
Поведение роя , или роение , представляет собой коллективное поведение , демонстрируемое существами, в частности животными, схожего размера, которые собираются вместе, возможно, толпясь в одном месте или, возможно, двигаясь скопом или мигрируя в каком-то направлении. Это весьма междисциплинарная тема. [1]
Как термин, роение применяется в частности к насекомым, но может также применяться к любому другому существу или животному, которое демонстрирует роевое поведение. Термин стайность или бормотание может относиться конкретно к роевому поведению у птиц, стадность — к роевому поведению у четвероногих , а скопление или косячность — к роевому поведению у рыб. Фитопланктон также собирается в огромные рои, называемые цветением , хотя эти организмы являются водорослями и не являются самоходными, как животные. В более широком смысле, термин «рой» применяется также к неодушевленным существам, которые демонстрируют параллельное поведение, как в рое роботов , рое землетрясений или рое звезд.
С более абстрактной точки зрения, роевое поведение представляет собой коллективное движение большого количества самодвижущихся сущностей . [2] С точки зрения математического моделировщика, это возникающее поведение, возникающее из простых правил, которым следуют индивидуумы, и не предполагающее никакой центральной координации. Поведение роя также изучается физиками активной материи как явление, которое не находится в термодинамическом равновесии , и как таковое требует разработки инструментов, выходящих за рамки тех, которые доступны в статистической физике систем в термодинамическом равновесии. В этом отношении роение сравнивают с математикой сверхтекучих жидкостей , особенно в контексте стай скворцов (бормотание). [3]
Поведение роя было впервые смоделировано на компьютере в 1986 году с помощью программы моделирования boids . [4] Эта программа симулирует простых агентов (boids), которым разрешено двигаться в соответствии с набором базовых правил. Первоначально модель была разработана для имитации стайного поведения птиц, но ее можно применять также к стайным рыбам и другим роящимся сущностям.
В последние десятилетия ученые обратились к моделированию поведения роя, чтобы глубже понять его.

Ранние исследования поведения роя использовали математические модели для моделирования и понимания поведения. Простейшие математические модели стай животных обычно представляют отдельных животных следующими тремя правилами:
Компьютерная программа boids , созданная Крейгом Рейнольдсом в 1986 году, имитирует поведение роя, следуя вышеуказанным правилам. [4] Многие последующие и современные модели используют вариации этих правил, часто реализуя их посредством концентрических «зон» вокруг каждого животного. В «зоне отталкивания», очень близко к животному, фокусное животное будет стремиться дистанцироваться от своих соседей, чтобы избежать столкновения. Чуть дальше, в «зоне выравнивания», фокусное животное будет стремиться выровнять свое направление движения со своими соседями. В самой дальней «зоне притяжения», которая простирается так далеко от фокусного животного, как оно может почувствовать, фокусное животное будет стремиться двигаться к соседу.
Форма этих зон обязательно будет зависеть от сенсорных возможностей данного животного. Например, поле зрения птицы не простирается за ее тело. Рыбы полагаются как на зрение, так и на гидродинамические восприятия, передаваемые через их боковые линии , в то время как антарктический криль полагается как на зрение, так и на гидродинамические сигналы, передаваемые через антенны .
Однако недавние исследования стай скворцов показали, что каждая птица изменяет свое положение относительно шести или семи животных, непосредственно окружающих ее, независимо от того, насколько близко или далеко находятся эти животные. [5] Таким образом, взаимодействие между стайными скворцами основано на топологическом , а не метрическом правиле. Еще предстоит выяснить, применимо ли это к другим животным. Другое недавнее исследование, основанное на анализе высокоскоростной съемки стай над Римом и предполагающее минимальные поведенческие правила, убедительно смоделировало ряд аспектов поведения стаи. [6] [7] [8] [9]
Чтобы понять, почему у животных развивается роевое поведение, ученые обратились к эволюционным моделям, которые имитируют популяции эволюционирующих животных. Обычно эти исследования используют генетический алгоритм для моделирования эволюции на протяжении многих поколений. Эти исследования изучили ряд гипотез, пытающихся объяснить, почему у животных развивается роевое поведение, такие как теория эгоистичного стада [10] [11] [12] [13] [14] эффект замешательства хищника, [15] [16] эффект разбавления, [17] [18] и теория многих глаз. [19]

Концепция возникновения — то, что свойства и функции, обнаруженные на иерархическом уровне, отсутствуют и не имеют значения на более низких уровнях — часто является основным принципом самоорганизующихся систем . [20] Пример самоорганизации в биологии , приводящей к возникновению в естественном мире, встречается в колониях муравьев. Королева не отдает прямых приказов и не говорит муравьям, что делать. [ требуется ссылка ] Вместо этого каждый муравей реагирует на стимулы в виде химических запахов от личинок, других муравьев, злоумышленников, пищи и накопления отходов, и оставляет за собой химический след, который, в свою очередь, дает стимул другим муравьям. Здесь каждый муравей является автономной единицей, которая реагирует только в зависимости от своей локальной среды и генетически закодированных правил для своего вида. Несмотря на отсутствие централизованного принятия решений, колонии муравьев демонстрируют сложное поведение и даже смогли продемонстрировать способность решать геометрические задачи. Например, колонии обычно находят максимальное расстояние от всех входов в колонию, чтобы избавиться от мертвых тел.
Еще одной ключевой концепцией в области роевого интеллекта является стигмергия . [21] [22] Стигмергия — это механизм косвенной координации между агентами или действиями. Принцип заключается в том, что след, оставленный в среде действием, стимулирует выполнение следующего действия тем же или другим агентом. Таким образом, последующие действия имеют тенденцию усиливать и основываться друг на друге, что приводит к спонтанному возникновению согласованной, по-видимому, систематической деятельности. Стигмергия — это форма самоорганизации. Она создает сложные, на первый взгляд разумные структуры, без необходимости какого-либо планирования, контроля или даже прямой коммуникации между агентами. Как таковая, она поддерживает эффективное сотрудничество между чрезвычайно простыми агентами, которые лишены какой-либо памяти, интеллекта или даже осведомленности друг о друге. [22]
Роевой интеллект — это коллективное поведение децентрализованных , самоорганизующихся систем, естественных или искусственных . Концепция используется в работе над искусственным интеллектом . Выражение было введено Херардо Бени и Цзин Ваном в 1989 году в контексте клеточных роботизированных систем. [23]
Системы роевого интеллекта обычно состоят из популяции простых агентов, таких как boids, локально взаимодействующих друг с другом и со своей средой. Агенты следуют очень простым правилам, и хотя нет централизованной структуры управления, диктующей, как должны себя вести отдельные агенты, локальные и в определенной степени случайные взаимодействия между такими агентами приводят к появлению интеллектуального глобального поведения, неизвестного отдельным агентам.
Исследования роевого интеллекта являются междисциплинарными. Их можно разделить на исследования естественного роя, изучающего биологические системы, и исследования искусственного роя, изучающего человеческие артефакты. Существует также научное направление, пытающееся смоделировать сами роевые системы и понять их базовые механизмы, и инженерное направление, сосредоточенное на применении идей, разработанных научным направлением, для решения практических проблем в других областях. [24]
Алгоритмы роя следуют лагранжевому подходу или эйлерову подходу. [25] Эйлеров подход рассматривает рой как поле , работая с плотностью роя и выводя средние свойства поля. Это гидродинамический подход, и он может быть полезен для моделирования общей динамики больших роев. [26] [27] [28] Однако большинство моделей работают с лагранжевым подходом, который представляет собой модель на основе агентов, следующих за отдельными агентами (точками или частицами), составляющими рой. Отдельные модели частиц могут следовать информации о направлении и расстоянии, которая теряется в эйлеровом подходе. [25] [29]
Оптимизация колонии муравьев — широко используемый алгоритм, который был вдохновлен поведением муравьев и оказался эффективным при решении задач дискретной оптимизации, связанных с роением. [31] Первоначально алгоритм был предложен Марко Дориго в 1992 году, [32] [33] и с тех пор был расширен для решения более широкого класса численных задач. Виды, имеющие несколько королев, могут иметь королеву, покидающую гнездо вместе с некоторыми рабочими, чтобы основать колонию на новом месте, процесс, схожий с роением у медоносных пчел . [34] [35]
Концепция самодвижущихся частиц (SPP) была введена в 1995 году Тамашем Вичеком и др. [37] как частный случай модели боидов, введенной в 1986 году Рейнольдсом. [4] Рой SPP моделируется совокупностью частиц, которые движутся с постоянной скоростью и реагируют на случайные возмущения, принимая на каждом временном приращении среднее направление движения других частиц в их локальной окрестности. [38]
Моделирование показывает, что подходящее «правило ближайшего соседа» в конечном итоге приводит к тому, что все частицы собираются вместе или движутся в одном направлении. Это происходит, даже если нет централизованной координации, и даже если соседи для каждой частицы постоянно меняются со временем. [37] Модели SPP предсказывают, что роящиеся животные разделяют определенные свойства на уровне группы, независимо от типа животных в рое. [39] Роящиеся системы порождают эмерджентное поведение , которое происходит во многих различных масштабах, некоторые из которых являются как универсальными, так и надежными. В теоретической физике стало проблемой найти минимальные статистические модели, которые охватывают это поведение. [40] [41]
Оптимизация роя частиц — еще один алгоритм, широко используемый для решения проблем, связанных со роями. Он был разработан в 1995 году Кеннеди и Эберхартом и изначально был нацелен на моделирование социального поведения и хореографии стай птиц и косяков рыб. [42] [43] Алгоритм был упрощен, и было замечено, что он выполняет оптимизацию. Система изначально засеивает популяцию случайными решениями. Затем она выполняет поиск в пространстве задач через последовательные поколения, используя стохастическую оптимизацию, чтобы найти наилучшие решения. Найденные ею решения называются частицами . Каждая частица сохраняет свое положение, а также наилучшее решение, которого она достигла до сих пор. Оптимизатор роя частиц отслеживает наилучшее локальное значение, полученное до сих пор любой частицей в локальной окрестности. Затем оставшиеся частицы перемещаются по пространству задач, следуя за оптимальными частицами. На каждой временной итерации оптимизатор роя частиц ускоряет каждую частицу к ее оптимальным положениям в соответствии с простыми математическими правилами . Оптимизация роя частиц применялась во многих областях. Он имеет несколько параметров для настройки, и версия, которая хорошо работает для определенного приложения, может также хорошо работать с небольшими изменениями в ряде связанных приложений. [44] В книге Кеннеди и Эберхарта описываются некоторые философские аспекты приложений оптимизации роя частиц и роевого интеллекта. [45] Обширный обзор приложений сделан Поли. [46] [47]
Исследователи из Швейцарии разработали алгоритм, основанный на правиле родственного отбора Гамильтона . Алгоритм показывает, как альтруизм в рое сущностей может со временем эволюционировать и привести к более эффективному поведению роя. [48] [49]

Самые ранние свидетельства поведения роя у животных датируются примерно 480 миллионами лет. Недавно было описано, что ископаемые останки трилобита Ampyx priscus были сгруппированы в линии вдоль дна океана. Все животные были взрослыми особями и смотрели в одном направлении, как будто они образовали линию конги или пелотон . Было высказано предположение, что они выстраиваются таким образом для миграции, подобно тому, как лангусты мигрируют в одиночные очереди; [50] также было высказано предположение, что формирование является предшественником спаривания, [51] как в случае с мухой Leptoconops torrens . Результаты показывают, что коллективное поведение животных имеет очень раннее эволюционное происхождение. [52]
Примеры биологического роения можно найти в стаях птиц , [53] косяках рыб , [54] [55] роях насекомых , [56] роях бактерий , [57] [58] плесени, [59] молекулярных двигателях , [60] стадах четвероногих [61] и людях. [62] [63] [64] [65]
Поведение социальных насекомых (насекомых, которые живут колониями , таких как муравьи, пчелы, осы и термиты) всегда было источником восхищения для детей, натуралистов и художников. Отдельные насекомые, кажется, делают свое дело без какого-либо центрального контроля, однако колония в целом ведет себя высокоскоординированным образом. [66] Исследователи обнаружили, что сотрудничество на уровне колонии в значительной степени самоорганизовано . Возникающая групповая координация часто является просто следствием того, как взаимодействуют особи в колонии. Эти взаимодействия могут быть удивительно простыми, например, один муравей просто следует по следу, оставленному другим муравьем. Однако, если объединить их, кумулятивный эффект такого поведения может решать весьма сложные проблемы, такие как поиск кратчайшего маршрута в сети возможных путей к источнику пищи. Организованное поведение, которое возникает таким образом, иногда называют роевым интеллектом , формой биологической эмерджентности . [66]
Отдельные муравьи не демонстрируют сложного поведения, однако колония муравьев коллективно выполняет сложные задачи, такие как строительство гнезд, забота о потомстве, строительство мостов и поиск пищи. Колония муравьев может коллективно выбирать (т. е. отправлять большинство рабочих к) лучшему или ближайшему источнику пищи из нескольких поблизости. [67] Такие коллективные решения достигаются с помощью механизмов положительной обратной связи. Выбор лучшего источника пищи достигается муравьями, следующими двум простым правилам. Во-первых, муравьи, которые находят пищу, возвращаются в гнездо, откладывая феромон . Больше феромонов откладывается для более качественных источников пищи. [68] Таким образом, если одновременно найдены два равноудаленных источника пищи разного качества, феромонный след к лучшему из них будет сильнее. Муравьи в гнезде следуют другому простому правилу, отдавая предпочтение в среднем более сильным следам. Затем больше муравьев следуют по более сильному следу, поэтому больше муравьев прибывают к высококачественному источнику пищи, и цикл положительной обратной связи обеспечивает, что приводит к коллективному решению о лучшем источнике пищи. Если есть два пути от муравейника до источника пищи, то колония обычно выбирает более короткий путь. Это происходит потому, что муравьи, которые первыми возвращаются в гнездо от источника пищи, с большей вероятностью будут теми, кто выбрал более короткий путь. Затем больше муравьев повторяют более короткий путь, усиливая феромонный след. [69]
Армейские муравьи , в отличие от большинства видов муравьев, не строят постоянных гнезд; колония армейских муравьев почти непрерывно перемещается в течение всего времени своего существования, оставаясь в по существу постоянном состоянии роения. Несколько линий независимо друг от друга развили один и тот же базовый поведенческий и экологический синдром, часто называемый «легионерским поведением», и могут быть примером конвергентной эволюции . [70]
Успешные методы, используемые колониями муравьев, были изучены в компьютерной науке и робототехнике для создания распределенных и отказоустойчивых систем для решения проблем. Эта область биомиметики привела к исследованиям передвижения муравьев, поисковых систем, которые используют «тропы добычи», отказоустойчивых алгоритмов хранения и сетевых алгоритмов . [71]

В умеренном климате медоносные пчелы обычно образуют рои поздней весной. Рой обычно содержит около половины рабочих вместе со старой королевой, в то время как новая королева остается с оставшимися рабочими в исходном улье. Когда медоносные пчелы выходят из улья, чтобы сформировать рой, они могут собраться на ветке дерева или на кусте всего в нескольких метрах от улья. Пчелы собираются вокруг королевы и отправляют 20–50 разведчиков, чтобы найти подходящие новые места для гнезда. Разведчики являются самыми опытными фуражирами в скоплении. Если разведчик находит подходящее место, он возвращается в скопление и продвигает его, танцуя версию танца виляния . Этот танец передает информацию о качестве, направлении и расстоянии до нового места. Чем больше она взволнована своими находками, тем энергичнее она танцует. Если она может убедить других, что они могут взлететь и проверить место, которое она нашла. Если они одобряют, они также могут продвигать его. В этом процессе принятия решений разведчики проверяют несколько мест, часто отказываясь от своего собственного первоначального места, чтобы продвигать лучшее место другого разведчика. Сначала разные разведчики могут продвигать несколько разных мест. Через несколько часов, а иногда и дней, в результате этого процесса принятия решений в конечном итоге выясняется предпочтительное место. Когда все разведчики соглашаются с окончательным местом, весь кластер взлетает и устремляется к нему. Иногда, если решение не принято, рой разделяется, некоторые пчелы идут в одном направлении, другие — в другом. Обычно это приводит к неудаче, и обе группы погибают. Новое место обычно находится в километре или более от исходного улья, хотя некоторые виды, например, Apis dorsata [ 72], могут основывать новые колонии всего в 500 метрах от натального гнезда. Этот коллективный процесс принятия решений является удивительно успешным в определении наиболее подходящего нового места гнездования и сохранении роя нетронутым. Хорошее место для улья должно быть достаточно большим, чтобы вместить рой (объемом около 15 литров), хорошо защищенным от непогоды, получать оптимальное количество солнечного света, находиться на некоторой высоте над землей, иметь небольшой вход и быть способным противостоять нашествию муравьев — вот почему часто выбирают дупла деревьев. [73] [74] [75] [76] [77]
В отличие от общественных насекомых, стаи необщественных насекомых, которые изучались в первую очередь, по-видимому, функционируют в таких контекстах, как спаривание, питание, избегание хищников и миграция.
Моль может демонстрировать синхронизированное спаривание, во время которого феромоны, выделяемые самками, инициируют поисковое и роевое поведение у самцов. [78] Самцы чувствуют феромоны с помощью чувствительных антенн и могут отслеживать самок на расстоянии нескольких километров. [79] Спаривание роя включает в себя выбор самок и конкуренцию самцов. Только один самец в рое — как правило, первый — успешно совокупляется. [80] Самки максимизируют преимущества приспособленности и минимизируют затраты, регулируя начало и величину высвобождаемого феромона. Слишком мало феромона не привлечет самку, слишком много позволяет менее приспособленным самцам почувствовать сигнал. [81] После спаривания самки откладывают яйца на растение-хозяина. Качество растения-хозяина может быть фактором, влияющим на место роения и откладки яиц. В одном случае исследователи наблюдали, как розовополосые дубовые черви ( Anisota virginiensis ) роились на месте падали , где разложение, вероятно, повышало уровень питательных веществ в почве и качество растения-хозяина. [82]
Мошки, такие как Tokunagayusurika akamusi , образуют рои, танцующие в воздухе. Роение служит нескольким целям, включая содействие спариванию путем привлечения самок к рою, явление, известное как спаривание лек . Такие похожие на облака рои часто образуются ранним вечером, когда солнце садится, на верхушке куста, на вершине холма, над водоемом или даже иногда над человеком. Формирование таких роев происходит не из инстинкта, а из адаптивного поведения – «консенсуса» – между особями внутри роя. Также предполагается, что роение является ритуалом , потому что самец мошки редко бывает сам по себе, а не в рое. Это могло образоваться из-за преимущества снижения инбридинга за счет того, что самцы с разными генами собираются в одном месте. [83] Род Culicoides , также известный как мокрецы, демонстрирует роевое поведение, которое, как полагают, вызывает замешательство у хищников. [84]
Тараканы оставляют химические следы в своих фекалиях, а также выделяют воздушные феромоны для спаривания. Другие тараканы будут следовать по этим следам, чтобы обнаружить источники пищи и воды, а также обнаружить, где прячутся другие тараканы. Таким образом, группы тараканов могут демонстрировать эмерджентное поведение , [85] в котором групповое или роевое поведение возникает из простого набора индивидуальных взаимодействий.
Тараканы в основном ведут ночной образ жизни и убегают, когда на них попадает свет. Исследование проверило гипотезу о том, что тараканы используют всего два вида информации, чтобы решить, куда идти в таких условиях: насколько темно и сколько там других тараканов. Исследование, проведенное Хосе Халлоем и его коллегами из Свободного университета Брюсселя и других европейских институтов, создало набор крошечных роботов , которые кажутся тараканам другими тараканами и, таким образом, могут изменить восприятие тараканами критической массы . Роботы также были специально ароматизированы, чтобы их приняли настоящие тараканы. [86]

Саранча — это роящаяся фаза короткорогих кузнечиков семейства Acrididae . Некоторые виды могут быстро размножаться при подходящих условиях и впоследствии становиться стайными и мигрирующими. Они образуют полосы в качестве нимф и стаи в качестве взрослых особей — и те, и другие могут перемещаться на большие расстояния, быстро опустошая поля и сильно повреждая посевы . Самые большие стаи могут покрывать сотни квадратных миль и содержать миллиарды саранчи. Саранча может съедать растения, равные своему собственному весу (около 2 граммов), каждый день. Это означает, что один миллион саранчи может съедать более одной тонны пищи каждый день, а самые большие стаи могут потреблять более 100 000 тонн каждый день. [87]
Было обнаружено, что роение у саранчи связано с повышенным уровнем серотонина , который заставляет саранчу менять цвет, есть гораздо больше, взаимно привлекаться и размножаться гораздо легче. Исследователи предполагают, что роевое поведение является ответом на перенаселение, и исследования показали, что повышенная тактильная стимуляция задних ног или, у некоторых видов, просто встреча с другими особями вызывает повышение уровня серотонина. Превращение саранчи в роящуюся разновидность может быть вызвано несколькими контактами в минуту в течение четырехчасового периода. [88] [89] [90] [91] Примечательно, что врожденная предрасположенность к агрегации была обнаружена у детенышей пустынной саранчи Schistocerca gregaria , независимо от их родительской фазы. [92]
Реакция отдельной саранчи на потерю выравнивания в группе, по-видимому, увеличивает хаотичность ее движения, пока снова не будет достигнуто выровненное состояние. Это вызванное шумом выравнивание, по-видимому, является внутренней характеристикой коллективного когерентного движения. [93]

Миграция насекомых — это сезонное перемещение насекомых , в частности , стрекоз , жуков , бабочек и моли . Расстояние может варьироваться от вида к виду, но в большинстве случаев эти перемещения охватывают большое количество особей. В некоторых случаях особи, мигрирующие в одном направлении, могут не вернуться, а следующее поколение может вместо этого мигрировать в противоположном направлении. Это существенное отличие от миграции птиц .
Бабочки-монархи особенно известны своей длительной ежегодной миграцией. В Северной Америке они совершают массовые миграции на юг, начиная с августа и до первых заморозков. Миграция на север происходит весной. Монарх — единственная бабочка, которая регулярно мигрирует как на север, так и на юг, как это делают птицы. Но ни одна особь не совершает полный круговой перелет. Самки-монархи откладывают яйца для следующего поколения во время этих миграций. [94] Продолжительность этих путешествий превышает обычную продолжительность жизни большинства монархов, которая составляет менее двух месяцев для бабочек, родившихся в начале лета. Последнее поколение лета вступает в нерепродуктивную фазу, известную как диапауза , и может жить семь месяцев или более. [95] Во время диапаузы бабочки перелетают в одно из многих мест зимовки. Поколение, которое зимует, обычно не размножается, пока не покинет место зимовки где-то в феврале и марте. Именно второе, третье и четвертое поколения возвращаются в свои северные места в Соединенных Штатах и Канаде весной. То, как этому виду удается возвращаться в те же самые места зимовки в течение нескольких поколений, все еще является предметом исследований; схемы полета, по-видимому, наследуются и основаны на сочетании положения солнца на небе [96] и компенсируемого по времени солнечного компаса, который зависит от циркадных часов , расположенных в их усиках. [97] [98]

 Бормотание скворцов
Бормотание скворцов[99]

Примерно 1800 из 10 000 видов птиц в мире являются дальними мигрантами. [100] Основной мотивацией миграции, по-видимому, является еда; например, некоторые колибри предпочитают не мигрировать, если их кормят зимой. Кроме того, более длинные дни северного лета предоставляют больше времени для размножения птиц, чтобы выкормить свое потомство. Это помогает дневным птицам производить более крупные кладки , чем родственные немигрирующие виды, которые остаются в тропиках. Поскольку дни осенью укорачиваются, птицы возвращаются в более теплые регионы, где доступный запас пищи мало меняется в зависимости от сезона. Эти преимущества компенсируют высокий стресс, затраты на физические нагрузки и другие риски миграции, такие как хищничество.
Многие птицы мигрируют стаями. Для более крупных птиц предполагается, что полет стаями снижает затраты энергии. V-образная формация часто должна повышать эффективность и дальность полета птиц, особенно на длинных миграционных маршрутах. Все птицы, за исключением первой, летят в восходящем потоке от одного из вихрей на концах крыльев птицы впереди. Восходящий поток помогает каждой птице поддерживать собственный вес в полете, так же как планер может подниматься или поддерживать высоту неограниченно долго в восходящем воздухе. Гуси, летящие в V-образной формации, экономят энергию, летя в восходящем потоке вихря на концах крыльев, созданном предыдущим животным в формации. Таким образом, птицам, летящим позади, не нужно прилагать столько усилий, чтобы достичь подъемной силы. Исследования показывают, что птицы в V-образной формации располагаются примерно на оптимальном расстоянии, предсказанном простой аэродинамической теорией. [101] Гуси в V-образной формации могут экономить 12–20% энергии, которая им потребовалась бы для полета в одиночку. [102] [103] В ходе радиолокационных исследований было обнаружено, что исландские песочники и чернозобики летают стаями на 5 км в час быстрее, чем когда они летят поодиночке. [104] Птицы, летящие на концах и впереди, меняются местами в циклическом режиме, чтобы равномерно распределить усталость от полета среди членов стаи. Такая формация также облегчает общение и позволяет птицам поддерживать визуальный контакт друг с другом.
Другие животные могут использовать похожие методы черчения во время миграции. Омары , например, мигрируют в тесном однорядном строю «лобстерные поезда», иногда на сотни миль.
Средиземное море и другие моря представляют собой серьезное препятствие для парящих птиц, которым приходится пересекать пролив в самых узких местах. Огромное количество крупных хищных птиц и аистов пролетает через такие районы, как Гибралтар , Фальстербо и Босфор во время миграции. Более распространенные виды, такие как европейский осоед , могут насчитывать сотни тысяч осенью. Другие препятствия, такие как горные хребты, также могут вызывать воронкообразное движение, особенно крупных дневных мигрантов. Это заметный фактор в центральноамериканском миграционном узком месте. Такая концентрация птиц во время миграции может поставить виды под угрозу. Некоторые впечатляющие мигранты уже вымерли, самым заметным из которых является странствующий голубь . Во время миграции стаи были шириной в милю (1,6 км) и длиной в 300 миль (500 км), на пролет уходило несколько дней, и насчитывалось до миллиарда птиц.

Термин «стая» можно использовать для описания любой группы рыб, включая смешанные группы, в то время как «стая» используется для более тесно сплоченных групп одного вида, плавающих в высокой степени синхронизированно и поляризованно.
Рыбы получают много преимуществ от поведения, связанного со стайным поведением, включая защиту от хищников (благодаря лучшему обнаружению хищников и снижению вероятности поимки), более успешную добычу пищи и более успешный поиск партнера. [106] Также вероятно, что рыбы получают выгоду от членства в стае за счет повышения гидродинамической эффективности. [107]
Рыбы используют множество признаков для выбора сородичей. Обычно они предпочитают более крупные косяки, сородичей своего вида, сородичей, похожих по размеру и внешнему виду на себя, здоровую рыбу и родственников (при распознавании). «Эффект странности» утверждает, что любой член стаи, который выделяется внешне, будет предпочтительно объектом нападения хищников. Это может объяснить, почему рыбы предпочитают собираться в стаи с особями, которые на них похожи. Таким образом, эффект странности будет иметь тенденцию гомогенизировать стаи. [108]
Один из загадочных аспектов выбора стаи заключается в том, как рыба может выбрать, присоединиться ли к стае животных, похожих на нее, учитывая, что она не может знать свою собственную внешность. Эксперименты с данио-рерио показали, что предпочтение стаи — это приобретенная способность, а не врожденная. Данио-рерио имеет тенденцию ассоциироваться со стаями, которые напоминают стаи, в которых она выросла, что является формой импринтинга . [109]
Другие открытые вопросы поведения стай включают определение того, какие особи отвечают за направление движения стай. В случае миграционного движения большинство членов стай, похоже, знают, куда они направляются. В случае поведения поиска пищи, стаи золотистых блестяшек (вид пескаря ) в неволе возглавляются небольшим числом опытных особей, которые знали, когда и где доступна еда. [110]
Радаков подсчитал, что косяки сельди в Северной Атлантике могут занимать площадь до 4,8 кубических километров (1,2 кубических миль) с плотностью рыбы от 0,5 до 1,0 рыбы/кубический метр, что в общей сложности составляет несколько миллиардов рыб в одном косяке. [111]
Между маем и июлем огромное количество сардин мечет икру в прохладных водах Агульяс-Бэнк , а затем следует за течением холодной воды на север вдоль восточного побережья Южной Африки. Эта большая миграция, называемая ходом сардин , создает впечатляющие пищевые безумства вдоль береговой линии, когда морские хищники, такие как дельфины, акулы и олуши, нападают на косяки.
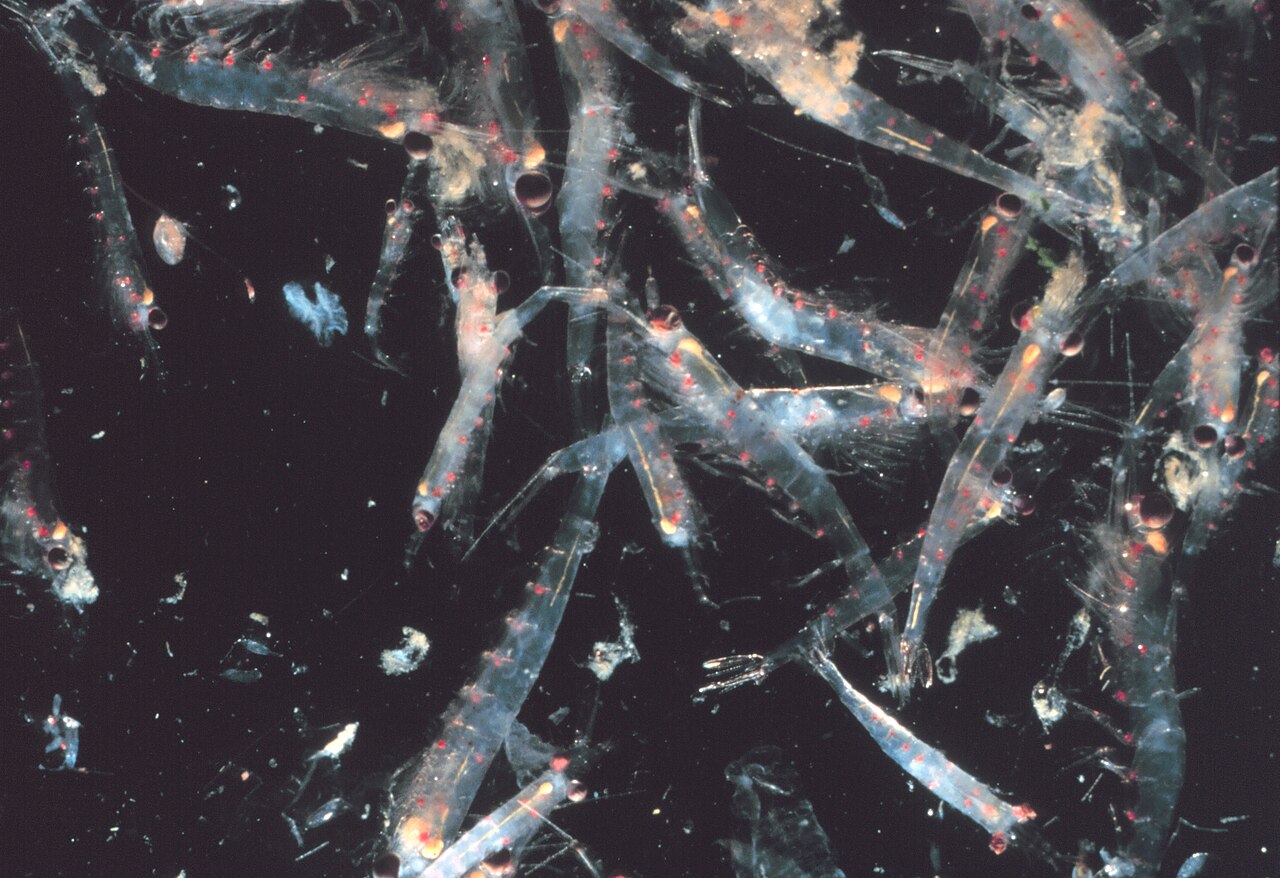
Большинство криля , мелких креветкообразных , образуют большие стаи, иногда достигающие плотности 10 000–60 000 особей на кубический метр. [113] [ 114] [115] Роение — это защитный механизм, сбивающий с толку мелких хищников, которые хотели бы выбрать отдельных особей. Самые большие стаи видны из космоса и могут отслеживаться со спутника. [116] Было замечено, что одна стая покрывала площадь в 450 квадратных километров (175 квадратных миль) океана на глубине 200 метров (650 футов) и, по оценкам, содержала более 2 миллионов тонн криля. [117] Недавние исследования показывают, что криль не просто пассивно дрейфует в этих течениях, но на самом деле изменяет их. [117] Криль обычно совершает суточную вертикальную миграцию . Двигаясь вертикально через океан в 12-часовом цикле, рои играют важную роль в смешивании более глубокой, богатой питательными веществами воды с бедной питательными веществами водой на поверхности. [117] До недавнего времени предполагалось, что они проводят день на большей глубине и поднимаются ночью к поверхности. Было обнаружено, что чем глубже они опускаются, тем больше они снижают свою активность, [118] по-видимому, чтобы уменьшить встречи с хищниками и сохранить энергию.
Более поздние исследования показали, что активность плавания криля меняется в зависимости от наполненности желудка. Насытившиеся животные, которые питались на поверхности, плавают менее активно и поэтому опускаются ниже смешанного слоя. [119] По мере того, как они опускаются, они производят фекалии, что может означать, что они играют важную роль в углеродном цикле Антарктики. Было обнаружено, что криль с пустыми желудками плавает более активно и, таким образом, направляется к поверхности. Это означает, что вертикальная миграция может быть явлением два или три раза в день. Некоторые виды образуют поверхностные стаи в течение дня для кормления и репродуктивных целей, хотя такое поведение опасно, поскольку оно делает их чрезвычайно уязвимыми для хищников. [120] Плотные стаи могут вызывать пищевое безумие среди рыб, птиц и млекопитающих хищников, особенно вблизи поверхности. Если их потревожить, рой рассеивается, и некоторые особи даже мгновенно линяют , оставляя экзувии в качестве приманки. [121] В 2012 году Гандоми и Алави представили то, что, по-видимому, является успешным стохастическим алгоритмом для моделирования поведения стай криля. Алгоритм основан на трех основных факторах: "(i) движение, вызванное присутствием других особей, (ii) деятельность по поиску пищи и (iii) случайная диффузия". [122]

Веслоногие рачки — это группа крошечных ракообразных, обитающих в море и озерах. Многие виды являются планктонными (дрейфующими в морских водах), а другие — бентосом (обитают на дне океана). Веслоногие рачки обычно имеют длину от 1 до 2 миллиметров (от 0,04 до 0,08 дюйма), каплевидное тело и большие антенны . Хотя, как и другие ракообразные, они имеют бронированный экзоскелет , они настолько малы, что у большинства видов эта тонкая броня и все тело почти полностью прозрачны. Веслоногие рачки имеют сложный, срединный одиночный глаз, обычно ярко-красный, в центре прозрачной головы.
Копеподы также роятся. Например, моноспецифические рои регулярно наблюдались вокруг коралловых рифов и морской травы , а также в озерах. Плотность роев составляла около миллиона копепод на кубический метр. Типичные рои были один или два метра в диаметре, но некоторые превышали 30 кубических метров. Копеподам нужен визуальный контакт, чтобы держаться вместе, и они рассеиваются ночью. [123]
Весной цветет роящийся фитопланктон , который обеспечивает пищей веслоногих рачков. Планктонные веслоногие рачки обычно являются доминирующими членами зоопланктона и , в свою очередь, являются основными пищевыми организмами для многих других морских животных. В частности, веслоногие рачки являются добычей кормовых рыб и медуз , которые могут собираться в огромные, миллионные стаи. Некоторые веслоногие рачки обладают чрезвычайно быстрой реакцией спасения , когда чувствуют хищника, и могут прыгать с высокой скоростью на несколько миллиметров (см. анимированное изображение ниже).

Планктонные веслоногие рачки важны для углеродного цикла . Некоторые ученые говорят, что они образуют самую большую биомассу животных на Земле. [124] Они конкурируют за этот титул с антарктическим крилем . Однако из-за своего меньшего размера и относительно более высоких темпов роста, а также из-за того, что они более равномерно распределены по большей части мирового океана, веслоногие рачки почти наверняка вносят гораздо больший вклад во вторичную продуктивность мирового океана и в глобальный поглотитель углерода в океане, чем криль , и, возможно, больше, чем все другие группы организмов вместе взятые. В настоящее время считается, что поверхностные слои океанов являются крупнейшим в мире поглотителем углерода, поглощая около 2 миллиардов тонн углерода в год, что эквивалентно, возможно, трети выбросов углерода человеком , тем самым уменьшая свое воздействие. Многие планктонные веслоногие рачки питаются вблизи поверхности ночью, а затем погружаются в более глубокие воды в течение дня, чтобы избежать визуальных хищников. Их линяющие экзоскелеты, фекальные гранулы и дыхание на глубине приносят углерод в глубокое море.
Многие одноклеточные организмы, называемые фитопланктоном, живут в океанах и озерах. При наличии определенных условий, таких как высокий уровень питательных веществ или света, эти организмы размножаются взрывообразно. Образующийся плотный рой фитопланктона называется цветением водорослей . Цветение может охватывать сотни квадратных километров и его легко увидеть на спутниковых снимках. Отдельные особи фитопланктона редко живут дольше нескольких дней, но цветение может длиться неделями. [125] [126]
Ученые приписывали роевое поведение растениям на протяжении сотен лет. В своей книге 1800 года « Фитология, или Философия сельского хозяйства и садоводства » Эразм Дарвин писал, что рост растений напоминает рои, наблюдаемые в других местах природы. [127] Хотя он ссылался на более общие наблюдения за морфологией растений и был сосредоточен как на поведении корней, так и побегов, недавние исследования подтвердили это утверждение.
Корни растений , в частности, демонстрируют наблюдаемое поведение роя, растущее по моделям, которые превышают статистический порог для случайной вероятности, и указывают на наличие связи между отдельными верхушками корней . Основная функция корней растений - поглощение питательных веществ из почвы , и именно эта цель движет поведением роя. Растения, растущие в непосредственной близости, адаптировали свой рост для обеспечения оптимальной доступности питательных веществ. Это достигается путем роста в направлении, которое оптимизирует расстояние между близлежащими корнями, тем самым увеличивая их шансы на использование неиспользованных запасов питательных веществ. Действие этого поведения принимает две формы: максимизация расстояния от соседних верхушек корней и отталкивание ими. [128] Переходная зона кончика корня в значительной степени отвечает за мониторинг наличия почвенных гормонов, сигнализируя об отзывчивых моделях роста по мере необходимости. Реакции растений часто являются сложными, интегрируя несколько входов для информирования об автономном ответе. Дополнительные входы, которые информируют о росте роя, включают свет и гравитацию, оба из которых также контролируются в переходной зоне верхушки корня. [129] Эти силы действуют, чтобы информировать любое количество растущих «главных» корней, которые демонстрируют свои собственные независимые выбросы ингибирующих химикатов для установления соответствующего интервала, тем самым способствуя модели поведения роя. Горизонтальный рост корней, будь то в ответ на высокое содержание минералов в почве или из-за роста столонов , производит разветвленный рост, который устанавливает также формирование своих собственных, независимых корневых роев. [130]
Роение также описывает группировки некоторых видов хищных бактерий, таких как миксобактерии . Миксобактерии роятся вместе в «волчьи стаи», активно перемещаясь, используя процесс, известный как бактериальное скольжение , и удерживаясь вместе с помощью межклеточных молекулярных сигналов . [57] [131]


Группа людей также может демонстрировать поведение роя, например, пешеходы [134] или солдаты, роящиеся на парапетах [ сомнительно – обсудить ] . В Кельне, Германия, два биолога из Университета Лидса продемонстрировали поведение, подобное поведению стаи, у людей. Группа людей демонстрировала похожую поведенческую модель, как у стаи, где если пять процентов стаи меняли направление, остальные следовали за ней. Если один человек был обозначен как хищник, а все остальные должны были избегать его, стая вела себя очень похоже на косяк рыб. [135] [136] Понимание того, как люди взаимодействуют в толпе, важно, если управление толпой должно эффективно избегать жертв на футбольных полях, музыкальных концертах и станциях метро. [137]
Математическое моделирование поведения стай является распространенной технологией и нашло применение в анимации. Моделирование стай использовалось во многих фильмах [138] для создания реалистично движущихся толп . «Бэтмен возвращается » Тима Бертона был первым фильмом, в котором использовалась технология роя для рендеринга, реалистично изображая движения группы летучих мышей с помощью системы boids . Трилогия фильмов «Властелин колец» использовала похожую технологию, известную как Massive , во время батальных сцен. Технология роя особенно привлекательна, поскольку она дешева, надежна и проста.
Компьютерное моделирование на основе муравьев, использующее всего шесть правил взаимодействия, также использовалось для оценки поведения при посадке в самолет. [139] Авиакомпании также использовали маршрутизацию на основе муравьев при назначении прибывающих самолетов на выходы в аэропорту. Система авиакомпании, разработанная Дугласом А. Лоусоном, использует теорию роя или роевой интеллект — идею о том, что колония муравьев работает лучше, чем один. Каждый пилот действует как муравей, ищущий лучшие выходы в аэропорту. «Пилот узнает на своем опыте, что лучше для него, и оказывается, что это лучшее решение для авиакомпании», — объясняет Лоусон. В результате «колония» пилотов всегда направляется к выходам, к которым они могут быстро прибыть и улететь. Программа может даже предупреждать пилота о резервных самолетах до того, как они произойдут. «Мы можем предвидеть, что это произойдет, поэтому у нас будут доступные выходы», — говорит Лоусон. [140]
Поведение роя также встречается в динамике транспортного потока , например, в транспортной волне . Двунаправленное движение можно наблюдать в муравьиных тропах. [141] [142] В последние годы это поведение исследовалось для понимания пешеходных и транспортных моделей. [143] [144] Моделирование, основанное на пешеходных моделях, также применялось к толпам, которые впадают в панику. [145]
Стадное поведение в маркетинге использовалось для объяснения зависимостей взаимного поведения клиентов. The Economist сообщил о недавней конференции в Риме на тему моделирования адаптивного поведения человека. [146] Он поделился механизмами для увеличения импульсивных покупок и побуждения людей «покупать больше, играя на стадном инстинкте». Основная идея заключается в том, что люди будут покупать больше продуктов, которые кажутся популярными, и упоминаются несколько механизмов обратной связи для предоставления потребителям информации о популярности продукта, включая технологию смарт-карт и использование технологии радиочастотной идентификации . Модель «движений роя» была представлена исследователем из Флоридского технологического института , которая привлекательна для супермаркетов, поскольку она может «увеличивать продажи без необходимости предоставлять людям скидки».

Применение принципов роя к роботам называется роевой робототехникой , в то время как роевой интеллект относится к более общему набору алгоритмов.
Частично вдохновленные колониями насекомых, таких как муравьи и пчелы, исследователи моделируют поведение роев из тысяч крошечных роботов, которые вместе выполняют полезную задачу, например, находят что-то спрятанное, убирают или шпионят. Каждый робот довольно прост, но возникающее поведение роя более сложно. [1] Весь набор роботов можно рассматривать как одну единую распределенную систему, так же как колонию муравьев можно считать суперорганизмом , демонстрирующим роевой интеллект . Самый большой созданный на сегодняшний день рой — это рой из 1024 роботов Kilobot. [148] Другие большие рои включают рой iRobot , проект SRI International /ActivMedia Robotics Centibots , [149] и рой Open-source Micro-robotic Project, которые используются для исследования коллективного поведения. [150] [151] Рои также более устойчивы к сбоям. В то время как один большой робот может выйти из строя и разрушить миссию, рой может продолжать работу, даже если несколько роботов выйдут из строя. Это может сделать их привлекательными для миссий по исследованию космоса, где неудача обычно обходится чрезвычайно дорого. [152] Помимо наземных транспортных средств, роевая робототехника также включает в себя исследование роев воздушных роботов [147] [153] и разнородных групп наземных и воздушных транспортных средств. [154] [155]
В отличие от макроскопических роботов, коллоидные частицы в микромасштабе также могут быть приняты в качестве агентов для выполнения коллективного поведения для выполнения задач с использованием механических и физических подходов, таких как реконфигурируемый торнадоподобный микророй [156], имитирующий стайную рыбу, [157] иерархические виды частиц [158], имитирующие хищное поведение млекопитающих, манипуляция микрообъектами с использованием трансформируемого микророя. [159] Изготовление таких коллоидных частиц обычно основано на химическом синтезе.

Военное роение — это поведение, при котором автономные или частично автономные единицы действия атакуют противника с нескольких разных направлений, а затем перегруппировываются. Пульсация , при которой единицы смещают точку атаки, также является частью военного роения. Военное роение подразумевает использование децентрализованной силы против противника таким образом, который подчеркивает мобильность, связь, автономность подразделений и координацию или синхронизацию. [160] Исторически военные силы использовали принципы роения, не исследуя их в действительности явно, но теперь активные исследования сознательно изучают военные доктрины, которые черпают идеи из роения.
Просто потому, что несколько подразделений сходятся на цели, они не обязательно роятся. Осадные операции не подразумевают роения, потому что нет маневра; есть схождение, но на осажденном укреплении. Партизанские засады также не являются роями, потому что они "бей и беги". Даже если засада может иметь несколько точек атаки на противника, партизаны отступают, когда они либо нанесли достаточный урон, либо когда они находятся в опасности.
В 2014 году Управление военно-морских исследований США опубликовало видео, демонстрирующее испытания роя небольших автономных ударных катеров-беспилотников, которые могут управлять и предпринимать скоординированные наступательные действия как группа. [161]

![Люди, толпящиеся у выхода, не всегда ведут себя как жидкость.[163][164]](http://upload.wikimedia.org/wikipedia/commons/thumb/5/54/Crowd_04378.JPG/1280px-Crowd_04378.JPG)


{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite web}}: CS1 maint: неподходящий URL ( ссылка )


![Сальпы, расположенные в цепочках, образуют огромные рои.[162]](http://upload.wikimedia.org/wikipedia/commons/thumb/6/63/Salp.jpg/1280px-Salp.jpg)

