Эхолокация , также называемая биосонаром , представляет собой биологически активный сонар , используемый несколькими группами животных как в воздухе, так и под водой. Эхолокирующие животные издают крики и прислушиваются к эху этих криков, которое возвращается от различных объектов рядом с ними. Они используют это эхо, чтобы найти и идентифицировать объекты. Эхолокация используется для навигации , поиска пищи и охоты на добычу .
Эхолокационные вызовы могут быть частотно-модулированными (FM, высота звука меняется во время разговора) или постоянной частотой (CF). FM предлагает точную дискриминацию по дальности для локализации добычи за счет уменьшения дальности действия. CF позволяет определять как скорость добычи, так и ее движения с помощью эффекта Доплера . FM может быть лучше всего подходит для тесных и загроможденных помещений, тогда как CF может быть лучше на открытом воздухе или для охоты, сидя на месте.
К эхолокационным животным относятся млекопитающие, особенно зубатые киты (зубистые киты) и некоторые виды летучих мышей , а также, если использовать более простые формы, виды других групп, например землероек . Несколько видов птиц из двух пещерных групп птиц используют эхолокацию, а именно пещерные стрижи и масляные птицы .
Некоторые животные-жертвы, на которых охотятся летучие мыши с эхолокацией, принимают активные контрмеры , чтобы избежать поимки. К ним относятся уклонение от хищников, отражение атаки и использование ультразвуковых щелчков, которые приобрели множество функций, включая апосематизм, мимику химически защищенных видов и эхолокационное подавление.
Термин эхолокация был придуман в 1944 году американским зоологом Дональдом Гриффином , который вместе с Робертом Галамбосом впервые продемонстрировал это явление у летучих мышей. [1] [2] Как описал Гриффин в своей книге, [3] итальянский учёный 18-го века Лаццаро Спалланцани посредством серии тщательно продуманных экспериментов пришел к выводу, что, когда летучие мыши летают ночью, они полагаются не только на зрение, но и на какие-то чувства, но он не обнаружил, что другое чувство — это слух. [4] [5] Швейцарский врач и натуралист Луи Жюрин повторил эксперименты Спалланцани (с использованием разных видов летучих мышей) и пришел к выводу, что, когда летучие мыши охотятся ночью, они полагаются на слух. [6] [7] [8] В 1908 году Уолтер Луи Хан подтвердил выводы Спалланцани и Юрина. [9]
В 1912 году изобретатель Хирам Максим независимо предположил, что летучие мыши используют звуки ниже человеческого слухового диапазона , чтобы избегать препятствий. [10] В 1920 году английский физиолог Гамильтон Хартридж правильно предположил, что летучие мыши используют частоты, превышающие диапазон человеческого слуха . [11] [12]
Эхолокация у зубатых китов (зубых китов) не была должным образом описана Шевиллом и Макбрайдом лишь через два десятилетия после работы Гриффина и Галамбоса в 1956 году . [13] Однако в 1953 году Жак Ив Кусто в своей первой книге «Безмолвный мир» предположил , что у морских свиней было что-то вроде гидролокатора , судя по их навигационным способностям. [14]
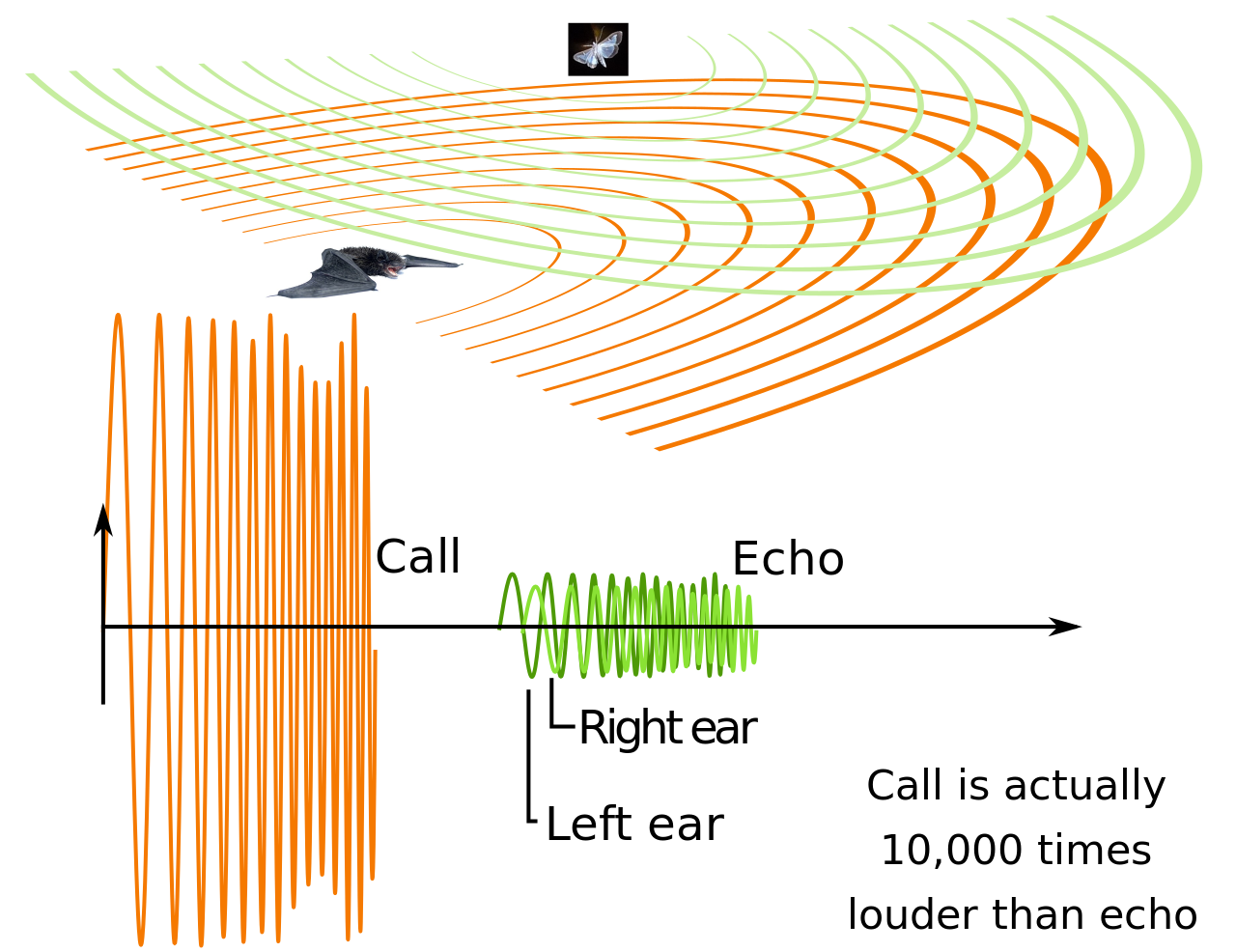
Эхолокация – это активный гидролокатор , использующий звуки, издаваемые самим животным. Определение дальности достигается путем измерения временной задержки между собственным звуковым излучением животного и любым эхом, возвращающимся из окружающей среды. Относительная интенсивность звука, принимаемого каждым ухом, а также временная задержка между приходом в два уха предоставляют информацию о горизонтальном угле (азимуте), под которым приходят отраженные звуковые волны. [15]
В отличие от некоторых гидролокаторов, созданных человеком, которые используют множество чрезвычайно узких лучей и множество приемников для локализации цели ( многолучевой гидролокатор ), эхолокация животных имеет только один передатчик и два приемника (уши), расположенные немного друг от друга. Эхо, возвращающееся в уши, приходит в разное время и с разной интенсивностью, в зависимости от положения объекта, генерирующего эхо. Разница во времени и громкости используется животными для восприятия расстояния и направления. С помощью эхолокации летучая мышь или другое животное может определить не только то, куда оно движется, но также насколько велико другое животное, что это за животное и другие особенности. [16] [17]
Описание разнообразия эхолокационных сигналов требует изучения частоты и временных особенностей звуков. Именно различия в этих аспектах вызывают эхолокационные сигналы, подходящие для различных акустических сред и охотничьего поведения. Наиболее интенсивно исследовались крики летучих мышей, но эти принципы применимы ко всем эхолокационным вызовам. [18] [19]
Частоты криков летучих мышей варьируются от 11 кГц до 212 кГц. [20] Насекомоядные летучие мыши-ястребы, преследующие добычу на открытом воздухе, имеют частоту крика от 20 до 60 кГц, поскольку именно эта частота обеспечивает наилучшую дальность и четкость изображения и делает их менее заметными для насекомых. [21] Однако низкие частоты являются адаптивными для некоторых видов с различной добычей и окружающей средой. Euderma maculatum , вид летучих мышей, питающийся мотыльками , использует особенно низкую частоту 12,7 кГц, которую мотыльки не слышат. [22]
Эхолокационные вызовы могут состоять из двух различных типов частотной структуры: частотно-модулированных (FM) разверток и тонов постоянной частоты (CF). Конкретный вызов может состоять из одной, другой или обеих структур. FM-развертка представляет собой широкополосный сигнал, то есть содержит нисходящую развертку по диапазону частот. Тон CF представляет собой узкополосный сигнал: звук остается постоянной на одной частоте на протяжении всего своего существования. [23]
Эхолокационные сигналы у летучих мышей измерялись с интенсивностью от 60 до 140 децибел . [24] Некоторые виды летучих мышей могут изменять интенсивность своего крика в середине разговора, снижая интенсивность по мере приближения к объектам, которые сильно отражают звук. Это не позволяет возвращающемуся эху оглушить летучую мышь. [19] Высокоинтенсивные крики, например, издаваемые летучими мышами (133 дБ), адаптируются к охоте под открытым небом. Их звуки высокой интенсивности необходимы даже для умеренного обнаружения окружающей обстановки, поскольку воздух обладает высоким поглощением ультразвука, а размер насекомых обеспечивает лишь небольшую цель для отражения звука. [25] Кроме того, так называемые «шепчущие летучие мыши» адаптировали низкоамплитудную эхолокацию, так что их добыча, мотыльки, способные слышать эхолокационные сигналы, менее способны обнаруживать и избегать приближающихся летучих мышей. [22] [26]
Одиночный эхолокационный вызов (вызов представляет собой один непрерывный след на звуковой спектрограмме и серию вызовов, состоящих из последовательности или прохода) может длиться от менее 3 до более 50 миллисекунд. Длительность импульса составляет около 3 миллисекунд у летучих мышей FM, таких как Phyllostomidae и некоторых Vespertilionidae; от 7 до 16 миллисекунд у летучих мышей с квазипостоянной частотой (QCF), таких как другие Vespertilionidae, Emballonuridae и Molossidae; и от 11 миллисекунд (Hipposideridae) до 52 миллисекунд (Rhinolophidae) у летучих мышей CF. [27] Продолжительность зависит также от стадии поведения по ловле добычи, на которой находится летучая мышь, обычно уменьшается, когда летучая мышь находится на заключительной стадии захвата добычи - это позволяет летучей мыши кричать быстрее, без дублирования крика и эха. Уменьшение продолжительности происходит за счет уменьшения общего количества звука, который может отражаться от объектов и быть услышан летучей мышью. [20]
Временной интервал между последующими эхолокационными вызовами (или импульсами) определяет два аспекта восприятия летучей мыши. Во-первых, он определяет, насколько быстро обновляется информация о слуховой сцене летучей мыши. Например, летучие мыши увеличивают частоту повторения своих криков (то есть уменьшают интервал пульса), когда приближаются к цели. Это позволяет летучей мыши быстрее получать новую информацию о местоположении цели, когда она ей больше всего нужна. Во-вторых, интервал импульсов определяет максимальную дальность, на которой летучие мыши могут обнаруживать объекты. Это связано с тем, что летучие мыши могут отслеживать эхо только одного звонка за раз; как только они делают еще один звонок, они перестают прислушиваться к эху от предыдущего звонка. Например, интервал импульса 100 мс (типичный для летучей мыши, ищущей насекомых) позволяет звуку распространяться в воздухе примерно на 34 метра, поэтому летучая мышь может обнаруживать объекты только на расстоянии до 17 метров (звук должен распространяться вперед и назад). . При интервале импульсов 5 мс (типично для летучей мыши в последние моменты попытки поимки) летучая мышь может обнаруживать объекты только на расстоянии до 85 см. Поэтому летучей мыши постоянно приходится делать выбор между быстрым обновлением новой информации и обнаружением объектов на большом расстоянии. [28]
Основным преимуществом FM-сигнала является чрезвычайно точное распознавание дальности или локализация цели. Дж. А. Симмонс продемонстрировал этот эффект с помощью серии экспериментов, которые показали, как летучие мыши, использующие FM-сигналы, могут различать две отдельные цели, даже если цели находились на расстоянии менее полмиллиметра друг от друга. Эта способность обусловлена широкополосной разверткой сигнала, которая позволяет лучше разрешать временную задержку между вызовом и возвращающимся эхом, тем самым улучшая взаимную корреляцию между ними. Если к ЧМ-сигналу добавить частоты гармоник, то эта локализация станет еще более точной. [29] [30] [31]
Одним из возможных недостатков FM-сигнала является уменьшение радиуса действия вызова. Поскольку энергия вызова распределена по многим частотам, расстояние, на котором FM-летучая мышь может обнаруживать цели, ограничено. [32] Частично это связано с тем, что любое эхо, возвращающееся на определенной частоте, можно оценить только в течение короткой доли миллисекунды, поскольку быстрое нисходящее колебание вызова не остается на какой-либо одной частоте надолго. [30]
Структура сигнала CF является адаптивной, поскольку она позволяет летучей мыши CF обнаруживать как скорость цели, так и трепетание крыльев цели при доплеровском сдвиге частот. Доплеровский сдвиг — это изменение частоты звуковой волны, которое возникает в двух важных ситуациях: когда летучая мышь и ее цель движутся относительно друг друга и когда крылья цели колеблются вперед и назад. CF-летучие мыши должны компенсировать доплеровские сдвиги, понижая частоту своего крика в ответ на эхо повышенной частоты – это гарантирует, что возвращающееся эхо останется на той частоте, на которую уши летучей мыши настроены наиболее точно. Колебания крыльев цели также вызывают сдвиги амплитуды, что дает летучей мыши CF дополнительную помощь в различении летящей цели от неподвижной. [33] [29] Подковоносы охотятся таким образом. [34]
Кроме того, поскольку энергия сигнала вызова CF сосредоточена в узкой полосе частот, рабочий диапазон вызова намного больше, чем у FM-сигнала. Это основано на том факте, что эхо-сигналы, возвращающиеся в узком диапазоне частот, можно суммировать по всей длине вызова, что обеспечивает постоянство частоты до 100 миллисекунд. [30] [32]
FM-компонент отлично подходит для охоты на добычу во время полета в тесноте и загроможденном пространстве. Этот факт объясняется двумя аспектами FM-сигнала: точной локализацией цели, обеспечиваемой широкополосным сигналом, и короткой продолжительностью разговора. Первое из них важно, потому что в загроможденной среде летучие мыши должны быть в состоянии распознать свою добычу по большому количеству фонового шума. Способность широкополосного сигнала к трехмерной локализации позволяет летучей мыши делать именно это, предоставляя ей то, что Симмонс и Штейн (1980) называют «стратегией отклонения помех». [31] Эта стратегия дополнительно улучшается за счет использования гармоник, которые, как уже говорилось ранее, улучшают свойства локализации вызова. Короткая продолжительность FM-вызова также лучше всего подходит для тесных и загроможденных помещений, поскольку позволяет летучей мыши очень быстро передавать множество вызовов без перекрытия. Это означает, что летучая мышь может получать почти непрерывный поток информации (что важно, когда объекты находятся близко, потому что они быстро проходят мимо), не путая, какое эхо соответствует какому вызову. [33] [29]
Компонент CF часто используется летучими мышами, охотящимися за добычей во время полета на открытой, свободной от помех среде, или летучими мышами, которые ждут на насестах появления своей добычи. Успех первой стратегии обусловлен двумя аспектами призыва CF, оба из которых обеспечивают превосходные способности к обнаружению добычи. Во-первых, больший рабочий диапазон сигнала позволяет летучим мышам обнаруживать цели, находящиеся на больших расстояниях – обычная ситуация на открытой местности. Во-вторых, длина звонка также подходит для целей, находящихся на больших расстояниях: в этом случае снижается вероятность того, что длинный звонок перекроется с возвращающимся эхом. Последняя стратегия стала возможной благодаря тому факту, что длинный узкополосный сигнал позволяет летучей мыши обнаружить доплеровские сдвиги, которые могут быть вызваны насекомым, движущимся либо к сидящей летучей мыши, либо от нее. [33] [31] [29]
Эхолокация встречается у множества млекопитающих и птиц, как описано ниже. [35] Он неоднократно развивался, что является примером конвергентной эволюции . [29] [36]

Эхолокирующие летучие мыши используют эхолокацию для навигации и поиска пищи, часто в полной темноте. Обычно они выходят из своих насестов в пещерах, на чердаках или на деревьях в сумерках и охотятся за насекомыми до поздней ночи. Используя эхолокацию, летучие мыши могут определить, насколько далеко находится объект, его размер, форму и плотность, а также направление (если есть), в котором объект движется. Использование ими эхолокации, наряду с активным полетом, позволяет им занимать нишу, где часто бывает много насекомых (которые выходят ночью, поскольку тогда меньше хищников), меньше конкуренции за пищу и меньше видов, которые могут охотиться на летучих мышей. сами себя. [37]
Эхолокирующие летучие мыши генерируют ультразвук через гортань и издают звук через открытый рот или, что гораздо реже, через нос. [38] Последнее наиболее выражено у подковоносов ( Rhinolophus spp. ). Частота эхолокационных сигналов летучих мышей варьируется от 14 000 до более 100 000 Гц, в основном за пределами диапазона человеческого уха (обычный диапазон человеческого слуха считается от 20 Гц до 20 000 Гц). Летучие мыши могут оценить высоту цели, интерпретируя интерференционные картины, вызванные эхом, отражающимся от козелка , лоскута кожи наружного уха. [39]
Отдельные виды летучих мышей используют эхолокацию в определенных диапазонах частот, которые соответствуют их среде обитания и типам добычи. Иногда исследователи использовали это для идентификации летучих мышей, летающих в определенной местности, просто записывая их крики с помощью ультразвуковых записывающих устройств, известных как «детекторы летучих мышей». Однако эхолокационные сигналы не всегда специфичны для вида, и у некоторых летучих мышей типы используемых ими криков совпадают, поэтому записи эхолокационных сигналов не могут использоваться для идентификации всех летучих мышей. Исследователи в нескольких странах разработали «библиотеки криков летучих мышей», которые содержат записи «справочных криков» местных видов летучих мышей для облегчения идентификации. [40] [41] [42]
При поиске добычи они издают звуки с низкой частотой (10–20 щелканий в секунду). Во время фазы поиска звуковое излучение сочетается с дыханием, которое снова связано с взмахами крыльев. Эта связь, по-видимому, значительно экономит энергию, поскольку летучим мышам практически не требуются дополнительные энергетические затраты на эхолокацию. [43] После обнаружения потенциальной добычи летучие мыши-эхолокаторы увеличивают частоту импульсов, заканчивающуюся гудением терминала , со скоростью до 200 щелчков в секунду. По мере приближения к обнаруженной цели длительность звуков постепенно уменьшается, как и энергия звука. [44]
Летучие мыши появились в начале эпохи эоцена , около 64 млн лет назад . Yangochiroptera появились примерно 55 млн лет назад, а Rhinolophoidea — примерно 52 млн лет назад. [45]
Существуют две гипотезы об эволюции эхолокации у летучих мышей. Первое предполагает, что эхолокация гортани развивалась дважды или более у рукокрылых, по крайней мере один раз у Yangochiroptera и по крайней мере один раз у подковоносов (Rhinolophidae): [46]
Второе предполагает, что гортанная эхолокация имела единственное происхождение у рукокрылых, т. е. была базальной по отношению к группе и впоследствии была утеряна в семействе Pteropodidae . [47] Позже род Rousettus семейства Pteropodidae развил другой механизм эхолокации с использованием системы щелкания языком: [48]
Эхолокирующие летучие мыши обитают в самых разных экологических условиях; их можно встретить живущими в таких разных средах, как Европа и Мадагаскар , и охотящимися за такими разными источниками пищи, как насекомые, лягушки, нектар, фрукты и кровь. Характеристики эхолокационного сигнала адаптированы к конкретной окружающей среде, охотничьему поведению и источнику пищи конкретной летучей мыши. Адаптация эхолокационных сигналов к экологическим факторам ограничена филогенетическими отношениями летучих мышей, что приводит к процессу, известному как «происхождение с модификациями», и приводит к сегодняшнему разнообразию рукокрылых. [29] [32] [31] Летучие мыши могут случайно заглушить друг друга, а в некоторых ситуациях они могут перестать кричать, чтобы избежать помех. [49]
Летающие насекомые являются распространенным источником пищи для летучих мышей, владеющих эхолокацией, а некоторые насекомые (в частности, мотыльки) могут слышать крики хищных летучих мышей. Есть свидетельства того, что слух бабочек развился в ответ на эхолокацию летучих мышей, позволяющую избежать поимки. [50] Более того, эти адаптации бабочек оказывают селективное давление на летучих мышей, заставляя их совершенствовать свои системы охоты на насекомых, и этот цикл завершается « эволюционной гонкой вооружений » между бабочками и летучими мышами . [51] [52]
Поскольку летучие мыши используют эхолокацию, чтобы ориентироваться и находить объекты, их слуховые системы приспособлены для этой цели и высокоспециализированы для восприятия и интерпретации стереотипных эхолокационных сигналов, характерных для их собственного вида. Эта специализация очевидна от внутреннего уха до самых высоких уровней обработки информации в слуховой коре. [53]
Летучие мыши CF и FM имеют специализированное внутреннее ухо, которое позволяет им слышать звуки в ультразвуковом диапазоне, далеко выходящие за пределы диапазона человеческого слуха. Хотя в большинстве других аспектов слуховые органы летучих мышей аналогичны органам слуха большинства других млекопитающих, некоторые летучие мыши ( подковоносы , Rhinolophus spp. и усатые летучие мыши , Pteronotus parnelii ) имеют компонент постоянной частоты (CF) в их крике (известный как летучие мыши с высоким рабочим циклом) имеют несколько дополнительных приспособлений для обнаружения преобладающей частоты (и гармоник) вокализации CF. К ним относится узкая частотная «настройка» органов внутреннего уха с особенно большой площадью, реагирующей на частоту возвращающегося эха летучей мыши. [33]
Базилярная мембрана внутри улитки содержит первую из этих специализаций для обработки эхоинформации. У летучих мышей, использующих сигналы CF, участок мембраны, реагирующий на частоту возвращающегося эха, намного больше, чем область ответа на любую другую частоту. Например, у большого подковоноса Rhinolophusferrumequinum имеется непропорционально удлиненный и утолщенный участок мембраны, который реагирует на звуки с частотой около 83 кГц — постоянной частоты эха, производимого криком летучей мыши. Эта область высокой чувствительности к определенному узкому диапазону частот известна как « акустическая ямка ». [54]
У эхолокирующих летучих мышей есть волоски улитки, которые особенно устойчивы к интенсивному шуму. Волосковые клетки улитки необходимы для слуховой чувствительности и могут быть повреждены сильным шумом. Поскольку летучие мыши регулярно подвергаются интенсивному шуму посредством эхолокации, необходима устойчивость к деградации из-за интенсивного шума. [55]
Далее по слуховому пути движение базилярной мембраны приводит к стимуляции первичных слуховых нейронов. Многие из этих нейронов специально «настроены» (наиболее сильно реагируют) на узкий диапазон частот возвращающихся эхо-сигналов сигналов МВ. Из-за большого размера слуховой ямки количество нейронов, реагирующих на эту область и, следовательно, на частоту эха, особенно велико. [56]
В нижних холмиках , структуре среднего мозга летучей мыши, информация из нижних отделов слухового пути интегрируется и отправляется в слуховую кору. Как показали Джордж Поллак и другие в серии работ в 1977 году, интернейроны в этой области обладают очень высоким уровнем чувствительности к разнице во времени, поскольку временная задержка между звонком и возвращающимся эхом сообщает летучей мыши о ее расстоянии от целевого объекта. . Хотя большинство нейронов быстрее реагируют на более сильные стимулы, колликулярные нейроны сохраняют точность синхронизации даже при изменении интенсивности сигнала. [57] Эти интернейроны специализируются на чувствительности ко времени несколькими способами. Во-первых, при активации они обычно реагируют только одним или двумя потенциалами действия . Эта короткая продолжительность реакции позволяет их потенциалам действия точно указать момент прибытия стимула и точно реагировать на стимулы, возникающие близко друг к другу по времени. У нейронов очень низкий порог активации — они быстро реагируют даже на слабые раздражители. Наконец, для FM-сигналов каждый интернейрон настроен на определенную частоту в пределах развертки, а также на ту же самую частоту в последующем эхо. На этом уровне также существует специализация для компонента вызова CF. На этом уровне фактически увеличивается доля нейронов, реагирующих на частоту слуховой ямки. [57]
Слуховая кора летучих мышей довольно велика по сравнению с другими млекопитающими. [58] Различные характеристики звука обрабатываются разными областями коры головного мозга, каждый из которых предоставляет различную информацию о местоположении или движении целевого объекта. Большинство существующих исследований по обработке информации в слуховой коре летучей мыши было проведено Нобуо Сугой на усатой летучей мыши Pteronotus parnellii . Звук этой летучей мыши включает в себя как тон CF, так и компоненты развертки FM. [59] [60]
Шуга и его коллеги показали, что кора головного мозга содержит ряд «карт» слуховой информации, каждая из которых систематически организована на основе таких характеристик звука, как частота и амплитуда . Нейроны в этих областях реагируют только на определенную комбинацию частоты и времени (задержка звукового эха) и известны как нейроны, чувствительные к комбинации. [59] [60]
Систематически организованные карты слуховой коры реагируют на различные аспекты эхо-сигнала, такие как его задержка и скорость. Эти области состоят из «комбинационно-чувствительных» нейронов, которым требуется как минимум два конкретных стимула, чтобы вызвать ответ. Нейроны систематически различаются по картам, которые организованы по акустическим особенностям звука и могут быть двумерными. Различные особенности крика и его эха используются летучей мышью для определения важных характеристик своей добычи. Карты включают: [59] [60]


Биосонары ценны как для зубатых китов (подотряд Odontoceti ), включая дельфинов , морских свиней , речных дельфинов , косаток и кашалотов , так и для усатых китов (подотряд Mysticeti ), включая правых , гренландских , карликовых правых , а также серых китов и рорквалов , поскольку они живут в подводной среде обитания, которая имеет благоприятные акустические характеристики и где дальность обзора часто крайне ограничена из-за поглощения или помутнения . [62] Зубатые киты обычно способны слышать звуки на ультразвуковых частотах, тогда как китообразные слышат звуки в инфразвуковом частотном режиме. [63]
Эволюция китообразных состояла из трех основных излучений . На протяжении всего периода среднего и позднего эоцена (49-31,5 млн лет назад) единственными китообразными были археоцеты — примитивные зубатые китообразные, произошедшие от наземных млекопитающих. [64] [65] Они не пользовались эхолокацией, но слегка адаптировали подводный слух. [66] К концу среднего эоцена акустически изолированные ушные кости развились, чтобы дать базилозавридным археоцетам направленный подводный слух на низких и средних частотах. [67] С вымиранием археоцетов в начале олигоцена ( 33,9–23 миллиона лет назад) во второй радиации возникли две новые линии. Ранние усатые киты и зубатые киты появились в среднем олигоцене в Новой Зеландии. [65] Существующие зубатые киты являются монофилетическими (единая эволюционная группа), но эхолокация развивалась дважды, конвергентно: один раз у Xenorophus , олигоценовых стволовых зубатых китов, и один раз у коронковых зубатых китов. [36]
Физическая реструктуризация океанов сыграла роль в эволюции эхолокации. Глобальное похолодание на границе эоцена и олигоцена привело к превращению мира из оранжереи в ледник . Тектонические разломы создали Южный океан со свободным течением Антарктического циркумполярного течения . [66] [67] [68] Эти события стимулировали отбор на способность находить и ловить добычу в мутных речных водах, что позволило зубатым китообразным вторгаться и питаться на глубинах ниже фототической зоны . В частности, эхолокация ниже фотической зоны могла быть адаптацией хищников к мигрирующим головоногим моллюскам . [67] [69] Семейство Delphinidae (дельфины) диверсифицировалось в неогене (23–2,6 миллиона лет назад), развив чрезвычайно специализированную эхолокацию. [70] [66]
Четыре белка играют важную роль в эхолокации зубатых китов. Престин , моторный белок наружных волосковых клеток внутреннего уха улитки млекопитающих , связан со слуховой чувствительностью. [71] Он претерпел два явных эпизода ускоренной эволюции у китообразных. [71] Первое связано с дивергенцией зубатых китов, когда впервые развилась эхолокация, а второе — с увеличением частоты эхолокации среди дельфинов. Tmc1 и Pjvk — это белки, связанные со слуховой чувствительностью: Tmc1 связан с развитием волосковых клеток и высокочастотным слухом, а Pjvk — с функцией волосковых клеток. [72] Молекулярная эволюция Tmc1 и Pjvk указывает на положительный отбор эхолокации у зубатых китов. [72] Cldn14 , член белков плотного соединения, которые образуют барьеры между клетками внутреннего уха, демонстрирует ту же эволюционную картину, что и Престин. [73] Два события эволюции белка, для Престина и Cldn14, произошли в то же время, что и тектоническое открытие пролива Дрейка (34–31 млн лет назад) и рост антарктического льда при переходном климате среднего миоцена (14 млн лет назад), с дивергенция зубатых и усатых китов происходит с первыми, а видообразование Delphinidae — со вторыми. [68]
Эволюция двух краниальных структур может быть связана с эхолокацией. Краниальное телескопирование (перекрытие лобных и верхнечелюстных костей и смещение ноздрей назад [74] ) впервые развилось у ксенорофидов . Далее он развился у стеблевых зубатых китов, достигнув полного телескопирования черепа у корончатых зубатых китов. [75] Движение ноздрей, возможно, позволило использовать носовой аппарат большего размера и дыню для эхолокации. [75] Это изменение произошло после расхождения неокетов от базилозавридов. [76] Первый сдвиг в сторону краниальной асимметрии произошел в раннем олигоцене, до появления ксенофидов. [76] Окаменелость ксенорофида ( Cotylocara macei ) имеет черепную асимметрию и демонстрирует другие показатели эхолокации. [77] Однако базальные ксенорофиды лишены краниальной асимметрии, что указывает на то, что она, вероятно, эволюционировала дважды. [76] У современных зубатых китов асимметричные носолицевые области; как правило, срединная плоскость смещена влево, а структуры справа крупнее. [77] И краниальное телескопирование, и асимметрия, вероятно, связаны с производством звука для эхолокации. [75]
Тринадцать видов современных зубатых китов конвергентно развили узкополосную высокочастотную эхолокацию (NBHF) в четырех отдельных событиях. К этим видам относятся семейства Kogiidae (карликовые кашалоты) и Phocoenidae (морские свиньи), а также некоторые виды рода Lagenorhynchus , все Cephalorhynchus и дельфин Ла-Платы . Считается, что NBHF развился как средство уклонения от хищников; Виды, продуцирующие NBHF, имеют небольшие размеры по сравнению с другими зубатыми китами, что делает их жизнеспособной добычей для крупных видов, таких как косатка . Однако, поскольку три группы развили NBHF до появления косаток, движущей силой развития NBHF должно было быть хищничество других древних хищных зубатых китов, а не хищничество со стороны косаток. Косатки и, предположительно, древние хищные зубатые киты, такие как Acrophyseter , не способны слышать частоты выше 100 кГц. [78]
Еще одна причина изменчивости эхолокации — среда обитания. Для всех гидроакустических систем ограничивающим фактором, определяющим, будет ли обнаружено возвращающееся эхо, является отношение эха к шуму (ENR). ENR определяется уровнем излучаемого источника (SL) плюс целевая мощность минус двусторонние потери при передаче (поглощение и распространение) и принимаемый шум. [79] Животные адаптируются либо для увеличения дальности действия в условиях ограниченного шума (увеличение уровня источника), либо для уменьшения шумового шума в мелкой и/или замусоренной среде обитания (уменьшение уровня источника). В загроможденных местах обитания, таких как прибрежные районы, ареалы добычи меньше, а такие виды, как дельфин Коммерсона ( Cephalorhynchus commersonii ), снизили уровни источника, чтобы лучше соответствовать окружающей среде. [79]
Зубатые киты издают сфокусированный луч высокочастотных щелчков в направлении, куда смотрит их голова. Звуки генерируются путем прохождения воздуха из костных ноздрей через слуховые губы . Эти звуки отражаются плотной вогнутой костью черепа и воздушным мешком в ее основании. Сфокусированный луч модулируется большим жировым органом, известным как дыня. Он действует как акустическая линза, поскольку состоит из липидов разной плотности. Большинство зубатых китов используют серии щелчков или серию щелчков для эхолокации, в то время как кашалоты могут производить щелчки индивидуально. Свистки зубатых китов, похоже, не используются для эхолокации. Различная скорость производства щелчков в поезде щелчков вызывает знакомый лай, визг и рычание афалины . Последовательность щелчков с частотой повторения более 600 в секунду называется пакетным импульсом. У афалин слуховая реакция мозга разрешает отдельные щелчки до 600 в секунду, но дает дифференцированную реакцию при более высокой частоте повторения. [80]
Было высказано предположение, что расположение зубов некоторых более мелких зубатых китов может быть адаптацией к эхолокации. [81] Зубы афалины, например, расположены несимметрично, если смотреть с вертикальной плоскости. Эта асимметрия, возможно, поможет определить, исходят ли эхо от биосонара с одной или другой стороны; но это не проверено экспериментально. [82]
Эхо принимается с использованием сложных жировых структур вокруг нижней челюсти в качестве основного пути приема, откуда они передаются в среднее ухо через сплошное жировое тело. Боковой звук может восприниматься через жировые доли, окружающие уши, плотность которых аналогична плотности воды. Некоторые исследователи полагают, что, приближаясь к интересующему объекту, они защищаются от более громкого эха, заглушая издаваемый звук. Известно, что такое случается и у летучих мышей, но и здесь слуховая чувствительность вблизи цели снижается. [83] [84]
_hunting_by_echolocation.JPG/1280px-Palawan_swiftlet_(Aerodramus_palawanensis)_hunting_by_echolocation.JPG)
Известно, что масличные птицы и некоторые виды стрижлетов используют относительно грубую форму эхолокации по сравнению с таковой у летучих мышей и дельфинов. Эти ночные птицы издают крики во время полета и используют их, чтобы перемещаться по деревьям и пещерам, где они живут. [85] [86]
Наземные млекопитающие, кроме летучих мышей, которые, как известно или предположительно, обладают эхолокацией, включают землероек , [87] [88] [89] тенреков Мадагаскара , [ 90 ] китайских карликовых соней , [ 91 ] и соленодонов . [92] Звуки землеройки, в отличие от звуков летучих мышей, имеют низкую амплитуду, широкополосный, многогармонический и частотно-модулированный. [89] Они не содержат эхолокационных щелчков с реверберацией и, по-видимому, используются для простой пространственной ориентации на близком расстоянии. В отличие от летучих мышей, землеройки используют эхолокацию только для исследования среды обитания, а не для поиска пищи. [89] Есть свидетельства того, что ослепленные лабораторные крысы могут использовать эхолокацию для навигации по лабиринтам. [93]

Некоторые насекомые, на которых охотятся летучие мыши, обладают адаптацией против хищников , включая избегание хищников, [95] отклонение атаки, [94] и ультразвуковые щелчки, которые, по-видимому, действуют как предупреждения, а не как помехи эхолокации . [49] [96]
Тигровые бабочки ( Arctiidae ) разных видов (две трети протестированных видов) реагируют на имитацию нападения эхолокационными летучими мышами, производя серию ускоряющихся щелчков. Было показано, что вид Bertholdia trigona подавляет эхолокацию летучих мышей: при столкновении с наивными большими коричневыми летучими мышами ультразвук немедленно и последовательно эффективно предотвращал нападение летучих мышей. Летучие мыши контактировали с молчаливой контрольной молью на 400% чаще, чем с B. trigona . [97]
Ультразвук моли также может пугать летучую мышь (тактика блефа), предупреждать летучую мышь о том, что моль неприятна (честная сигнализация, апосематизм ) или имитировать химически защищенные виды. Было показано, что и апосематизм, и мимикрия дают преимущество в выживании против нападения летучих мышей. [98] [99]
Большая восковая моль ( Galleria mellonella ) при обнаружении ультразвуковых волн предпринимает действия, избегающие хищников, такие как падение, петля и замирание, что указывает на то, что она может как обнаруживать, так и различать частоты ультразвука, используемые хищниками, и сигналы других представителей своего вида. [95] Некоторые представители семейства бабочек Saturniidae , в которое входят гигантские шелкопряды, имеют длинные хвосты на задних крыльях, особенно представители подгрупп Attacini и Arsenurinae . Хвосты колеблются в полете, создавая эхо, которое отражает атаку летучей мыши от тела бабочки к хвостам. Вид Argema mimosae (африканская лунная моль), обладающий особенно длинными хвостами, с наибольшей вероятностью избежал поимки. [94]
Из стр. 140: Из этих экспериментов автор заключает: …что орган слуха, по-видимому, обеспечивает зрение при обнаружении тел и снабжает этих животных различными ощущениями, позволяющими направлять их полет, и позволяет им избегать тех препятствий, которые могут возникнуть. сами себя.
Летучие мыши и дельфины известны своей способностью использовать эхолокацию. ...некоторые слепые научились делать то же самое...
