Therapsida [a] — клада, включающая в себя основную группу эупеликозавровых синапсид , включающую млекопитающих , их предков и близких родственников. Многие из черт, которые сегодня считаются уникальными для млекопитающих, возникли у ранних терапсид, включая конечности, которые были ориентированы больше под телом, что привело к более «стоячей» четвероногой позе, в отличие от более низкой раскинутой позы многих рептилий и амфибий .
Терапсиды произошли от более ранних синапсидов, обычно называемых « пеликозаврами », в частности, в пределах Sphenacodontia , более 279,5 миллионов лет назад. Они заменили пеликозавров в качестве доминирующих крупных наземных животных в гваделупском периоде вплоть до раннего триаса. После пермско-триасового вымирания терапсиды потеряли относительную значимость по сравнению с быстро диверсифицирующимися архозавровыми завропсидами ( псевдозухиями , динозаврами и птерозаврами и т. д.) в течение среднего триаса.
Терапсиды включают цинодонтов , группу, которая дала начало млекопитающим ( Mammaliaformes ) в позднем триасе около 225 миллионов лет назад, единственную кладу терапсид, которая выжила после окончания триаса . Единственная другая группа терапсид, которая выжила в позднем триасе , дицинодонты , вымерли к концу периода. Последней выжившей группой немаммалистных цинодонтов были Tritylodontidae , которые вымерли в раннем мелу .

Височные окна терапсид были больше, чем у пеликозавров. Челюсти некоторых терапсид были более сложными и мощными, а зубы были дифференцированы на фронтальные резцы для кусания, большие боковые клыки для прокалывания и разрывания и моляры для разрезания и измельчения пищи.
Ноги терапсидов располагались более вертикально под их телами, чем раскинутые ноги рептилий и пеликозавров. Также по сравнению с этими группами, ступни были более симметричными, с короткими первым и последним пальцами и длинными средними пальцами, что указывает на то, что ось ступни была расположена параллельно оси животного, а не раскинулась в стороны. Такая ориентация придала бы походке больше млекопитающего , чем походке ящерицы пеликозавров. [2]
Физиология терапсид плохо изучена. У большинства пермских терапсид было пинеальное отверстие, что указывает на то, что у них был теменной глаз, как у многих современных рептилий и амфибий. Теменной глаз играет важную роль в терморегуляции и циркадном ритме эктотермных животных, но отсутствует у современных млекопитающих, которые являются эндотермными . [3] Ближе к концу перми дицинодонты, тероцефалы и цинодонты демонстрируют параллельные тенденции к утрате пинеального отверстия, а у пробайногнатных цинодонтов отверстие полностью отсутствует. Данные изотопов кислорода, которые коррелируют с температурой тела, свидетельствуют о том, что большинство пермских терапсид были эктотермными животными и что эндотермия развилась конвергентно у дицинодонтов и цинодонтов ближе к концу перми. [4] Напротив, данные гистологии показывают, что эндотермия свойственна всем представителям Therapsida, [5] тогда как оценки скорости кровотока и продолжительности жизни млекопитающих Morganucodon показывают, что даже ранние млекопитающие имели скорость метаболизма, подобную скорости метаболизма рептилий. [6] Доказательства наличия дыхательных носовых раковин, которые, как предполагалось, являются признаком эндотермии, были зарегистрированы у тероцефала Glanosuchus , но последующее исследование показало, что очевидные места прикрепления носовых раковин могут быть просто результатом деформации черепа. [7]
Эволюция покровов у терапсид плохо изучена, и существует мало ископаемых, которые предоставляют прямые доказательства наличия или отсутствия меха. Наиболее базальные синапсиды с недвусмысленными прямыми доказательствами наличия меха - это докодонты , которые являются формами млекопитающих, очень тесно связанными с млекопитающими группы крон. Два "мумифицированных" молодых образца дицинодонта Lystrosaurus murrayi сохранили отпечатки кожи; кожа безволосая, кожистая и ямчатая, несколько сравнимая с кожей слона. [8] [9] Окаменевшая кожа лица диноцефала Estemmenosuchus была описана как показывающая, что кожа была железистой и не имела как чешуи, так и волос. [10]
Копролиты, содержащие нечто похожее на волосы, были обнаружены в позднепермских отложениях . [11] [12] Хотя источник этих волос точно неизвестен, они могут указывать на то, что волосы присутствовали по крайней мере у некоторых пермских терапсид.
Закрытие пинеального отверстия у пробайногнатных цинодонтов может указывать на мутацию в регуляторном гене Msx2, который участвует как в закрытии крыши черепа, так и в поддержании волосяных фолликулов у мышей. [13] Это говорит о том, что волосы могли впервые развиться у пробайногнатных, хотя это не полностью исключает более раннее происхождение меха. [13]
Усы, вероятно, развились у пробайногнатных цинодонтов. [13] [14] Некоторые исследования предполагали более раннее происхождение усов на основе наличия отверстий на морде тероцефалов и ранних цинодонтов, но расположение отверстий в этих таксонах на самом деле очень напоминает ящериц, [15] что делает маловероятным наличие млекопитающеподобных усов. [14]

Терапсиды произошли от группы пеликозавров, называемых сфенакодонтами . [17] [18] Терапсиды стали доминирующими наземными животными в средней перми , вытеснив пеликозавров. Терапсиды состоят из четырех основных клад : диноцефалы , травоядные аномодонты , плотоядные биармозухии и в основном плотоядные териодонты . После краткого всплеска эволюционного разнообразия диноцефалы вымерли в поздней средней перми ( гваделупском ярусе ), но аномодонтные дицинодонты, а также териодонтные горгонопсии и тероцефалы процветали, и в самом конце перми к ним присоединились первые цинодонты .

Как и все наземные животные, терапсиды серьезно пострадали от пермско-триасового вымирания , когда очень успешные горгонопсии и биармозухии полностью вымерли, а оставшиеся группы — дицинодонты , тероцефалы и цинодонты — сократились до нескольких видов каждая к самому раннему триасу . Выжившие дицинодонты были представлены двумя семействами таксонов катастроф ( Lystrosauridae и Myosauridae ), малоизвестной Kombuisia и единственной группой крупных коренастых травоядных , Kannemeyeriiformes , которые были единственной линией дицинодонтов, процветавшей в триасе. [20] Они и цинодонты среднего размера (включая как плотоядные, так и травоядные формы) процветали по всему миру в течение раннего и среднего триаса. Они исчезают из ископаемых остатков на большей части Пангеи в конце карнийского периода (поздний триас), хотя продолжали существовать еще некоторое время во влажной экваториальной полосе и на юге. [ необходима цитата ]
Некоторые исключения были еще более развитыми эвцинодонтами . [21] По крайней мере три группы из них выжили. Все они появились в позднетриасовый период. Чрезвычайно похожее на млекопитающих семейство, Tritylodontidae , сохранилось в раннем меловом периоде . Другое чрезвычайно похожее на млекопитающих семейство, Tritheledontidae , неизвестно позднее ранней юры . Mammaliaformes были третьей группой, включая Morganucodon и подобных животных. Некоторые таксономисты называют этих животных «млекопитающими», хотя большинство ограничивает этот термин группой млекопитающих .

Неэуцинодонты пережили пермско-триасовое вымирание; Thrinaxodon , Galesaurus и Platycraniellus известны из раннего триаса . Однако к среднему триасу остались только эвцинодонты.
Тероцефалы , родственники цинодонтов, сумели пережить пермско-триасовое вымирание и продолжали диверсифицироваться в течение раннего триасового периода. Однако, приближаясь к концу периода, тероцефалы пришли в упадок и в конечном итоге вымерли, вероятно, вытесненные быстро диверсифицирующейся линией ящеров- диапсидов , оснащенных сложными дыхательными системами, лучше подходящими для очень жаркого, сухого и бедного кислородом мира конца триаса.
Дицинодонты были одной из самых успешных групп терапсид в поздней перми и дожили почти до конца триаса.
Млекопитающие — единственные живущие терапсиды. Группа млекопитающих , которая развилась в раннем юрском периоде, произошла от группы млекопитающих, включавшей докодонтов . Сами млекопитающие произошли от пробайногнатов , линии подотряда эвцинодонтов.
Обычно выделяют шесть основных групп терапсид: Biarmosuchia , Dinocephalia , Anomodontia , Gorgonopsia , Therocephalia и Cynodontia . Клада, объединяющая тероцефалов и цинодонтов, называемая Eutheriodontia , хорошо поддерживается, но отношения между остальными четырьмя кладами являются спорными. [23] Наиболее широко принятая гипотеза о взаимоотношениях терапсид, парадигма Хопсона и Баргхаузена, была впервые предложена в 1986 году. Согласно этой гипотезе, биармозухии являются самой ранней расходящейся основной группой терапсид, а остальные пять групп образуют Eutherapsida, а внутри Eutherapsida горгонопсии являются сестринским таксоном эутериодонтов, вместе образуя Theriodontia . Хопсон и Баргхаузен изначально не пришли к выводу о том, как диноцефалы, аномодонты и териодонты были связаны друг с другом, но последующие исследования показали, что аномодонты и териодонты должны быть классифицированы вместе как Neotherapsida. Тем не менее, споры об этих отношениях продолжаются; в частности, некоторые исследования предполагают, что аномодонты, а не горгонопсии, являются сестринским таксоном Eutheriodontia, другие исследования обнаружили, что диноцефалы и аномодонты образуют кладу, и как филогенетическое положение, так и монофилия Biarmosuchia остаются спорными.
В дополнение к шести основным группам, существует несколько других линий и видов с неопределенной классификацией. Raranimus из ранней средней перми Китая, вероятно, является самым ранним известным терапсидом, дивергировавшим из известных. [24] Tetraceratops из ранней перми Соединенных Штатов был выдвинут в качестве еще более раннего дивергировавшего терапсида, [25] [26] но более позднее исследование показало, что он, скорее всего, является нетерапсидом сфенакодонтом. [27]

Biarmosuchia — это недавно признанная клада терапсид, впервые выделенная в отдельную линию Хопсоном и Баргхаузеном в 1986 году и официально названная Сигоньо-Расселом в 1989 году. Большинство биармозухий ранее классифицировались как горгонопсии. Biarmosuchia включает отличительную Burnetiamorpha , но поддержка монофилии Biarmosuchia относительно низкая. Многие биармозухии известны обширной черепной орнаментацией.

Диноцефалы включают в себя две различные группы: антеозавры и тапиноцефалы .
Исторически плотоядные диноцефалы, включая антеозавров и титанозухидов, назывались титанозухиями и относились к териодонтам, в то время как травоядные тапиноцефалы относились к аномодонтам.
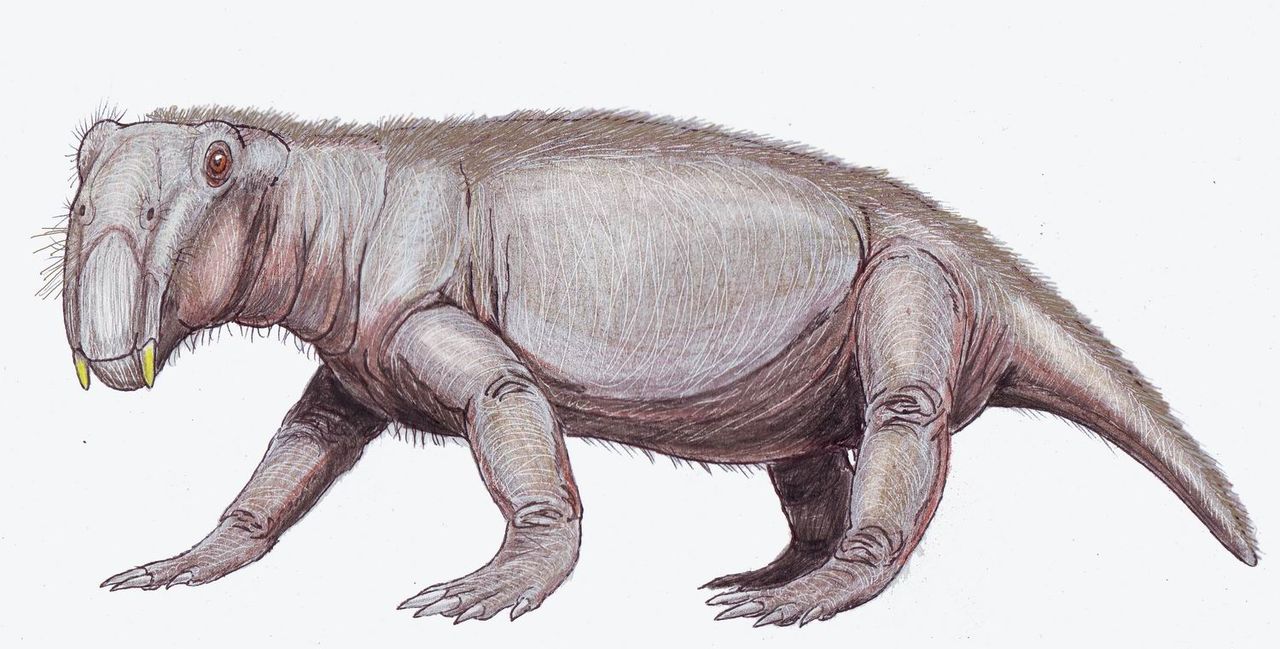
Anomodontia включает дицинодонтов , кладу травоядных с бивнями и клювами, и самую разнообразную и долгоживущую кладу нецинодонтовых терапсид. Другие члены Anomodontia включают суминию , которая, как полагают, была лазающей формой.

Горгонопсии — многочисленная, но морфологически однородная группа саблезубых хищников .

Было высказано предположение, что Therocephalia может не быть монофилетическим, поскольку некоторые виды более тесно связаны с цинодонтами, чем другие. [28] Однако большинство исследований рассматривают Therocephalia как монофилетическое семейство.

Цинодонты являются наиболее разнообразными и долгоживущими из групп терапсид, поскольку Cynodontia включает млекопитающих . Цинодонты являются единственной крупной кладой терапсид, не имеющей среднепермской ископаемой летописи, причем самым ранним известным цинодонтом является Charassognathus из учапинского яруса поздней перми. Не млекопитающие цинодонты включают как плотоядные, так и травоядные формы.