LACM 28471, «иорданский теропод»), по оценкам, весила всего 29,9 кг (66 фунтов) в возрасте всего 2 лет, в то время как самая большая, такая как FMNH PR2081 (« Сью »), скорее всего, весила около 5654 кг (12465 фунтов) в возрасте 28 лет, что, возможно, близко к максимальному для этого вида. [49] Молодые особи T. rex оставались весом менее 1800 кг (4000 фунтов) примерно до 14 лет, когда размер тела начал резко увеличиваться. Во время этой фазы быстрого роста молодой T. rex набирал в среднем 600 кг (1300 фунтов) в год в течение следующих четырех лет. Это замедлилось после 16 лет, и в возрасте 18 лет кривая снова вышла на плато, что указывает на резкое замедление роста. [50] Например, всего 600 кг (1300 фунтов) отделяли 28-летнюю «Сью» от 22-летнего канадского экземпляра ( RTMP 81.12.1). [49] Это внезапное изменение в скорости роста может указывать на физическую зрелость, гипотеза, которая подтверждается обнаружением медуллярной ткани в бедренной кости 18-летнего тираннозавра из Монтаны ( MOR 1125, также известного как «B-rex»). [51] Медуллярная ткань обнаруживается только у самок птиц во время овуляции, что указывает на то, что «B-rex» находился в репродуктивном возрасте. [52]
Другие тираннозавриды демонстрируют чрезвычайно похожие кривые роста, хотя с более низкими темпами роста, соответствующими их меньшим размерам во взрослом состоянии. [53] По сравнению с альбертозаврами, дасплетозавр показал более высокую скорость роста в период быстрого роста из-за своего более высокого веса во взрослом состоянии. Максимальная скорость роста дасплетозавра составляла 180 килограммов (400 фунтов) в год, исходя из оценки массы взрослых особей в 1800 кг (4000 фунтов). Другие авторы предполагали более высокий вес взрослых особей для дасплетозавра ; это изменило бы величину скорости роста, но не общую картину. [49] Самый молодой известный альбертозавр — двухлетний, обнаруженный в костном слое острова Драй-Айленд, который весил около 50 кг (110 фунтов) и имел длину чуть более 2 метров (6,6 фута). 10-метровый (33 фута) экземпляр из того же карьера является старейшим и крупнейшим из известных, ему 28 лет. По оценкам, самая быстрая скорость роста наблюдается в возрасте 12–16 лет, достигая 122 кг (269 фунтов) в год, исходя из веса взрослой особи 1300 кг (2900 фунтов), что составляет примерно пятую часть скорости для T.-rex . Для Gorgosaurus расчетная максимальная скорость роста составляет около 110 килограммов (240 фунтов) во время фазы быстрого роста, что сопоставимо с таковой для Albertosaurus . [49]
Открытие эмбрионального тираннозавра пока неизвестного рода предполагает, что тираннозавриды развили свои отличительные черты скелета во время развития в яйце. Кроме того, размер образца, зубная кость длиной 1,1 дюйма (2,8 см) из нижней челюсти, найденная в формации Ту-Медисин в Монтане в 1983 году, и коготь стопы, найденный в формации Хорсшу-Каньон в 2018 году и описанный в 2020 году, предполагает, что новорожденные тираннозавриды рождались с черепами размером с мышь или грызунов аналогичного размера и могли быть примерно размером с небольшую собаку при рождении. Считается, что образец челюсти принадлежал животному длиной около 2,5 футов (0,76 м), в то время как коготь, как полагают, принадлежит образцу длиной около 3 футов (0,91 м). Хотя яичная скорлупа не была найдена ни у одного из образцов, место, где были обнаружены эти новорожденные тираннозавриды, позволяет предположить, что эти животные использовали те же места гнездования, что и другие виды, с которыми они жили и на которых охотились. [54] Отсутствие яичной скорлупы, связанной с этими образцами, также открыло возможность для предположений о том, что тираннозавриды откладывали яйца с мягкой скорлупой, как, как полагают, делали роды Mussaurus и Protoceratops . [55]
Ископаемые следы из формации Вапити предполагают, что по мере роста тираннозавридов, ступни становились шире с более толстыми пальцами, чтобы выдерживать их вес. Более широкие ступни предполагают, что взрослые тираннозавриды были медленнее, чем их потомки. [56] [57]

Окончание фазы быстрого роста предполагает начало половой зрелости у альбертозавра , хотя рост продолжался более медленными темпами на протяжении всей жизни животных. [49] [53] Половое созревание во время активного роста, по-видимому, является общей чертой среди мелких [58] и крупных [59] динозавров, а также среди крупных млекопитающих, таких как люди и слоны . [59] Эта модель относительно раннего полового созревания разительно отличается от модели у птиц, которые откладывают свое половое созревание до тех пор, пока они не закончат расти. [59] [60]
Составив таблицу с количеством особей каждой возрастной группы, Эриксон и его коллеги смогли сделать выводы о жизненном цикле популяций тираннозавридов. Их анализ показал, что, хотя молодые особи были редки в палеонтологической летописи, полувзрослые особи в фазе быстрого роста и взрослые особи были гораздо более распространены. Более половины известных особей T. rex , по-видимому, умерли в течение шести лет после достижения половой зрелости, что также наблюдается у других тираннозавров и у некоторых крупных долгоживущих птиц и млекопитающих сегодня. Для этих видов характерны высокие показатели детской смертности, за которыми следует относительно низкая смертность среди молодых особей. Смертность снова увеличивается после половой зрелости, отчасти из-за стрессов, связанных с размножением. Хотя это могло быть связано с предвзятостью сохранения или сбора , Эриксон предположил, что разница была вызвана низкой смертностью среди молодых особей после определенного размера, что также наблюдается у некоторых современных крупных млекопитающих, таких как слоны . Такая низкая смертность могла быть результатом отсутствия хищников, поскольку тираннозавры превосходили всех современных хищников по размеру к двум годам. Палеонтологи не нашли достаточно останков дасплетозавра для аналогичного анализа, но Эриксон отмечает, что та же общая тенденция, по-видимому, применима. [53]
Тираннозаврид проводил до половины своей жизни в ювенильной фазе, прежде чем раздуться до почти максимального размера всего за несколько лет. [49] Это, наряду с полным отсутствием хищников, промежуточных по размеру между огромными взрослыми тираннозавридами и другими мелкими тероподами, предполагает, что эти ниши могли быть заполнены молодыми тираннозавридами. Это видно на примере современных комодских варанов , где детеныши начинают как насекомоядные, живущие на деревьях , и медленно созревают в огромных хищников, способных уничтожать крупных позвоночных. [10] Например, альбертозавры были обнаружены в скоплениях, которые, по мнению некоторых, представляют собой стаи разновозрастных особей . [61] [62]
Способности передвижения лучше всего изучены для тираннозавра , и есть два основных вопроса, касающихся этого: насколько хорошо он мог поворачиваться; и какова была его максимальная скорость по прямой. Тираннозавр , возможно, медленно поворачивался, возможно, тратя одну-две секунды, чтобы повернуть всего на 45° — величина, которую люди, будучи вертикально ориентированными и бесхвостыми, могут повернуть за долю секунды. [63] Причиной трудности является инерция вращения , поскольку большая часть массы тираннозавра находилась на некотором расстоянии от его центра тяжести, как человек, несущий тяжелое бревно. [ 64]
Ученые вывели широкий диапазон оценок максимальной скорости, в основном около 11 метров в секунду (25 миль в час), но некоторые из них были всего 5–11 метров в секунду (11–25 миль в час), а некоторые — 20 метров в секунду (45 миль в час). Исследователям приходится полагаться на различные методы оценки, поскольку, хотя есть много следов очень крупных тероподов, идущих пешком, до сих пор не было найдено ни одного следа очень крупных тероподов, бегущих — и это отсутствие может указывать на то, что они не бегали. [65]
Джек Хорнер и Дон Лессем утверждали в 1993 году, что тираннозавр был медлительным и, вероятно, не мог бегать (отсутствие фазы полета в середине шага). [66] Однако Хольц (1998) пришел к выводу, что тираннозавриды и их близкие родственники были самыми быстрыми крупными тероподами. [67] Кристиансен (1998) подсчитал, что кости ног тираннозавра были не намного прочнее, чем у слонов, которые относительно ограничены в своей максимальной скорости и никогда не бегают (отсутствие фазы полета), и поэтому предположил, что максимальная скорость динозавра составляла около 11 метров в секунду (25 миль в час), что примерно соответствует скорости человека-спринтера. [68] Фарлоу и коллеги (1995) утверждали, что тираннозавр весом от 6 до 8 тонн получил бы критические или даже смертельные травмы, если бы упал во время быстрого движения, поскольку его туловище врезалось бы в землю с замедлением 6 g (в шесть раз больше ускорения свободного падения, или около 60 метров/с2 ) , а его крошечные руки не смогли бы смягчить удар. [69] [70] Однако известно , что жирафы скачут со скоростью 50 км/ч (31 миля в час), несмотря на риск сломать ногу или что-то похуже, что может быть фатальным даже в «безопасной» среде, такой как зоопарк. [71] [72] Таким образом, вполне возможно, что тираннозавр также быстро передвигался, когда это было необходимо, и должен был принимать такие риски; этот сценарий изучался и для аллозавра . [73] [74] Последние исследования передвижения тираннозавра не сужают диапазон скоростей дальше, чем от 17 до 40 км/ч (от 11 до 25 миль/ч), то есть от ходьбы или медленного бега до бега с умеренной скоростью. [65] [75] [76] Исследование компьютерной модели в 2007 году оценило скорость бега на основе данных, взятых непосредственно из ископаемых, и заявило, что максимальная скорость бега у T. rex составляла 8 метров в секунду (18 миль/ч). [77] [78] (Вероятно, молодая особь. [79] )
Исследования Эрика Снивели и др., опубликованные в 2019 году, показывают, что тираннозавриды, такие как тарбозавр и сам тираннозавр, были более маневренными, чем аллозавроиды сопоставимого размера из-за низкой инерции вращения по сравнению с их массой тела в сочетании с большими мышцами ног. В результате предполагается, что тираннозавриды были способны совершать относительно быстрые повороты и, вероятно, могли поворачивать свое тело быстрее, находясь близко к своей добыче, или что во время поворота они могли «пируэты» на одной поставленной ноге, в то время как чередующаяся нога удерживалась в подвешенном состоянии во время преследования. Результаты этого исследования потенциально могут пролить свет на то, как ловкость могла способствовать успеху эволюции тираннозавридов. [80]
Кроме того, исследование 2020 года показывает, что тираннозавриды были исключительно эффективными ходоками. Исследования Декекки и др. сравнили пропорции ног, массу тела и походку более 70 видов динозавров-теропод, включая тираннозавридов. Затем исследовательская группа применила различные методы для оценки максимальной скорости каждого динозавра при беге, а также того, сколько энергии каждый динозавр тратит при движении с более спокойной скоростью, например, при ходьбе. Среди видов меньшего и среднего размера, таких как дромеозавриды, более длинные ноги, по-видимому, являются адаптацией к более быстрому бегу, что соответствует предыдущим результатам других исследователей. Но для теропод весом более 1000 кг (2200 фунтов) максимальная скорость бега ограничена размером тела, поэтому было обнаружено, что более длинные ноги коррелируют с ходьбой с низким потреблением энергии. Результаты исследования также показали, что более мелкие тероподы развили длинные ноги для скорости как средства как для помощи в охоте, так и для спасения от более крупных хищников, в то время как более крупные хищные тероподы, которые развили длинные ноги, сделали это для снижения затрат энергии и повышения эффективности добычи пищи, поскольку они были освобождены от требований давления хищников из-за своей роли в качестве высших хищников. По сравнению с более базальными группами теропод в исследовании, тираннозавриды показали заметное увеличение эффективности добычи пищи из-за снижения затрат энергии во время охоты и сбора падали. Это, вероятно, привело к тому, что тираннозавры имели меньшую потребность в охотничьих вылазках и в результате требовали меньше пищи для поддержания себя. Кроме того, исследование, в сочетании с исследованиями, которые показывают, что тираннозавры были более проворными, чем другие крупные тероподы, указывает на то, что они были довольно хорошо адаптированы к дальнему преследованию с последующим быстрым всплеском скорости для убийства. В результате можно отметить аналогии между тираннозаврами и современными волками, подтверждаемые доказательствами того, что по крайней мере некоторые тираннозавриды, такие как альбертозавр, охотились в группах. [81] [82]

Продолжающиеся дебаты в палеонтологическом сообществе касаются степени и природы покровного покрытия тираннозавров. Длинные нитевидные структуры сохранились вместе со скелетными останками многочисленных целурозавров из раннемеловой формации Исянь и других близлежащих геологических формаций из Ляонина , Китай. [83] Эти нити обычно интерпретировались как «протоперья», гомологичные разветвленным перьям, обнаруженным у птиц и некоторых нептичьих теропод , [84] [85] хотя были предложены и другие гипотезы. [86] В 2004 году был описан скелет Дилонга , который включал первый пример «протоперьев» у тираннозавроида. Подобно пуховым перьям современных птиц, «протоперья», обнаруженные у Дилонга, были разветвленными, но не контурными , и, возможно, использовались для изоляции . [23] Открытие и описание 9-метрового (30 футов) пернатого тираннозавроида Ютираннуса в 2012 году указывает на возможность того, что крупные тираннозавриды также были оперенными во взрослом возрасте. [87]

На основе принципа филогенетического брекетинга было предсказано, что тираннозавриды также могут обладать таким оперением. Однако исследование, опубликованное в 2017 году группой исследователей в Biology Letters, описало отпечатки кожи тираннозавридов, собранные в Альберте, Монтане и Монголии, которые принадлежали пяти родам ( Tyrannosaurus , Albertosaurus , Gorgosaurus , Daspletosaurus и Tarbosaurus ). [88] Хотя отпечатки кожи небольшие, они широко разбросаны по посткраниуму, будучи коллективно расположенными на животе, грудной области, подвздошной кости, тазу, хвосте и шее. Они показывают плотный рисунок из мелких, не перекрывающихся галечных чешуек (которые соавтор Скотт Персонс сравнил с теми, которые видны на боках крокодила [89] ), и не сохраняют никаких намеков на оперение. Основная текстура состоит из крошечных «базальных чешуек» диаметром приблизительно от 1 до 2 мм, с некоторыми отпечатками, показывающими 7-миллиметровые «чешуйки черт», вкрапленные между ними. Дополнительные чешуйки можно увидеть в отпечатках ног тираннозавров. [90] Исследования показывают, что лицевой покров тираннозавров имел чешуйки на зубной и верхней челюстях, ороговевший эпидермис и подобную броне кожу на подчиненных областях. [91] [92]
Белл и др. провели реконструкцию предкового признака на основе того, что известно о распределении покровов у тираннозавроидов. Несмотря на 89% вероятность того, что тираннозавроиды изначально имели перья, они определили, что чешуйчатые тираннозавриды имеют 97% вероятность быть правдой. Данные «предоставляют убедительные доказательства полностью чешуйчатого покрытия у тираннозавра», написала команда, хотя они признали, что оперение могло все еще присутствовать на дорсальной области, где отпечатки кожи еще не были обнаружены. Белл и др. выдвигают гипотезу, что отпечатки чешуи тираннозавридов, возможно, являются сетками, которые вторично произошли от перьев, хотя для подтверждения этого необходимы доказательства. [88] Однако другие утверждают, что это из-за тафономического смещения у тираннозавридов. [93]
Пока еще не определено, почему могло произойти такое изменение покрова. Прецедент потери перьев можно увидеть в других группах динозавров, таких как орнитисхии , у которых нитевидные структуры были утеряны, а чешуя появилась снова. [94] Хотя гигантизм был предложен в качестве механизма, Фил Р. Белл, который был соавтором исследования, отметил, что пернатый ютираннус перекрывался по размеру с горгозавром и альбертозавром . «Проблема здесь в том, что у нас есть большие тираннозавры, некоторые с перьями, некоторые без них, живущие в довольно похожем климате. Так в чем же причина этой разницы? Мы действительно не знаем». [95]

Глазницы тираннозавра расположены так, что глаза смотрят вперед, что дает им бинокулярное зрение немного лучше, чем у современных ястребов . В то время как хищные тероподы в целом имели бинокулярное зрение прямо перед своим черепом, тираннозавры имели значительно большую область перекрытия. Джек Хорнер также указал, что у линии тираннозавров была история неуклонного улучшения бинокулярного зрения. Трудно понять, как естественный отбор благоприятствовал бы этой долгосрочной тенденции, если бы тираннозавры были чистыми падальщиками, которым не требовалось бы развитое восприятие глубины , которое обеспечивает стереоскопическое зрение . [96] [97] У современных животных бинокулярное зрение встречается в основном у хищников (главным исключением являются приматы , которым оно необходимо для прыжков с ветки на ветку). В отличие от тираннозавра , у тарбозавра был более узкий череп, более типичный для других тираннозавридов, у которых глаза были обращены в основном вбок. Все это говорит о том, что тарбозавр больше полагался на обоняние и слух, чем на зрение. [98] У образцов горгозавра глазница была круглой, а не овальной или в форме замочной скважины, как у других родов тираннозавров. [11] У дасплетозавра это был высокий овал, где-то между круглой формой, наблюдаемой у горгозавра , и формой «замочной скважины» у тираннозавра . [10] [11] [44]
На основе сравнения текстуры костей дасплетозавра с современными крокодилами , подробное исследование, проведенное в 2017 году Томасом Д. Карром и соавторами, показало, что у тираннозавров были большие плоские чешуйки на мордах . [99] [100] В центре этих чешуек находились небольшие ороговевшие участки. У крокодилов такие участки покрывают пучки сенсорных нейронов , которые могут обнаруживать механические, термические и химические раздражители . [101] [102] Они предположили, что у тираннозавров, вероятно, также были пучки сенсорных нейронов под лицевыми чешуйками , и они могли использовать их для идентификации объектов, измерения температуры своих гнезд и осторожного подбора яиц и детенышей . [99]

Костные гребни обнаружены на черепах многих теропод, включая многих тираннозавридов. Alioramus , возможный тираннозавр из Монголии, имеет один ряд из пяти выступающих костных бугорков на носовых костях; аналогичный ряд гораздо более низких бугорков присутствует на черепе Appalachiosaurus , а также некоторых экземпляров Daspletosaurus , Albertosaurus и Tarbosaurus . [16] У Albertosaurus , Gorgosaurus и Daspletosaurus есть выступающий рог перед каждым глазом на слезной кости. Слезный рог отсутствует у Tarbosaurus и Tyrannosaurus , у которых вместо этого есть гребень в форме полумесяца позади каждого глаза на заглазничной кости . Эти гребни на голове могли использоваться для демонстрации , возможно, для распознавания вида или ухаживания . [10]
Tyrannosaurus , как и большинство динозавров, долгое время считался имеющим эктотермический («холоднокровный») рептильный метаболизм , но был оспорен такими учеными, как Роберт Т. Баккер и Джон Остром в первые годы « Ренессанса динозавров », начавшегося в конце 1960-х годов. [103] [104] Сам Tyrannosaurus rex , как утверждалось , был эндотермическим («теплокровным»), что подразумевает очень активный образ жизни. [105] С тех пор несколько палеонтологов пытались определить способность Tyrannosaurus регулировать температуру своего тела . Гистологические свидетельства высоких темпов роста у молодых T. rex , сопоставимых с темпами роста у млекопитающих и птиц, могут подтверждать гипотезу о высоком метаболизме. Кривые роста указывают на то, что, как и у млекопитающих и птиц, рост T. rex был ограничен в основном неполовозрелыми животными, а не неопределенным ростом, наблюдаемым у большинства других позвоночных . [50] Было указано, что разница температур могла быть не более 4–5 °C (7–9 °F) между позвонками туловища и большеберцовой костью голени. Этот небольшой диапазон температур между ядром тела и конечностями был заявлен палеонтологом Ризом Барриком и геохимиком Уильямом Шоуэрсом как указание на то, что T. rex поддерживал постоянную внутреннюю температуру тела ( гомеотермию ) и что он обладал метаболизмом где-то между эктотермными рептилиями и эндотермными млекопитающими. [106] Позже они обнаружили похожие результаты у особей Giganotosaurus , которые жили на другом континенте и на десятки миллионов лет раньше по времени. [107] Даже если Tyrannosaurus rex действительно демонстрирует признаки гомеотермии, это не обязательно означает, что он был эндотермическим. Такая терморегуляция также может быть объяснена гигантотермией , как у некоторых современных морских черепах . [108] [109] [110]

В формации Dinosaur Park горгозавр жил рядом с более редким видом тираннозавров Daspletosaurus . Это один из немногих примеров сосуществования двух родов тираннозавров. Хищники схожего размера в современных гильдиях хищников разделены на разные экологические ниши анатомическими, поведенческими или географическими различиями, которые ограничивают конкуренцию. Дифференциация ниш между тираннозаврами Dinosaur Park не очень хорошо изучена. [111] В 1970 году Дейл Рассел выдвинул гипотезу , что более распространенные горгозавры активно охотились на быстроногих гадрозавров , в то время как более редкие и более проблемные цератопсы и анкилозавры (рогатые и тяжелобронированные динозавры ) были предоставлены более крепко сложенным дасплетозаврам . [12] Однако образец Daspletosaurus (OTM 200) из современной ему формации Ту-Медисин в Монтане сохранил переваренные останки молодого гадрозавра в области кишечника. [112] В отличие от некоторых других групп динозавров, ни один из родов не был более распространен на более высоких или низких высотах, чем другой. [111] Однако Gorgosaurus , по -видимому, более распространен в северных формациях, таких как Парк динозавров, а виды Daspletosaurus более многочисленны на юге. Та же картина наблюдается и в других группах динозавров. Цератопсы-хасмозавры и гадрозаврины-гадрозавры также более распространены в формации Ту-Медисин в Монтане и на юго-западе Северной Америки в кампанском веке, в то время как центрозаврины и ламбеозаврины доминируют в северных широтах. Хольц предположил, что эта картина указывает на общие экологические предпочтения между тираннозавринами, хасмозавринами и гадрозавринами. В конце позднего маастрихтского яруса тираннозаврины, такие как Tyrannosaurus rex , гадрозаврины, такие как Edmontosaurus , и хасмозаврины, такие как Triceratops, были широко распространены по всей западной части Северной Америки, в то время как альбертозаврины и центрозаврины вымерли, а ламбеозаврины были редки. [10]
Существуют ограниченные свидетельства социального поведения среди тираннозавридов. Исследователи сообщили, что в том же карьере, что и образец «Сью», были найдены скелеты полувзрослой и молодой особи, что использовалось для подтверждения гипотезы о том, что тираннозавры могли жить в каких-то социальных группах. [113] Хотя нет никаких свидетельств стадного поведения у горгозавра , [61] [62] есть свидетельства некоторого стайного поведения у альбертозавра и дасплетозавра .
Молодой экземпляр вида Dinosaur Park Daspletosaurus ( TMP 94.143.1) демонстрирует следы укусов на лице, нанесенные другим тираннозавром. Следы укусов зажили, что указывает на то, что животное пережило укус. Взрослый экземпляр Dinosaur Park Daspletosaurus (TMP 85.62.1) также демонстрирует следы укусов тираннозавра, показывающие, что атаки на лицо не ограничивались молодыми животными. Хотя возможно, что укусы были приписаны другим видам, внутривидовая агрессия, включая укусы лица, очень распространена среди хищников. Укусы лица наблюдаются у других тираннозавров, таких как Gorgosaurus и Tyrannosaurus , а также у других родов теропод, таких как Sinraptor и Saurornitholestes . Даррен Танке и Фил Карри выдвигают гипотезу, что укусы вызваны внутривидовой конкуренцией за территорию или ресурсы или за доминирование в социальной группе. [61]

Доказательства того, что дасплетозавры жили социальными группами, получены из костного слоя, найденного в формации Ту-Медисин в Монтане. Костный слой включает останки трех дасплетозавров , включая крупного взрослого, маленького детеныша и еще одну особь среднего размера. По крайней мере пять гадрозавров сохранились в одном месте. Геологические данные указывают на то, что останки не были принесены вместе речными течениями , а что все животные были захоронены одновременно в одном месте. Останки гадрозавров разбросаны и имеют множество отметин от зубов тираннозавра, что указывает на то, что дасплетозавры питались гадрозаврами во время смерти. Причина смерти неизвестна. Карри предполагает, что дасплетозавры образовали стаю , хотя это нельзя утверждать с уверенностью. [62] Другие ученые скептически относятся к доказательствам существования социальных групп у дасплетозавров и других крупных теропод; [114] Брайан Роуч и Дэниел Бринкман предположили, что социальное взаимодействие дасплетозавров больше напоминало бы современное взаимодействие комодских варанов , где не склонные к сотрудничеству особи нападают на туши, часто нападая и даже поедая друг друга в процессе. [115]
Костеносный слой острова Драй-Айленд, обнаруженный Барнумом Брауном и его командой, содержит останки 22 альбертозавров , наибольшее количество особей, найденных в одном месте среди всех меловых тероподов, и второе по количеству среди всех крупных динозавров-тероподов после скопления аллозавров в карьере динозавров Кливленд-Ллойд в штате Юта . Группа, по-видимому, состоит из одной очень старой взрослой особи; восьми взрослых в возрасте от 17 до 23 лет; семи полувзрослых особей, проходящих быстрые фазы роста в возрасте от 12 до 16 лет; и шести молодых особей в возрасте от 2 до 11 лет, которые еще не достигли фазы роста. [53] Почти полное отсутствие останков травоядных и схожая степень сохранности многих особей в карьере с костями альбертозавра привели Фила Карри к выводу, что это место не было ловушкой для хищников, как в La Brea Tar Pits в Калифорнии , и что все сохранившиеся животные умерли в одно и то же время. Карри утверждает, что это доказательство стайного поведения. [116] Другие ученые настроены скептически, отмечая, что животные могли быть согнаны вместе засухой, наводнением или по другим причинам. [53] [114] [117]
Хотя в целом это остается спорным, существуют доказательства, подтверждающие теорию о том, что по крайней мере некоторые тираннозавриды были социальными. В формации Вапити в Британской Колумбии местным охотником по имени Аарон Фредлунд была обнаружена дорожка следов, состоящая из отпечатков ног трех отдельных тираннозавридов (названных ихногеном Bellatoripes fredlundi ), которая была описана в журнале PLOS One Ричардом МакКри и др. Исследование дорожки следов не обнаружило никаких доказательств того, что одна дорожка следов была оставлена намного позже другой, что еще раз подтверждает гипотезу о том, что три отдельных тираннозавра путешествовали вместе как группа. Дальнейшие исследования показали, что животные двигались со скоростью от 3,9 до 5,2 миль в час (от 6,3 до 8,4 км/ч) и, вероятно, имели высоту бедра около 7-9 футов. Поскольку из этой формации известны три разных рода тираннозавридов ( Gorgosaurus , Daspletosaurus и Albertosaurus соответственно), неизвестно, какой род оставил следы. [118] [119] [120] Дополнительные доказательства в виде костного слоя из карьера Рэйнбоус и Юникорнс в формации Кайпаровиц в Южной Юте, описанного в 2021 году и приписываемого Тератофонею, предполагают, что другие тираннозавриды также были социальными животными. Окаменелости, состоящие из четырех или, возможно, пяти разных животных в возрасте от 4 до 22 лет, предполагают массовую гибель, возможно, вызванную наводнением или, что менее вероятно, токсикозом цианобактерий, пожаром или засухой. Тот факт, что все сохранившиеся животные, по-видимому, погибли в течение короткого промежутка времени, еще больше усиливает аргумент в пользу стадного поведения у тираннозавридов, поскольку костные отложения таких родов, как Teratophoneus , Albertosaurus , Tyrannosaurus и Daspletosaurus, демонстрируют предполагаемое социальное поведение, которое могло быть широко распространено среди тираннозавридов в целом. [121] [122] [123]
Следы зубов тираннозавров являются наиболее часто сохраняемыми следами питания плотоядных динозавров. [124] Они были зарегистрированы у цератопсов , гадрозавров и других тираннозавров. [124] Кости тираннозавров со следами зубов составляют около 2% известных окаменелостей с сохранившимися следами зубов. [124] Зубы тираннозавров использовались в качестве захватов для стаскивания мяса с тела, а не для режущих функций, подобных ножам . [125] Модели износа зубов указывают на то, что сложное поведение встряхивания головой могло быть задействовано при питании тираннозавров. [125]

Предположения о привычках альбертозавра к стайной охоте были высказаны несколькими исследователями, которые предполагают, что молодые члены стаи могли быть ответственны за то, чтобы загонять свою добычу к взрослым особям, которые были крупнее и сильнее, но также и медленнее. [116] Молодь также могла иметь другой образ жизни, чем взрослые, заполняя ниши хищников между таковыми огромных взрослых особей и более мелких современных теропод, самые крупные из которых были на два порядка меньше взрослого альбертозавра по массе. [10] Однако, поскольку сохранение поведения в ископаемой летописи чрезвычайно редко, эти идеи не могут быть легко проверены. Фил Карри предполагает, что дасплетозавры формировали стаи для охоты , хотя это нельзя утверждать с уверенностью. [62] Нет никаких свидетельств такого стадного поведения у горгозавра . [61] [62]
Споры о том, был ли тираннозавр хищником или чистым падальщиком, так же стары, как и споры о его передвижении. Лэмб (1917) описал хороший скелет близкого родственника тираннозавра горгозавра и пришел к выводу , что он, а следовательно, и тираннозавр, был чистым падальщиком, потому что зубы горгозавра почти не имели признаков износа. [126] Этот аргумент больше не воспринимается всерьез, потому что тероподы довольно быстро заменяли свои зубы. С момента первого открытия тираннозавра большинство ученых согласились, что он был хищником, хотя, как и современные крупные хищники, он был бы счастлив съесть или украсть добычу другого хищника, если бы у него была такая возможность. [127] [128]
Известный эксперт по гадрозаврам Джек Хорнер в настоящее время является главным сторонником идеи о том, что тираннозавр был исключительно падальщиком и вообще не занимался активной охотой. [66] [129] [130] Хорнер представил несколько аргументов в поддержку гипотезы чистого падальщика. Наличие больших обонятельных луковиц и обонятельных нервов предполагает высокоразвитое обоняние для вынюхивания туш на больших расстояниях. Зубы могли дробить кости и, следовательно, могли извлекать как можно больше пищи ( костного мозга ) из остатков туши, обычно наименее питательных частей. По крайней мере, часть его потенциальной добычи могла быстро передвигаться, в то время как данные свидетельствуют о том, что тираннозавр ходил, а не бегал. [129] [131]
Другие данные свидетельствуют об охотничьем поведении тираннозавра . Глазницы тираннозавров расположены таким образом, что глаза смотрят вперед, что дает им бинокулярное зрение немного лучше, чем у современных ястребов . Повреждения, нанесенные тираннозаврами, были обнаружены на скелетах гадрозавров и трицератопсов , которые, казалось, пережили первоначальные атаки. [132] [133] [134] Некоторые исследователи утверждают, что если тираннозавр был падальщиком, то другим динозавром должен был быть главный хищник в амеразийском верхнем мелу. Главной добычей были более крупные маргиноцефалы и орнитоподы . Другие тираннозавриды имеют так много общих характеристик с тираннозавром , что только небольшие дромеозавры остаются возможными главными хищниками. В этом свете приверженцы гипотезы падальщиков предположили, что размер и сила тираннозавров позволяли им красть добычу у более мелких хищников. [131]

В 2023 году в формации Dinosaur Park был обнаружен молодой Gorgosaurus с содержимым желудка in situ , содержащим двух молодых особей Citipes возрастом около года . На момент смерти этому молодому особи было 5-7 лет, его длина составляла около 4 метров (13 футов), а вес — около 335 килограммов (739 фунтов). Он намного крупнее двух молодых особей Citipes , которые весят около 9–12 килограммов (20–26 фунтов), что противоречит предположению о том, что тираннозавриды питались добычей своего размера, когда достигали веса 16–32 килограмма (35–71 фунт), что указывает на то, что молодые тираннозавриды все еще потребляли гораздо более мелкую добычу после превышения определенного порогового размера. Открытие этого образца показало, что тираннозавриды, вероятно, не охотились стаями из нескольких поколений, поскольку размер его добычи слишком мал, чтобы делиться с сородичами. Это также прямое диетическое доказательство, которое подкрепляет теорию «онтогенетического диетического сдвига» для тираннозавридов, как ранее предполагалось с помощью экологического моделирования и анатомических особенностей среди разных возрастных групп. Только остатки задних конечностей и хвостовых позвонков молодых особей Citipes присутствовали в брюшной полости тираннозаврида, что предполагает, что молодые особи Gorgosaurus могли преимущественно потреблять мускулистые задние конечности. [135]
Доказательства также убедительно свидетельствуют о том, что тираннозавриды были по крайней мере иногда каннибалами. Сам тираннозавр имеет убедительные доказательства, указывающие на то, что он был каннибалом, по крайней мере, в способности питаться падалью, основанные на следах зубов на костях стопы, плечевой кости и плюсневых костях одного образца. [136] Ископаемые останки из формации Фрутленд , формации Киртланд (обе кампанского возраста) и формации Охо-Аламо маастихтского возраста предполагают, что каннибализм присутствовал у различных родов тираннозавридов бассейна Сан-Хуан. Доказательства, полученные из образцов, предполагают оппортунистическое пищевое поведение у тираннозавридов, которые поедали членов своего собственного вида. [137]

Самые ранние известные останки тиранозавра были найдены между 129,4 и 125 миллионами лет назад в Исикаве, Япония, около широты 36.166668 долготы 136.633331. Найден Маршем в 1881 году и идентифицирован Х. Ф. Осборном в 1906 году, возраст останков был определен Х. Мацуокой и др. в 2002 году.
В то время как более ранние тираннозавроиды были найдены на всех трех северных континентах, окаменелости тираннозавридов известны только из Северной Америки и Азии. Иногда фрагментарные останки, обнаруженные в Южном полушарии, сообщались как «тираннозавриды Южного полушария», хотя, похоже, это были неправильно идентифицированные окаменелости абелизаврид . [138]
Останки тираннозавридов никогда не были обнаружены в восточной части Северной Америки, в то время как более базальные тираннозавроиды, такие как Dryptosaurus и Appalachiosaurus , сохранялись там до конца мелового периода, что указывает на то, что тираннозавриды, должно быть, эволюционировали или расселились по западной части Северной Америки после того, как континент был разделен пополам Западным внутренним проливом в середине мелового периода. [16] Окаменелости тираннозавридов были найдены на Аляске , которая, возможно, обеспечила путь для распространения между Северной Америкой и Азией. [139] В одном кладистическом анализе обнаружено, что Alioramus и Tarbosaurus связаны, образуя уникальную азиатскую ветвь семейства. [18] Позднее это было опровергнуто открытием Qianzhousaurus и описанием семейства тираннозавров Alioramini. Зубы тираннозавридов крупного вида неизвестного вида были обнаружены на полуострове Нагасаки исследователями из Музея динозавров префектуры Фукуи, что еще больше расширило ареал группы. Возраст зубов оценивается в 81 миллион лет ( кампанский век). [140]

Из двух подсемейств тираннозаврины, по-видимому, были более распространены. Альбертозаврины неизвестны в Азии, которая была домом для тираннозавринов, таких как Tarbosaurus и Zhuchengtyrannus , а также Qianzhousaurus и Alioramus из Alioramini. Оба подсемейства Tyrannosaurinae и Albertosaurinae присутствовали в кампанском и раннем маастрихтском ярусах Северной Америки, причем тираннозаврины, такие как Daspletosaurus, обитали по всей Западной внутренней части, в то время как альбертозаврины Albertosaurus и Gorgosaurus в настоящее время известны только из северо-западной части континента. [141]
К концу маастрихта альбертозаврины, по-видимому, вымерли, в то время как тираннозаврины Tyrannosaurus бродили от Саскачевана до Техаса . Эта картина отражена в других таксонах североамериканских динозавров. В кампанском и раннем маастрихте ламбеозаврины гадрозавры и центрозаврины цератопсы были распространены на северо-западе, в то время как гадрозаврины и хасмозаврины были более распространены на юге. К концу мелового периода центрозаврины неизвестны, а ламбеозаврины редки, в то время как гадрозаврины и хасмозаврины были распространены по всей Западной внутренней части. [10] Исследование, опубликованное в журнале Scientific Reports 2 февраля 2016 года Стивом Брусатте, Томасом Карром и др . указывает на то, что в течение позднего маастрихта сам тираннозавр мог быть частично ответственен за вымирание других тираннозавридов в большей части западной части Северной Америки. Исследование показывает, что тираннозавр мог быть иммигрантом из Азии, а не эволюционировал в Северной Америке (возможно, потомком близкородственного тарбозавра ), который вытеснил и превзошел других тираннозавридов. Эта теория дополнительно подтверждается тем фактом, что в известном ареале тираннозавра обнаружено мало или совсем нет других типов тираннозавридов . [142]
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка )Выборочная продолжительность жизни T. rex составила от 2 до 28 лет.(Опечатка: doi :10.1038/nature16487, PMID 26675726, Retraction Watch ) и соответствующие оценки массы тела варьировались от 29,9 до 5654 кг.
{{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite web}}: CS1 maint: бот: исходный статус URL неизвестен ( ссылка ){{cite journal}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite web}}: CS1 maint: архивная копия как заголовок ( ссылка ){{cite web}}: CS1 maint: числовые имена: список авторов ( ссылка )[1]