Остеокласт (от древнегреческого ὀστέον (osteon) «кость» и κλαστός (clastos) «сломанный») — это тип костной клетки , которая разрушает костную ткань . Эта функция имеет решающее значение для поддержания, восстановления и ремоделирования костей позвоночного скелета . Остеокласт разбирает и переваривает композит гидратированного белка и минерала на молекулярном уровне, выделяя кислоту и коллагеназу , этот процесс известен как резорбция кости . Этот процесс также помогает регулировать уровень кальция в крови .
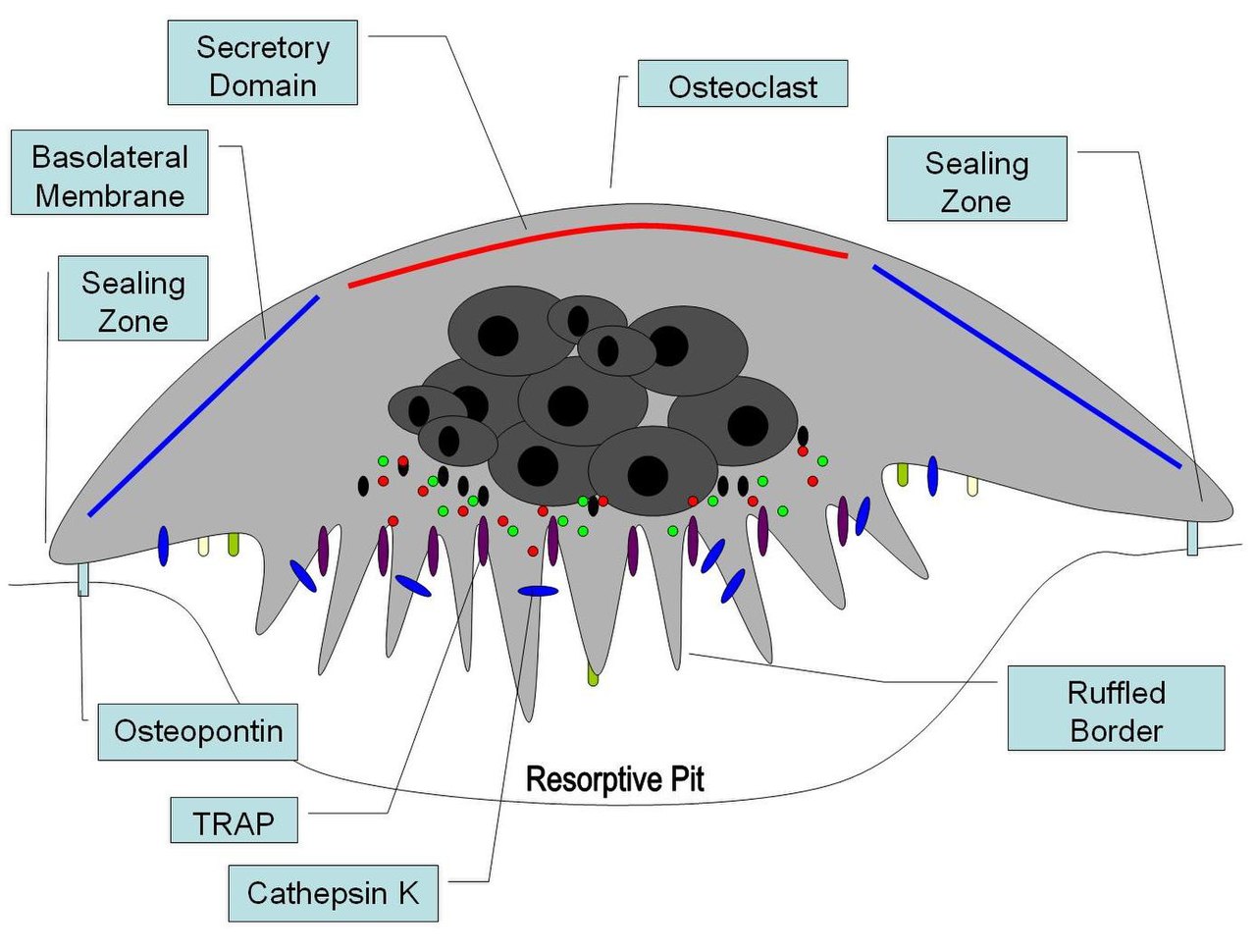
Остеокласты находятся на тех поверхностях кости, которые подвергаются резорбции. На таких поверхностях остеокласты располагаются в неглубоких углублениях, называемых заливами резорбции (лакуны Хаушипа) . Заливы резорбции создаются эрозионным действием остеокластов на лежащую под ними кость. Граница нижней части остеокласта демонстрирует пальцевидные отростки из-за наличия глубоких складок клеточной мембраны ; эта граница называется гофрированной границей . Гофрированная граница лежит в контакте с поверхностью кости в заливе резорбции. Периферия гофрированной границы окружена кольцевой зоной цитоплазмы , которая лишена клеточных органелл , но богата актиновыми нитями . Эта зона называется чистой зоной или зоной уплотнения . Актиновые нити позволяют клеточной мембране, окружающей зону уплотнения, прочно прикрепляться к костной стенке лакун Хаушипа. Таким образом, между гофрированной границей и костью, которая подвергается резорбции, создается закрытый субостеокластический отсек. Остеокласты секретируют ионы водорода , коллагеназу , катепсин К и гидролитические ферменты в этот отсек. Резорбция костного матрикса остеокластами включает два этапа: (1) растворение неорганических компонентов (минералов) и (2) переваривание органического компонента костного матрикса. Остеокласты перекачивают ионы водорода в субостеокластический отсек и таким образом создают кислую микросреду, которая увеличивает растворимость костного минерала, что приводит к высвобождению и повторному поступлению костных минералов в цитоплазму остеокластов для доставки в близлежащие капилляры. После удаления минералов коллагеназа и желатиназа секретируются в субостеокластический отсек. Эти ферменты переваривают и разрушают коллаген и другие органические компоненты декальцинированного костного матрикса. Продукты деградации фагоцитируются остеокластами на гофрированной границе. Из-за своих фагоцитарных свойств остеокласты считаются компонентом системы мононуклеарных фагоцитов (MPS). Активность остеокластов контролируется гормонами и цитокинами. Кальцитонин, гормон щитовидной железы, подавляет активность остеокластов. Остеокласты не имеют рецепторов для паратиреоидного гормона (ПТГ). Однако ПТГ стимулирует остеобласты к секреции цитокина, называемого фактором стимуляции остеокластов, который является мощным стимулятором активности остеокластов. [1]
Одонтокласт (/odon·to·clast/; o-don´to-klast) — это остеокласт, связанный с абсорбцией корней молочных зубов . [2] [3] [4]

Остеокласт — это крупная многоядерная клетка , а человеческие остеокласты на кости обычно имеют четыре ядра [5] и диаметр 150–200 мкм. Когда цитокины, индуцирующие остеокласты, используются для преобразования макрофагов в остеокласты, возникают очень крупные клетки, которые могут достигать 100 мкм в диаметре. Они могут иметь десятки ядер и, как правило, экспрессируют основные белки остеокластов, но имеют значительные отличия от клеток в живой кости из-за неестественного субстрата. [6] [7] Размер многоядерного собранного остеокласта позволяет ему сосредоточить возможности транспорта ионов, секреции белков и везикулярного транспорта многих макрофагов на локализованной области кости.
В костях остеокласты находятся в углублениях на поверхности кости, которые называются резорбционными заливами или лакунами Хоушипа . Остеокласты характеризуются цитоплазмой с гомогенным, «пенистым» видом. Этот вид обусловлен высокой концентрацией везикул и вакуолей . Эти вакуоли включают лизосомы, заполненные кислой фосфатазой . Это позволяет характеризовать остеокласты путем их окрашивания на высокую экспрессию тартрат -устойчивой кислой фосфатазы (TRAP) и катепсина K. Шероховатый эндоплазматический ретикулум остеокластов разрежен, а комплекс Гольджи обширен. [8] [9] [10]
В месте активной резорбции кости остеокласт образует специализированную клеточную мембрану , «гофрированную каёмку», которая противостоит поверхности костной ткани. Эта сильно складчатая или гофрированная каёмка облегчает удаление кости, резко увеличивая поверхность клетки для секреции и поглощения содержимого отсека резорбции и является морфологической характеристикой остеокласта, который активно резорбирует кость.
С момента их открытия в 1873 году было много споров об их происхождении. Доминировали три теории: с 1949 по 1970 год была популярна теория происхождения из соединительной ткани, которая утверждала, что остеокласты и остеобласты имеют одно и то же происхождение, а остеобласты сливаются вместе, образуя остеокласты. После многих лет споров теперь ясно, что эти клетки развиваются из самослияния макрофагов. [11] Именно в начале 1980-х годов фагоцитарная система моноцитов была признана предшественником остеокластов. [12] Для образования остеокластов требуется присутствие RANKL (рецепторный активатор лиганда ядерного фактора κβ) и M-CSF (фактор стимуляции колоний макрофагов) . Эти связанные с мембраной белки продуцируются соседними стромальными клетками и остеобластами , поэтому требуется прямой контакт между этими клетками и предшественниками остеокластов .
M-CSF действует через свой рецептор на остеокласте, c-fms ( рецептор колониестимулирующего фактора 1 ), трансмембранный рецептор тирозинкиназы , что приводит к активации вторичного мессенджера тирозинкиназы Src. Обе эти молекулы необходимы для остеокластогенеза и широко вовлечены в дифференциацию клеток, полученных из моноцитов/макрофагов.
RANKL является членом семейства генов некроза опухолей ( TNF ) и играет важную роль в остеокластогенезе. У мышей с нокаутом RANKL наблюдается фенотип остеопетроза и дефекты прорезывания зубов, а также отсутствие или дефицит остеокластов. RANKL активирует NF-κβ (ядерный фактор-κβ) и NFATc1 (ядерный фактор активированных Т-клеток, цитоплазматический, кальциневрин-зависимый 1) через RANK . Активация NF-κβ стимулируется почти сразу после взаимодействия RANKL-RANK и не повышается. Однако стимуляция NFATc1 начинается примерно через 24–48 часов после связывания, и было показано, что его экспрессия зависит от RANKL.
Дифференциация остеокластов ингибируется остеопротегерином (OPG), который вырабатывается остеобластами и связывается с RANKL, тем самым предотвращая взаимодействие с RANK. В то время как остеокласты происходят из гемопоэтической линии, остеобласты происходят из мезенхимальных стволовых клеток. [13] [14]
После активации остеокласты перемещаются в области микропереломов в кости посредством хемотаксиса . Остеокласты лежат в небольших полостях, называемых лакунами Хоушипа, которые образуются в результате переваривания подлежащей кости. Зона уплотнения представляет собой прикрепление плазматической мембраны остеокласта к подлежащей кости. Зоны уплотнения ограничены поясами специализированных адгезионных структур, называемых подосомами . Прикрепление к костному матриксу облегчается рецепторами интегрина, такими как αvβ3, через специфический аминокислотный мотив Arg-Gly-Asp в белках костного матрикса, таких как остеопонтин . Остеокласт высвобождает ионы водорода посредством действия карбоангидразы ( H2O + CO2 → HCO3 − + H + ) через гофрированную границу в резорбтивную полость, подкисляя и способствуя растворению минерализованного костного матрикса в Ca2 + , H3PO4 , H2CO3 , воду и другие вещества. Было задокументировано , что дисфункция карбоангидразы вызывает некоторые формы остеопетроза. Ионы водорода перекачиваются против градиента высокой концентрации протонными насосами , в частности уникальной вакуолярной АТФазой . Этот фермент был направлен на профилактику остеопороза . Кроме того, несколько гидролитических ферментов , таких как члены групп катепсина и матриксной металлопротеазы (ММП), высвобождаются для переваривания органических компонентов матрикса. Эти ферменты высвобождаются в отсек лизосомами . Из этих гидролитических ферментов наибольшее значение имеет катепсин К.
Катепсин К — коллагенолитическая папаиноподобная цистеиновая протеаза , которая в основном экспрессируется в остеокластах и секретируется в резорбтивную ямку. Катепсин К — основная протеаза, участвующая в деградации коллагена I типа и других неколлагеновых белков. Мутации в гене катепсина К связаны с пикнодизостозом , наследственным остеопетрозным заболеванием, характеризующимся отсутствием функциональной экспрессии катепсина К. Исследования нокаута катепсина К у мышей приводят к остеопетрозному фенотипу, который частично компенсируется повышенной экспрессией протеаз, отличных от катепсина К, и усиленным остеокластогенезом.
Катепсин К имеет оптимальную ферментативную активность в кислых условиях. Он синтезируется как профермент с молекулярной массой 37 кДа и при активации путем автокаталитического расщепления трансформируется в зрелую активную форму с молекулярной массой ~27 кДа.
При поляризации остеокласта над местом резорбции катепсин К секретируется из гофрированной границы в резорбтивную ямку. Катепсин К трансмигрирует через гофрированную границу с помощью межклеточных пузырьков и затем высвобождается функциональным секреторным доменом . Внутри этих межклеточных пузырьков катепсин К вместе с активными формами кислорода , генерируемыми TRAP , дополнительно разрушает внеклеточный матрикс кости.
Несколько других катепсинов экспрессируются в остеокластах, включая катепсины B , C , D , E , G и L. Функция этих цистеиновых и аспарагиновых протеаз в костях, как правило, неизвестна, и они экспрессируются на гораздо более низких уровнях, чем катепсин K.
Исследования мышей с нокаутированным катепсином L дали неоднозначные результаты: в одном отчете сообщалось об уменьшении трабекулярной кости у гомозиготных и гетерозиготных мышей с нокаутированным катепсином L по сравнению с мышами дикого типа, а в другом отчете не было обнаружено никаких аномалий скелета.
Матриксные металлопротеиназы (ММП) представляют собой семейство из более чем 20 цинк-зависимых эндопептидаз. Роль матриксных металлопротеиназ (ММП) в биологии остеокластов плохо определена, но в других тканях они связаны с деятельностью, способствующей развитию опухолей, такой как активация факторов роста , и необходимы для метастазирования опухолей и ангиогенеза.
MMP9 связан с микросредой костей. Он экспрессируется остеокластами и, как известно, необходим для миграции остеокластов и является мощной желатиназой. У трансгенных мышей, у которых отсутствует MMP-9, развиваются дефекты развития костей, внутрикостного ангиогенеза и заживления переломов.
Считается, что ММП-13 участвует в резорбции костей и дифференцировке остеокластов, поскольку у мышей с нокаутированным геном наблюдалось снижение количества остеокластов, остеопетроз и снижение резорбции костей.
ММП, экспрессируемые остеокластом, включают ММП-9, -10, -12 и -14. За исключением ММП-9, мало что известно об их связи с остеокластом, однако в зоне уплотнения обнаружены высокие уровни ММП-14.
В 1980-х и 90-х годах физиология типичных остеокластов была подробно изучена. С выделением гофрированной границы ионный транспорт через нее был изучен непосредственно в биохимических деталях. Энергозависимый транспорт кислоты был проверен, и постулированный протонный насос очищен. [15] [16] С успешным культивированием остеокластов стало очевидно, что они организованы для поддержки массивного транспорта протонов для подкисления отсека резорбции и солюбилизации костного минерала. Это включает проницаемость гофрированной границы Cl − для контроля мембранного потенциала и базолатеральный обмен Cl − /HCO 3 − для поддержания цитозольного pH в физиологически приемлемых диапазонах. [17] [18] [19]
Эффективность его ионной секреции зависит от того, образует ли остеокласт эффективное уплотнение вокруг отсека резорбции. Позиционирование этой «зоны уплотнения», по-видимому, опосредовано интегринами, экспрессируемыми на поверхности остеокласта. [20] При наличии зоны уплотнения многоядерный остеокласт реорганизуется. Развитие высокоинвагинированной гофрированной мембраны, противостоящей отсеку резорбции, обеспечивает массивную секреторную активность. Кроме того, это обеспечивает везикулярный трансцитоз минерала и деградированного коллагена из гофрированной границы в свободную мембрану клетки и его высвобождение во внеклеточный отсек. [21] [22] Эта активность завершает резорбцию кости, и как минеральные компоненты, так и фрагменты коллагена высвобождаются в общий кровоток.
Остеокласты регулируются несколькими гормонами , включая паратиреоидный гормон (ПТГ) из паращитовидной железы, кальцитонин из щитовидной железы и фактор роста интерлейкин 6 (ИЛ-6). Этот последний гормон, ИЛ-6 , является одним из факторов заболевания остеопорозом . Остеопороз возникает, когда существует дисбаланс между активностью остеокластов по резорбции костной ткани и активностью остеобластов по формированию костной ткани. [23]
Активность остеокластов также опосредована взаимодействием двух молекул, продуцируемых остеобластами, а именно остеопротегерина и лиганда RANK . Эти молекулы также регулируют дифференциацию остеокластов. [24] [25]
Одонтокласт — это остеокласт , связанный с абсорбцией корней молочных зубов . [2] [3] [4]
Остеокласт также может быть инструментом, используемым для перелома и вправления костей (происхождение от греческого osteon : кость и klastos : сломанный). Чтобы избежать путаницы, клетку изначально называли осотокластом. Когда хирургический инструмент вышел из употребления, клетка стала известна под своим нынешним названием.
Гигантские остеокласты могут возникать при некоторых заболеваниях, включая болезнь костей Педжета и отравление бисфосфонатами .
У кошек аномальная активность одонтокластов может вызвать одонтокластические резорбтивные поражения , требующие удаления пораженных зубов.
Остеокласты играют важную роль в ортодонтическом перемещении зубов и патологической миграции зубов с поражением пародонта.
Остеокласты были открыты Кёлликером в 1873 году. [12]