Нейтрофилы — это тип фагоцитарных белых кровяных клеток и часть врожденного иммунитета . Более конкретно, они образуют наиболее распространенный тип гранулоцитов и составляют от 40% до 70% всех белых кровяных клеток у людей. [1] Их функции различаются у разных животных. [2] Они также известны как нейтрофилы, гетерофилы или полиморфноядерные лейкоциты.
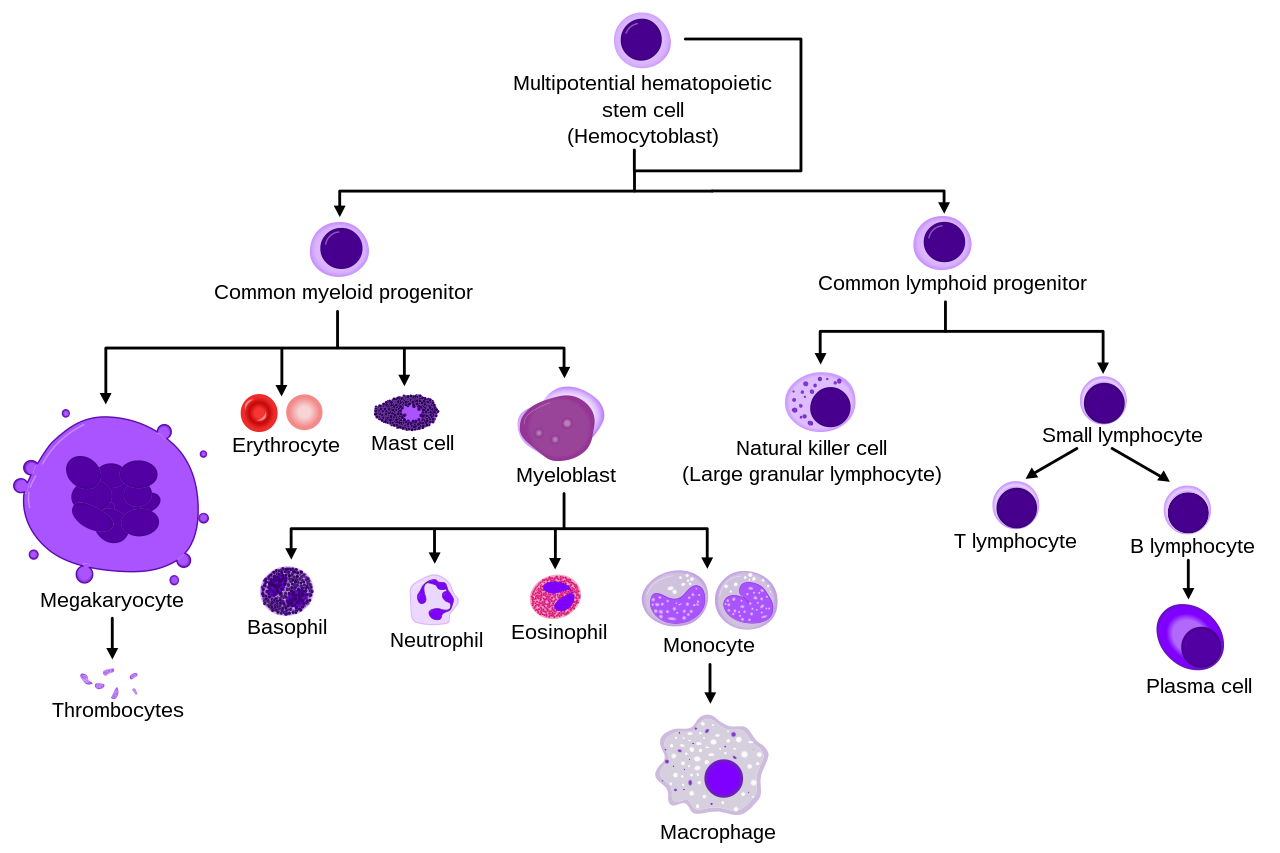
Они образуются из стволовых клеток в костном мозге и дифференцируются в субпопуляции нейтрофилов-киллеров и нейтрофилов-клеток. Они недолговечны (от 5 до 135 часов, см. § Продолжительность жизни) и очень мобильны, так как могут проникать в те части ткани, куда другие клетки/молекулы не могут. Нейтрофилы можно подразделить на сегментированные нейтрофилы и полосчатые нейтрофилы (или полосы ). Они входят в состав семейства полиморфноядерных клеток (ПМЯ) вместе с базофилами и эозинофилами . [3] [4] [5]
Название нейтрофил происходит от особенностей окрашивания гематоксилином и эозином ( H&E ) гистологических или цитологических препаратов. В то время как базофильные лейкоциты окрашиваются в темно-синий цвет, а эозинофильные лейкоциты — в ярко-красный, нейтрофилы окрашиваются в нейтральный розовый цвет. Обычно нейтрофилы содержат ядро, разделенное на 2–5 долей. [6]
Нейтрофилы являются типом фагоцитов и обычно находятся в кровотоке . Во время начальной ( острой ) фазы воспаления , особенно в результате бактериальной инфекции , воздействия окружающей среды [7] и некоторых видов рака, [8] [9] нейтрофилы являются одними из первых реагирующих на воспалительные клетки, мигрирующих к месту воспаления. Они мигрируют по кровеносным сосудам, а затем через интерстициальное пространство, следуя химическим сигналам, таким как интерлейкин-8 (IL-8), C5a , fMLP , лейкотриен B4 и перекись водорода (H 2 O 2 ) [10] в процессе, называемом хемотаксисом . Они являются преобладающими клетками в гное , что объясняет его беловатый/желтоватый вид. [11]
Нейтрофилы собираются в месте повреждения в течение нескольких минут после травмы и являются отличительным признаком острого воспаления; [12] однако, поскольку некоторые патогены не перевариваются, они не могут справиться с определенными инфекциями без помощи других типов иммунных клеток.
Структура
Нейтрофильный гранулоцит мигрирует из кровеносного сосуда в матрикс, секретируя протеолитические ферменты для растворения межклеточных связей (улучшения его подвижности) и обволакивания бактерий посредством фагоцитоза.Гиперсегментированный нейтрофил
Прикрепленные к поверхности нейтрофильные гранулоциты имеют средний диаметр 12–15 микрометров (мкм) в мазках периферической крови . В суспензии человеческие нейтрофилы имеют средний диаметр 8,85 мкм. [13]
Вместе с эозинофилами и базофилами они образуют класс полиморфноядерных клеток , названных так из-за многодольчатой формы ядра (по сравнению с лимфоцитами и моноцитами , другими типами белых клеток). Ядро имеет характерный дольчатый вид, отдельные доли соединены хроматином . Ядрышко исчезает по мере созревания нейтрофила, что происходит лишь в нескольких других типах ядросодержащих клеток. [14] : 168 До 17% ядер женских нейтрофилов человека имеют придаток в форме барабанной палочки, который содержит инактивированную Х-хромосому . [15] В цитоплазме аппарат Гольджи небольшой, митохондрии и рибосомы редки, а шероховатый эндоплазматический ретикулум отсутствует. [14] : 170 Цитоплазма также содержит около 200 гранул, из которых треть являются азурофильными . [14] : 170
Нейтрофилы будут показывать увеличивающуюся сегментацию (множество сегментов ядра) по мере созревания. Нормальный нейтрофил должен иметь 3–5 сегментов. Гиперсегментация не является нормой, но встречается при некоторых расстройствах, в частности, при дефиците витамина B 12. Это отмечается при ручном просмотре мазка крови и является положительным, когда большинство или все нейтрофилы имеют 5 или более сегментов.
Нейтрофилы являются наиболее распространенными белыми кровяными клетками в организме человека (ежедневно вырабатывается около 10 11 ); они составляют около 50–70 % всех белых кровяных клеток (лейкоцитов). Указанный нормальный диапазон для показателей крови человека варьируется в зависимости от лаборатории, но количество нейтрофилов 2,5–7,5 × 10 9 / л является стандартным нормальным диапазоном. У людей африканского и ближневосточного происхождения могут быть более низкие показатели, которые все равно являются нормальными. [16] В отчете нейтрофилы могут быть разделены на сегментированные и палочкоядерные .
При циркуляции в кровотоке и инактивации нейтрофилы имеют сферическую форму. После активации они меняют форму и становятся более аморфными или амебоподобными и могут расширять псевдоподии , охотясь за антигенами . [17]
Способность нейтрофилов поглощать бактерии снижается при употреблении простых сахаров, таких как глюкоза, фруктоза, а также сахароза, мед и апельсиновый сок, в то время как употребление крахмалов не оказывает никакого эффекта. С другой стороны, голодание усиливает фагоцитарную способность нейтрофилов поглощать бактерии. Был сделан вывод, что функция, а не количество фагоцитов при поглощении бактерий изменяется при употреблении сахаров. [18] В 2007 году исследователи из Института биомедицинских исследований Уайтхеда обнаружили, что при выборе сахаров на микробных поверхностях нейтрофилы реагируют преимущественно на некоторые типы сахаров. Нейтрофилы преимущественно поглощают и убивают мишени бета-1,6-глюкана по сравнению с мишенями бета-1,3-глюкана. [19] [20]
Согласно разным данным, средняя продолжительность жизни инактивированных человеческих нейтрофилов в кровотоке составляет от 5 до 135 часов. [21] [22]
После активации они маргинализируются (размещаются рядом с эндотелием кровеносных сосудов) и подвергаются селективно -зависимому захвату с последующей интегрин -зависимой адгезией в большинстве случаев, после чего они мигрируют в ткани, где выживают в течение 1–2 дней. [23] Также было показано, что нейтрофилы высвобождаются в кровь из селезеночного резерва после инфаркта миокарда . [24]
Соотношение распределения нейтрофилов в костном мозге, крови и соединительной ткани составляет 28:1:25.
Нейтрофилы гораздо многочисленнее, чем долгоживущие моноциты / макрофаги фагоцитов. Патоген (болезнетворный микроорганизм или вирус), скорее всего, первым встретится с нейтрофилом. Некоторые эксперты предполагают, что короткое время жизни нейтрофилов является эволюционной адаптацией. Короткое время жизни нейтрофилов сводит к минимуму распространение тех патогенов, которые паразитируют на фагоцитах (например, Leishmania [25] ), поскольку чем больше времени такие паразиты проводят вне клетки-хозяина , тем больше вероятность того, что они будут уничтожены каким-либо компонентом защитных сил организма. Кроме того, поскольку нейтрофильные антимикробные продукты также могут повреждать ткани хозяина , их короткая жизнь ограничивает повреждение хозяина во время воспаления . [23]
Нейтрофилы подвергаются процессу, называемому хемотаксисом , посредством амебоидного движения , что позволяет им мигрировать к местам инфекции или воспаления. Рецепторы клеточной поверхности позволяют нейтрофилам обнаруживать химические градиенты молекул, таких как интерлейкин-8 (ИЛ-8), интерферон гамма (ИФН-γ), C3a, C5a и лейкотриен B4 , которые эти клетки используют для определения пути своей миграции.
Нейтрофилы имеют множество специфических рецепторов, включая рецепторы для комплемента , цитокинов, таких как интерлейкины и IFN-γ, хемокинов , лектинов и других белков. Они также экспрессируют рецепторы для обнаружения и присоединения к эндотелию и Fc-рецепторы для опсонина . [26]
В лейкоцитах, реагирующих на хемоаттрактант , клеточная полярность регулируется активностью малых Ras или Rho гуанозинтрифосфатаз (Ras или Rho GTPases ) и фосфоинозитид 3-киназ ( PI3K ). В нейтрофилах липидные продукты PI3K регулируют активацию Rac1, гемопоэтических Rac2 и RhoG GTPases семейства Rho и необходимы для подвижности клеток . Ras-GTPases и Rac-GTPases регулируют динамику цитоскелета и способствуют адгезии, миграции и распространению нейтрофилов. [27] [28] [29] Они накапливаются асимметрично плазматической мембране на переднем крае поляризованных клеток. Пространственно регулируя Rho ГТФазы и организуя передний край клетки, PI3K и их липидные продукты могут играть ключевую роль в установлении полярности лейкоцитов, выступая в качестве молекул-компасов, указывающих клетке, куда ей двигаться.
На мышах было показано, что при определенных условиях нейтрофилы демонстрируют особый тип миграционного поведения, называемый роением нейтрофилов, во время которого они мигрируют высокоскоординированным образом, накапливаются и группируются в местах воспаления. [30]
Антимикробная функция
Будучи очень подвижными , нейтрофилы быстро собираются в очаге инфекции , привлекаемые цитокинами , экспрессируемыми активированным эндотелием , тучными клетками и макрофагами . Нейтрофилы экспрессируют [31] и выделяют цитокины, которые, в свою очередь, усиливают воспалительные реакции со стороны нескольких других типов клеток.
Помимо привлечения и активации других клеток иммунной системы, нейтрофилы играют ключевую роль в защите первой линии от вторжения патогенов и содержат широкий спектр белков. [32] У нейтрофилов есть три метода для прямой атаки на микроорганизмы: фагоцитоз (поглощение), дегрануляция (выделение растворимых антимикробных препаратов) и генерация нейтрофильных внеклеточных ловушек (НВЛ). [33]
Нейтрофилы — это фагоциты , способные поглощать микроорганизмы или частицы. Для распознавания целей они должны быть покрыты опсонинами — процесс, известный как опсонизация антител . [17] Они могут поглощать и убивать множество микробов , каждое фагоцитарное событие приводит к образованию фагосомы , в которую секретируются активные формы кислорода и гидролитические ферменты. Потребление кислорода во время генерации активных форм кислорода было названо « респираторным взрывом », хотя и не связано с дыханием или выработкой энергии.
Респираторный взрыв включает активацию фермента НАДФН -оксидазы , который производит большое количество супероксида , реактивного вида кислорода. Супероксид распадается спонтанно или расщепляется с помощью ферментов, известных как супероксиддисмутазы (Cu/ZnSOD и MnSOD), до перекиси водорода, которая затем преобразуется в хлорноватистую кислоту (HClO) зеленым гемовым ферментом миелопероксидазой . Считается, что бактерицидных свойств HClO достаточно, чтобы убить бактерии, фагоцитированные нейтрофилами, но вместо этого это может быть шагом, необходимым для активации протеаз. [34]
Хотя нейтрофилы могут убивать многие микробы, взаимодействие нейтрофилов с микробами и молекулами, продуцируемыми микробами, часто изменяет оборот нейтрофилов. Способность микробов изменять судьбу нейтрофилов весьма разнообразна, может быть специфичной для микробов и варьируется от продления продолжительности жизни нейтрофилов до быстрого лизиса нейтрофилов после фагоцитоза. Сообщалось, что Chlamydia pneumoniae и Neisseria gonorrhoeae задерживают апоптоз нейтрофилов . [35] [36] [37] Таким образом, некоторые бактерии — и те, которые являются преимущественно внутриклеточными патогенами — могут продлевать продолжительность жизни нейтрофилов, нарушая нормальный процесс спонтанного апоптоза и/или PICD (фагоцитоз-индуцированной гибели клеток). С другой стороны, некоторые патогены, такие как Streptococcus pyogenes, способны изменять судьбу нейтрофилов после фагоцитоза, способствуя быстрому лизису клеток и/или ускоряя апоптоз до точки вторичного некроза. [38] [39]
Дегрануляция
Нейтрофилы также высвобождают набор белков в трех типах гранул с помощью процесса, называемого дегрануляцией . Содержимое этих гранул обладает антимикробными свойствами и помогает бороться с инфекцией. Клетки-блестки представляют собой полиморфноядерные лейкоцитарные нейтрофилы с гранулами. [40]
Нейтрофильные внеклеточные ловушки
В 2004 году Бринкманн и его коллеги описали поразительное наблюдение, что активация нейтрофилов вызывает высвобождение паутинных структур ДНК; это представляет собой третий механизм уничтожения бактерий. [42] Эти нейтрофильные внеклеточные ловушки (NET) представляют собой сеть волокон, состоящих из хроматина и сериновых протеаз [43] , которые улавливают и убивают внеклеточные микробы. Предполагается, что NET обеспечивают высокую локальную концентрацию антимикробных компонентов и связывают, обезвреживают и убивают микробы независимо от фагоцитарного поглощения. В дополнение к их возможным антимикробным свойствам, NET могут служить физическим барьером, который предотвращает дальнейшее распространение патогенов. Улавливание бактерий может быть особенно важной ролью NET при сепсисе , когда NET образуются внутри кровеносных сосудов. [44] Наконец, было продемонстрировано, что образование NET усиливает бактерицидную активность макрофагов во время инфекции. [45] [46] Недавно было показано, что NET играют роль в воспалительных заболеваниях, поскольку NET могут быть обнаружены при преэклампсии , воспалительном заболевании, связанном с беременностью, при котором, как известно, активируются нейтрофилы. [47] Образование нейтрофильных NET также может влиять на сердечно-сосудистые заболевания , поскольку NET могут влиять на образование тромбов в коронарных артериях . [48] [49]
Теперь известно, что NET проявляют протромботические эффекты как in vitro [50] , так и in vivo . [51] [52] Совсем недавно, в 2020 году, NET были вовлечены в образование тромбов в случаях тяжелого течения COVID-19 . [53]
Нейтрофилы, ассоциированные с опухолью (TANS)
TAN могут демонстрировать повышенную скорость внеклеточного закисления при повышении уровней гликолиза. [54] Когда происходит метаболический сдвиг в TAN, это может привести к прогрессированию опухоли в определенных областях тела, таких как легкие. TAN поддерживают рост и прогрессирование опухолей в отличие от обычных нейтрофилов, которые ингибируют прогрессирование опухоли посредством фагоцитоза опухолевых клеток. Используя модель мыши, они [ кто? ] определили, что как метаболизм Glut1, так и метаболизм глюкозы увеличились в TAN, обнаруженных у мыши, у которой была аденокарцинома легкого. [54] Исследование показало, что клетки опухоли легкого могут удаленно инициировать остеобласты, и эти остеобласты могут ухудшать опухоли двумя способами. Во-первых, они могут индуцировать образование нейтрофилов с высокой экспрессией SiglecF, что, в свою очередь, способствует росту и прогрессированию опухоли легкого. Во-вторых, остеобласты могут способствовать росту костей, тем самым формируя благоприятную среду для роста опухолевых клеток с образованием метастазов в кости. [55]
Клиническое значение
Микрофотография, показывающая несколько нейтрофилов во время острого воспаления.
Низкое количество нейтрофилов называется нейтропенией . Это может быть врожденным (развиваться при рождении или до него) или может развиться позже, как в случае апластической анемии или некоторых видов лейкемии . Это также может быть побочным эффектом лекарств , в первую очередь химиотерапии . Нейтропения делает человека очень восприимчивым к инфекциям. Это также может быть результатом колонизации внутриклеточными нейтрофильными паразитами.
При дефиците альфа-1-антитрипсина важная нейтрофильная эластаза не ингибируется должным образом альфа-1-антитрипсином , что приводит к чрезмерному повреждению тканей при наличии воспаления – наиболее заметным из которых является эмфизема . Отрицательные эффекты эластазы также были показаны в случаях, когда нейтрофилы чрезмерно активированы (у в остальном здоровых людей) и высвобождают фермент во внеклеточное пространство. Нерегулируемая активность нейтрофильной эластазы может привести к нарушению легочного барьера, проявляя симптомы, соответствующие острому повреждению легких . [56] Фермент также влияет на активность макрофагов, расщепляя их толл-подобные рецепторы (TLR) и снижая экспрессию цитокинов путем ингибирования ядерной транслокации NF-κB . [57]
Гипергликемия может привести к дисфункции нейтрофилов. Дисфункция в биохимическом пути нейтрофилов миелопероксидазы , а также сниженная дегрануляция связаны с гипергликемией. [59]
Абсолютное количество нейтрофилов (ANC) также используется в диагностике и прогнозировании. ANC является золотым стандартом для определения тяжести нейтропении и, следовательно, нейтропенической лихорадки. Любое ANC < 1500 клеток / мм 3 считается нейтропенией, но < 500 клеток / мм 3 считается тяжелой. [60] Также есть новые исследования, связывающие ANC с инфарктом миокарда в качестве помощи в ранней диагностике. [61] [62] Нейтрофилы способствуют желудочковой тахикардии при остром инфаркте миокарда. [63]
При инсульте они начинают проникать в пораженный инфарктом мозг через 6–8 часов. [65]
Уклонение от патогенов и резистентность
Подобно фагоцитам, патогены могут уклоняться от нейтрофилов или инфицировать их. [66] Некоторые бактериальные патогены выработали различные механизмы, такие как молекулы вирулентности, чтобы избежать гибели от нейтрофилов. Эти молекулы в совокупности могут изменять или нарушать набор нейтрофилов, апоптоз или бактерицидную активность. [66]
Нейтрофилы также могут служить клетками-хозяевами для различных паразитов, которые заражают их, избегая фагоцитоза, в том числе:
Leishmania major – использует нейтрофилы в качестве средства для паразитирования на фагоцитах [25]
Существует пять (HNA 1–5) наборов нейтрофильных антигенов. Три антигена HNA-1 (ac) расположены на низкоаффинном рецепторе Fc-γ IIIb (FCGR3B: CD16b ). Единственный известный антиген HNA-2a расположен на CD177 . Система антигенов HNA-3 имеет два антигена (3a и 3b), которые расположены на седьмом экзоне гена CLT2 ( SLC44A2 ). Системы антигенов HNA-4 и HNA-5 имеют по два известных антигена (a и b) и расположены в интегрине β2 . HNA-4 расположен на цепи αM ( CD11b ), а HNA-5 расположен на единице интегрина αL ( CD11a ). [68]
Субпопуляции
Активность нейтрофилов-киллеров и нейтрофилов-клеток в тесте НСТ [69]
Были идентифицированы две функционально неравные субпопуляции нейтрофилов на основе разного уровня генерации ими активных форм кислорода, проницаемости мембраны, активности ферментной системы и способности к инактивации. Клетки одной субпопуляции с высокой проницаемостью мембраны (нейтрофилы-киллеры) интенсивно генерируют активные формы кислорода и инактивируются в результате взаимодействия с субстратом, тогда как клетки другой субпопуляции (нейтрофилы-клетки) продуцируют активные формы кислорода менее интенсивно, не прилипают к субстрату и сохраняют свою активность. [69] [70] [71] [72] [73] Дополнительные исследования показали, что опухоли легких могут быть инфильтрированы различными популяциями нейтрофилов. [74]
Здесь можно увидеть нейтрофил, избирательно захватывающий несколько дрожжей Candida ( флуоресцентно помеченных зеленым цветом), несмотря на несколько контактов с конидиями Aspergillus fumigatus (не помеченными, белыми/прозрачными) в 3-D коллагеновой матрице. Время визуализации составило 2 часа с одним кадром каждые 30 секунд.
Нейтрофилы демонстрируют высоконаправленную амебоидную подвижность в инфицированных подушечках лап и фалангах. Интравитальная визуализация проводилась в пути подушечки лап мышей LysM-eGFP через 20 минут после заражения Listeria monocytogenes . [75]
^ Актер Дж. (2012). Интегрированный обзор иммунологии и микробиологии издательства Elsevier (второе издание). doi :10.1016/B978-0-323-07447-6.00002-8.
^ Эрмерт Д., Нимиец М.Дж., Рем М., Глентхой А., Боррегор Н., Урбан CF (август 2013 г.). «Candida albicans ускользает от нейтрофилов мыши». Журнал биологии лейкоцитов . 94 (2): 223–236. дои : 10.1189/jlb.0213063. PMID 23650619. S2CID 25619835.
^ Witko-Sarsat V, Rieu P, Descamps-Latscha B, Lesavre P, Halbwachs-Mecarelli L (май 2000 г.). «Нейтрофилы: молекулы, функции и патофизиологические аспекты». Лабораторные исследования; Журнал технических методов и патологии . 80 (5): 617–653. doi : 10.1038/labinvest.3780067 . PMID 10830774. S2CID 22536645.
^ Клебанофф С.Дж., Кларк РА (1978). Нейтрофил: функция и клинические расстройства . Elsevier/North-Holland Amsterdam. ISBN978-0-444-80020-6.
^ Nathan C (март 2006 г.). «Нейтрофилы и иммунитет: проблемы и возможности». Nature Reviews. Иммунология . 6 (3): 173–182. doi :10.1038/nri1785. PMID 16498448. S2CID 1590558.
^ Welsh CJ (2021). Основы анатомии и физиологии человека Хоула (14-е изд.). Нью-Йорк, США: McGraw Hill. стр. 336. ISBN978-1-260-57521-7. Получено 28 февраля 2023 г. .
^ Jacobs L, Nawrot TS, de Geus B, Meeusen R, Degraeuwe B, Bernard A и др. (октябрь 2010 г.). «Субклинические реакции у здоровых велосипедистов, кратковременно подвергавшихся загрязнению воздуха, связанному с транспортом: исследование вмешательства». Environmental Health . 9 (64): 64. Bibcode :2010EnvHe...9...64J. doi : 10.1186/1476-069X-9-64 . PMC 2984475 . PMID 20973949.
^ Waugh DJ, Wilson C (ноябрь 2008 г.). «Путь интерлейкина-8 при раке». Clinical Cancer Research . 14 (21): 6735–6741. doi :10.1158/1078-0432.CCR-07-4843. PMID 18980965. S2CID 9415085.
^ De Larco JE, Wuertz BR, Furcht LT (август 2004 г.). «Потенциальная роль нейтрофилов в продвижении метастатического фенотипа опухолей, высвобождающих интерлейкин-8». Clinical Cancer Research . 10 (15): 4895–4900. doi :10.1158/1078-0432.CCR-03-0760. PMID 15297389. S2CID 9782495.
^ Yoo SK, Starnes TW, Deng Q, Huttenlocher A (ноябрь 2011 г.). «Lyn — это окислительно-восстановительный сенсор, который опосредует притяжение ран лейкоцитов in vivo». Nature . 480 (7375): 109–112. Bibcode :2011Natur.480..109Y. doi :10.1038/nature10632. PMC 3228893 . PMID 22101434.
^ Barer MR (2012). «Естественная история инфекции». Медицинская микробиология . Elsevier. стр. 168–173. doi :10.1016/b978-0-7020-4089-4.00029-9. ISBN978-0-7020-4089-4.
^ Коэн С., Бернс Р. К. (2002). Пути целлюлозы (8-е изд.). Сент-Луис: Мосби. С. 465.
^ Ниемец М.Дж., Де Самбер Б., Гарревет Дж., Вергухт Э., Векеманс Б., Де Райке Р. и др. (июнь 2015 г.). «Микроэлементный ландшафт покоящихся и активированных нейтрофилов человека на субмикрометровом уровне». Металломика . 7 (6): 996–1010. дои : 10.1039/c4mt00346b . ПМИД 25832493.
^ abc Zucker-Franklin D, Greaves MF, Grossi CE, Marmont AM (1988). "Нейтрофилы". Атлас клеток крови: функции и патологии . Том 1 (2-е изд.). Филадельфия: Lea & Ferbiger. ISBN978-0-8121-1094-4.
^ Karni RJ, Wangh LJ, Sanchez JA (август 2001 г.). «Неслучайное расположение и ориентация неактивной X-хромосомы в ядрах нейтрофилов человека». Chromosoma . 110 (4): 267–274. doi :10.1007/s004120100145. PMID 11534818. S2CID 24750407.
^ Reich D, Nalls MA, Kao WH, Akylbekova EL, Tandon A, Patterson N и др. (январь 2009 г.). «Снижение количества нейтрофилов у людей африканского происхождения обусловлено регуляторным вариантом в гене рецептора антигена Даффи для хемокинов». PLOS Genetics . 5 (1): e1000360. doi : 10.1371/journal.pgen.1000360 . PMC 2628742 . PMID 19180233.
^ ab Edwards SW (1994). Биохимия и физиология нейтрофилов . Cambridge University Press. стр. 6. ISBN978-0-521-41698-6.
^ Sanchez A, Reeser JL, Lau HS, Yahiku PY, Willard RE, McMillan PJ и др. (ноябрь 1973 г.). «Роль сахаров в человеческом нейтрофильном фагоцитозе». The American Journal of Clinical Nutrition . 26 (11): 1180–1184. doi : 10.1093/ajcn/26.11.1180 . PMID 4748178. Эти данные свидетельствуют о том, что функция, а не количество фагоцитов были изменены при приеме сахаров. Это подразумевает глюкозу и другие простые углеводы в контроле фагоцитоза и показывает, что эффекты длятся по крайней мере 5 часов. С другой стороны, голодание в течение 36 или 60 часов значительно увеличивало (P < 0,001) фагоцитарный индекс
^ Rubin-Bejerano I, Abeijon C, Magnelli P, Grisafi P, Fink GR (июль 2007 г.). «Фагоцитоз человеческими нейтрофилами стимулируется уникальным компонентом клеточной стенки грибка». Cell Host & Microbe . 2 (1): 55–67. doi :10.1016/j.chom.2007.06.002. PMC 2083279 . PMID 18005717.
^ Кнеллер А. (2007). «Белые кровяные клетки разборчивы в отношении сахара». Институт Уайтхеда . Получено 09.08.2013 .
^ Tak T, Tesselaar K, Pillay J, Borghans JA, Koenderman L (октябрь 2013 г.). «Сколько вам лет? Повторное определение полупериода жизни нейтрофилов человека». Journal of Leukocyte Biology . 94 (4): 595–601. doi :10.1189/jlb.1112571. PMID 23625199. S2CID 40113921.
^ Pillay J, den Braber I, Vrisekoop N, Kwast LM, de Boer RJ, Borghans JA и др. (Июль 2010 г.). «In vivo маркировка с помощью 2H2O выявляет продолжительность жизни человеческих нейтрофилов в 5,4 дня». Blood . 116 (4): 625–627. doi : 10.1182/blood-2010-01-259028 . PMID 20410504. S2CID 909519.
^ ab Wheater PR, Stevens A (2002). Базовая гистопатология Wheater: цветной атлас и текст . Эдинбург: Churchill Livingstone. ISBN978-0-443-07001-3.
^ Akbar N, Braithwaite AT, Corr EM, Koelwyn GJ, van Solingen C, Cochain C и др. (март 2023 г.). «Быстрая мобилизация нейтрофилов внеклеточными везикулами, полученными из эндотелиальных клеток VCAM-1+». Cardiovascular Research . 119 (1): 236–251. doi :10.1093/cvr/cvac012. PMC 10022859 . PMID 35134856.
^ ab Ritter U, Frischknecht F, van Zandbergen G (ноябрь 2009 г.). «Являются ли нейтрофилы важными клетками-хозяевами для паразитов Leishmania?». Trends in Parasitology . 25 (11): 505–510. doi :10.1016/j.pt.2009.08.003. PMID 19762280.
^ Serhan CN, Ward PA, Gilroy DW (2010). Основы воспаления. Cambridge University Press. С. 53–54. ISBN978-0-521-88729-8.
^ Pantarelli C, Welch HC (ноябрь 2018 г.). «Rac-GTPases и Rac-GEFs в адгезии, миграции и рекрутинге нейтрофилов». European Journal of Clinical Investigation . 48 (Suppl 2): e12939. doi :10.1111/eci.12939. PMC 6321979 . PMID 29682742.
^ Lin Y, Pal DS, Banerjee P, Banerjee T, Qin G, Deng Y и др. (Июль 2024 г.). «Подавление Ras усиливает поляризацию и миграцию клеток, вызванную контрактильностью заднего актомиозина». Nature Cell Biology : 1–15. doi :10.1038/s41556-024-01453-4. PMID 38951708.
^ Pal DS, Banerjee T, Lin Y, de Trogoff F, Borleis J, Iglesias PA и др. (Июль 2023 г.). «Активация отдельных нисходящих узлов в сети факторов роста управляет миграцией иммунных клеток». Developmental Cell . 58 (13): 1170–1188.e7. doi :10.1016/j.devcel.2023.04.019. PMC 10524337 . PMID 37220748.
^ Lämmermann T, Afonso PV, Angermann BR, Wang JM, Kastenmüller W, Parent CA и др. (июнь 2013 г.). «Нейтрофильные рои требуют LTB4 и интегрины в местах гибели клеток in vivo». Nature . 498 (7454): 371–375. Bibcode :2013Natur.498..371L. doi :10.1038/nature12175. PMC 3879961 . PMID 23708969.
^ Ear T, McDonald PP (апрель 2008 г.). «Генерация цитокинов, активация промотора и независимая от окислителей активация NF-kappaB в трансфицируемой модели человеческих нейтрофильных клеток». BMC Immunology . 9 : 14. doi : 10.1186/1471-2172-9-14 . PMC 2322942. PMID 18405381 .
^ Ambatipudi KS, Old JM, Guilhaus M, Raftery M, Hinds L, Deane EM (2006). Протеомный анализ нейтрофильных белков таммарского валлаби ( Macropus eugenii ). Сравнительная биохимия и физиология. Часть D: Геномика и протеомика. 1(3), 283-291. DOI: 10.1016/j.cbd.2006.05.002
^ Хики М.Дж., Кубес П. (май 2009 г.). «Внутрисосудистый иммунитет: встреча хозяина и патогена в кровеносных сосудах». Nature Reviews. Иммунология . 9 (5): 364–375. doi :10.1038/nri2532. PMID 19390567. S2CID 8068543.
^ Segal AW (2005). «Как нейтрофилы убивают микробы». Annual Review of Immunology . 23 (5): 197–223. doi :10.1146/annurev.immunol.23.021704.115653. PMC 2092448. PMID 15771570 .
^ Simons MP, Nauseef WM, Griffith TS, Apicella MA (ноябрь 2006 г.). «Neisseria gonorrhoeae задерживает начало апоптоза в полиморфноядерных лейкоцитах». Cellular Microbiology . 8 (11): 1780–1790. doi :10.1111/j.1462-5822.2006.00748.x. PMID 16803582. S2CID 25253422.
^ Chen A, Seifert HS (ноябрь 2011 г.). «Ингибирование апоптотической сигнализации в полиморфноядерных лейкоцитах, опосредованное Neisseria gonorrhoeae». Инфекция и иммунитет . 79 (11): 4447–4458. doi :10.1128/IAI.01267-10. PMC 3257915. PMID 21844239 .
^ van Zandbergen G, Gieffers J, Kothe H, Rupp J, Bollinger A, Aga E и др. (февраль 2004 г.). «Chlamydia pneumoniae размножаются в нейтрофильных гранулоцитах и задерживают их спонтанный апоптоз». Журнал иммунологии . 172 (3): 1768–1776. doi : 10.4049/jimmunol.172.3.1768 . PMID 14734760. S2CID 27422510.
^ Kobayashi SD, Braughton KR, Whitney AR, Voyich JM, Schwan TG, Musser JM и др. (сентябрь 2003 г.). «Бактериальные патогены модулируют программу дифференциации апоптоза в человеческих нейтрофилах». Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 10948–10953. doi : 10.1073/pnas.1833375100 . PMC 196908. PMID 12960399.
^ Kobayashi SD, Malachowa N, DeLeo FR (2017). «Влияние микробов на жизнь и смерть нейтрофилов». Frontiers in Cellular and Infection Microbiology . 7 (159): 159. doi : 10.3389/fcimb.2017.00159 . PMC 5410578. PMID 28507953.Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ Berman LB, Feys JO, Schreiner GE (ноябрь 1956 г.). «Наблюдения за феноменом блестящей клетки». The New England Journal of Medicine . 255 (21): 989–991. doi :10.1056/NEJM195611222552104. PMID 13378597.
^ Бринкманн В., Рейхард У., Гусманн К., Фаулер Б., Улеманн И., Вайс Д.С. и др. (март 2004 г.). «Нейтрофильные внеклеточные ловушки убивают бактерии». Science . 303 (5663): 1532–1535. Bibcode :2004Sci...303.1532B. doi :10.1126/science.1092385. PMID 15001782. S2CID 21628300.
^ Urban CF, Ermert D, Schmid M, Abu-Abed U, Goosmann C, Nacken W и др. (октябрь 2009 г.). «Нейтрофильные внеклеточные ловушки содержат кальпротектин, цитозольный белковый комплекс, участвующий в защите хозяина от Candida albicans». PLOS Pathogens . 5 (10): e1000639. doi : 10.1371/journal.ppat.1000639 . PMC 2763347. PMID 19876394 .
^ Clark SR, Ma AC, Tavener SA, McDonald B, Goodarzi Z, Kelly MM и др. (апрель 2007 г.). «TLR4 тромбоцитов активирует внеклеточные ловушки нейтрофилов, чтобы заманить бактерии в септическую кровь». Nature Medicine . 13 (4): 463–469. doi :10.1038/nm1565. PMID 17384648. S2CID 22372863.
^ Monteith AJ, Miller JM, Maxwell CN, Chazin WJ, Skaar EP (сентябрь 2021 г.). «Нейтрофильные внеклеточные ловушки усиливают уничтожение бактериальных патогенов макрофагами». Science Advances . 7 (37): eabj2101. Bibcode :2021SciA....7.2101M. doi :10.1126/sciadv.abj2101. PMC 8442908 . PMID 34516771.
^ Monteith AJ, Miller JM, Beavers WN, Maloney KN, Seifert EL, Hajnoczky G и др. (февраль 2022 г.). «Митохондриальный унипортер кальция влияет на бактерицидную активность нейтрофилов во время инфекции Staphylococcus aureus». Инфекция и иммунитет . 90 (2): e0055121. doi :10.1128/IAI.00551-21. PMC 8853686. PMID 34871043 .
^ Gupta AK, Hasler P, Holzgreve W, Hahn S (июнь 2007 г.). «Нейтрофильные НЭТ: новый фактор, способствующий плацентарной гипоксии, связанной с преэклампсией?». Семинары по иммунопатологии . 29 (2): 163–167. doi :10.1007/s00281-007-0073-4. PMID 17621701. S2CID 12887059.
^ Хойер ФФ, Нарендорф М (февраль 2017 г.). «Вклад нейтрофилов в ишемическую болезнь сердца». European Heart Journal . 38 (7): 465–472. doi : 10.1093/eurheartj/ehx017 . PMID 28363210.
^ Mangold A, Alias S, Scherz T, Hofbauer M, Jakowitsch J, Panzenböck A и др. (март 2015 г.). «Нагрузка внеклеточной ловушки коронарных нейтрофилов и активность дезоксирибонуклеазы при остром коронарном синдроме с подъемом ST являются предикторами разрешения сегмента ST и размера инфаркта». Circulation Research . 116 (7): 1182–1192. doi : 10.1161/CIRCRESAHA.116.304944 . PMID 25547404. S2CID 2532741.
^ Fuchs TA, Brill A, Duerschmied D, Schatzberg D, Monestier M, Myers DD и др. (сентябрь 2010 г.). «Внеклеточные ловушки ДНК способствуют тромбозу». Труды Национальной академии наук Соединенных Штатов Америки . 107 (36): 15880–15885. Bibcode : 2010PNAS..10715880F. doi : 10.1073/pnas.1005743107 . PMC 2936604. PMID 20798043 .
^ Brill A, Fuchs TA, Savchenko AS, Thomas GM, Martinod K, De Meyer SF и др. (январь 2012 г.). «Нейтрофильные внеклеточные ловушки способствуют тромбозу глубоких вен у мышей». Журнал тромбоза и гемостаза . 10 (1): 136–144. doi :10.1111/j.1538-7836.2011.04544.x. PMC 3319651. PMID 22044575 .
^ Бориссофф JI, тен Кейт H (сентябрь 2011 г.). «От высвобождения нейтрофильных внеклеточных ловушек к тромбозу: избыточный механизм защиты хозяина?». Журнал тромбоза и гемостаза . 9 (9): 1791–1794. doi : 10.1111/j.1538-7836.2011.04425.x . PMID 21718435. S2CID 5368241.
^ Цзо И., Ялаварти С., Ши Х., Гокман К., Цзо М., Мэдисон Дж.А. и др. (июнь 2020 г.). «Нейтрофильные внеклеточные ловушки при COVID-19». JCI-инсайт . 5 (11): e138999. doi : 10.1172/jci.insight.138999. ПМК 7308057 . PMID 32329756. S2CID 216109364.
^ ab Ancey PB, Contat C, Boivin G, Sabatino S, Pascual J, Zangger N, et al. (Май 2021 г.). «Экспрессия GLUT1 в нейтрофилах, связанных с опухолью, способствует росту рака легких и устойчивости к радиотерапии». Cancer Research . 81 (9): 2345–2357. doi :10.1158/0008-5472.CAN-20-2870. PMC 8137580 . PMID 33753374.
^ Azevedo PO, Paiva AE, Santos GS, Lousado L, Andreotti JP, Sena IF и др. (декабрь 2018 г.). «Перекрестные помехи между раком легких и костями приводят к образованию нейтрофилов, способствующих прогрессированию опухоли». Cancer and Metastasis Reviews . 37 (4): 779–790. doi :10.1007/s10555-018-9759-4. PMC 6358512 . PMID 30203108.
^ Кавабата К, Хагио Т, Мацуока С (сентябрь 2002 г.). «Роль нейтрофильной эластазы при остром повреждении легких». Европейский журнал фармакологии . 451 (1): 1–10. doi :10.1016/S0014-2999(02)02182-9. PMID 12223222.
^ Domon H, Nagai K, Maekawa T, Oda M, Yonezawa D, Takeda W и др. (2018). «Нейтрофильная эластаза подрывает иммунный ответ путем расщепления Toll-подобных рецепторов и цитокинов при пневмококковой пневмонии». Frontiers in Immunology . 9 : 732. doi : 10.3389/fimmu.2018.00732 . PMC 5996908. PMID 29922273.
^ Озен С. (июль 2003 г.). «Семейная средиземноморская лихорадка: пересмотр древней болезни». Европейский журнал педиатрии . 162 (7–8): 449–454. doi :10.1007/s00431-003-1223-x. PMID 12751000. S2CID 3464945.
^ Xiu F, Stanojcic M, Diao L, Jeschke MG (8 мая 2014 г.). «Стрессовая гипергликемия, лечение инсулином и врожденные иммунные клетки». Международный журнал эндокринологии . 2014 : 486403. doi : 10.1155/2014/486403 . PMC 4034653. PMID 24899891 .
^ Al-Gwaiz LA, Babay HH (2007). «Диагностическая ценность абсолютного количества нейтрофилов, количества палочкоядерных нейтрофилов и морфологических изменений нейтрофилов в прогнозировании бактериальных инфекций». Medical Principles and Practice . 16 (5): 344–347. doi : 10.1159/000104806 . PMID 17709921. S2CID 5499290.
^ Хан HA, Алхомида AS, Собки SH, Могайри AA, Коронки HE (2012). «Количество клеток крови и их корреляция с креатинкиназой и С-реактивным белком у пациентов с острым инфарктом миокарда». Международный журнал клинической и экспериментальной медицины . 5 (1): 50–55. PMC 3272686. PMID 22328948 .
^ Базили С., Ди Франко М., Роза А., Феррони П., Дюрни В., Скарпеллини М.Г. и др. (апрель 2004 г.). «Абсолютное количество нейтрофилов и уровень фибриногена как помощь в ранней диагностике острого инфаркта миокарда». Акта Кардиологика . 59 (2): 135–140. дои : 10.2143/ac.59.2.2005167. PMID 15139653. S2CID 37382677.
^ Grune J, Lewis AJ, Yamazoe M, Hulsmans M, Rohde D, Xiao L и др. (Июль 2022 г.). «Нейтрофилы стимулируют, а макрофаги предотвращают электрический шторм после инфаркта миокарда». Nature Cardiovascular Research . 1 (7): 649–664. doi :10.1038/s44161-022-00094-w. PMC 9410341. PMID 36034743 .
^ Michaud K, Basso C, d'Amati G, Giordano C, Kholová I, Preston SD и др. (февраль 2020 г.). «Диагностика инфаркта миокарда при аутопсии: переоценка AECVP в свете текущей клинической классификации». Virchows Archiv . 476 (2): 179–194. doi : 10.1007 /s00428-019-02662-1 . PMC 7028821. PMID 31522288. «Данная статья распространяется на условиях лицензии Creative Commons Attribution 4.0 International (http://creativecommons.org/licenses/by/4.0/ Архивировано 21 ноября 2015 г. на Wayback Machine )»
^ Jickling GC, Liu D, Ander BP, Stamova B, Zhan X, Sharp FR (июнь 2015 г.). «Нацеливание на нейтрофилы при ишемическом инсульте: трансляционные идеи экспериментальных исследований». Журнал мозгового кровотока и метаболизма . 35 (6): 888–901. doi :10.1038/jcbfm.2015.45. PMC 4640255. PMID 25806703 .
^ ab Kobayashi SD, Malachowa N, DeLeo FR (2018). «Нейтрофилы и уклонение от бактериального иммунитета». Журнал врожденного иммунитета . 10 (5–6): 432–441. doi :10.1159/000487756. PMC 6784029. PMID 29642066 .
^ abcd Parker HA, Forrester L, Kaldor CD, Dickerhof N, Hampton MB (2021-12-23). "Антимикробная активность нейтрофилов против микобактерий". Frontiers in Immunology . 12 : 782495. doi : 10.3389/fimmu.2021.782495 . PMC 8732375. PMID 35003097 .
^ Chu HT, Lin H, Tsao TT, Chang CF, Hsiao WW, Yeh TJ и др. (сентябрь 2013 г.). «Генотипирование антигенов нейтрофилов человека (HNA) по данным секвенирования всего генома». BMC Medical Genomics . 6 (1): 31. doi : 10.1186/1755-8794-6-31 . PMC 3849977 . PMID 24028078. В данной статье используется текст, доступный по лицензии CC BY 2.0.
^ ab Игнатов Д.Ю. (2012). Функциональная гетерогенность нейтрофилов человека и их роль в регуляции количества лейкоцитов периферической крови (канд. мед. наук). Донецкий национальный медицинский университет. doi :10.13140/RG.2.2.35542.34884.
^ Герасимов ИГ, Игнатов ДИ (2001). "[Функциональная гетерогенность нейтрофилов крови человека: генерация активных форм кислорода]". Цитология . 43 (5): 432–436. PMID 11517658.
^ Герасимов ИГ, Игнатов ДИ (2004). «[Активация нейтрофилов in vitro]». Цитология . 46 (2): 155–158. ПМИД 15174354.
^ Герасимов ИГ, Игнатов ДИ, Котельницкий МА (2005). "[Восстановление нитросинего тетразолия нейтрофилами крови человека. I. Влияние pH]". Цитология . 47 (6): 549–553. PMID 16708848.
^ Герасимов ИГ, Игнатов ДИ (2005). "[Восстановление нитросинего тетразолия нейтрофилами крови человека. II. Влияние ионов натрия и калия]". Цитология . 47 (6): 554–558. PMID 16708849.
^ Zilionis R, Engblom C, Pfirschke C, Savova V, Zemmour D, Saatcioglu HD и др. (май 2019 г.). «Транскриптомика отдельных клеток рака легких человека и мыши выявляет консервативные популяции миелоидных клеток у разных особей и видов». Immunity . 50 (5): 1317–1334.e10. doi :10.1016/j.immuni.2019.03.009. PMC 6620049 . PMID 30979687.
^ Graham DB, Zinselmeyer BH, Mascarenhas F, Delgado R, Miller MJ, Swat W (2009). Unutmaz D (ред.). "ITAM signaling by Vav family Rho guanine nucleotide exchange factors adjustable interstitial transit rates of neutrophils in vivo". PLOS ONE . 4 (2): e4652. Bibcode :2009PLoSO...4.4652G. doi : 10.1371/journal.pone.0004652 . PMC 2645696 . PMID 19247495.
Внешние ссылки
Информация о нейтропении, заархивированная 2015-12-02 на Wayback Machine
Калькулятор абсолютного количества нейтрофилов
Содержание и распределение микроэлементов в нейтрофилах






_diagram_en.svg/1280px-Hematopoiesis_(human)_diagram_en.svg.png)
![Нейтрофилы видны при инфаркте миокарда примерно через 12–24 часа[64], как показано на этой микрофотографии.](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg/1280px-Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg)
![При инсульте они начинают проникать в пораженный инфарктом мозг через 6–8 часов.[65]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1e/Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg/1280px-Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg)
