Plesiosauria [a] [4] или плезиозавры — отряд или клада вымерших мезозойских морских рептилий , принадлежащих к Sauropterygiaм .
Плезиозавры впервые появились в конце триасового периода , возможно, в ретийском ярусе, около 203 миллионов лет назад. [5] Они стали особенно распространены в юрский период, процветая до своего исчезновения из-за мел-палеогенового вымирания в конце мелового периода, около 66 миллионов лет назад. Они имели всемирное океаническое распространение, и некоторые виды, по крайней мере частично, населяли пресноводные среды. [6]
Плезиозавры были одними из первых обнаруженных ископаемых рептилий. В начале девятнадцатого века ученые поняли, насколько отличительным было их строение, и в 1835 году они были названы отдельным отрядом. Первый род плезиозавров, одноименный Plesiosaurus , был назван в 1821 году. С тех пор было описано более ста действительных видов. В начале двадцать первого века число открытий возросло, что привело к улучшению понимания их анатомии, взаимоотношений и образа жизни.
Плезиозавры имели широкое плоское тело и короткий хвост. Их конечности эволюционировали в четыре длинных ласта, которые приводились в действие сильными мышцами, прикрепленными к широким костным пластинам, образованным плечевым поясом и тазом. Ласты совершали летательные движения в воде. Плезиозавры дышали воздухом и рождали живых детенышей; есть указания на то, что они были теплокровными.
Плезиозавры показали два основных морфологических типа. Некоторые виды с строением «плезиозавроморф» имели (иногда чрезвычайно) длинные шеи и маленькие головы; они были относительно медлительными и ловили мелких морских животных. Другие виды, некоторые из которых достигали длины до семнадцати метров, имели строение «плиозавроморф» с короткой шеей и большой головой; они были высшими хищниками , быстрыми охотниками на крупную добычу. Два типа связаны с традиционным строгим разделением плезиозавров на два подотряда, длинношеих Plesiosauroidea и короткошеих Pliosauroidea . Однако современные исследования показывают, что несколько групп «с длинной шеей» могли иметь некоторых членов с короткой шеей или наоборот. Поэтому были введены чисто описательные термины «плезиозавроморф» и «плиозавроморф», которые не подразумевают прямой связи. «Plesiosauroidea» и «Pliosauroidea» сегодня имеют более ограниченное значение. Термин «плезиозавр» правильно использовать для обозначения всего семейства плезиозавров, но неформально его иногда используют только для обозначения длинношеих форм — древних плезиозавроидов.
Как и другие древние морские рептилии, такие как представители кладов Ichthyosauria и Mosasauria , роды Plesiosauria не являются частью клада Dinosauria . [7]

Элементы скелета плезиозавров являются одними из первых ископаемых вымерших рептилий, признанных таковыми. [8] В 1605 году Ричард Верстеген из Антверпена проиллюстрировал в своей работе «Восстановление угасшего интеллекта» позвонки плезиозавров, которые он отнес к рыбам и считал доказательством того, что Великобритания когда-то была связана с европейским континентом. [9] Валлиец Эдвард Ллойд в своей работе «Lithophylacii Brittannici Ichnographia» от 1699 года также включил изображения позвонков плезиозавров, которые снова считались позвонками рыб или ихтиоспондили . [10] Другие натуралисты в семнадцатом веке добавили останки плезиозавров в свои коллекции, например, Джон Вудворд ; только гораздо позже они были признаны плезиозавровыми по своей природе и сегодня частично хранятся в Музее Седжвика . [8]
В 1719 году Уильям Стьюкли описал частичный скелет плезиозавра, на который его внимание обратил прадед Чарльза Дарвина , Роберт Дарвин из Элстона . Каменная пластина была добыта в карьере в Фулбеке в Линкольншире и использовалась с ископаемым на нижней стороне для укрепления склона водопоя в Элстоне в Ноттингемшире . После того, как были обнаружены странные кости, которые она содержала, она была выставлена в местном викарии как останки грешника, утонувшего во время Великого потопа . Стьюкли подтвердил ее « дилювиальную » природу, но понял, что она представляла собой некое морское существо, возможно, крокодила или дельфина. [11] Образец сегодня экспонируется в Музее естественной истории , и его инвентарный номер — NHMUK PV R.1330 (ранее BMNH R.1330). Это самый ранний обнаруженный более или менее полный ископаемый скелет рептилии в музейной коллекции. Вероятно, его можно отнести к Plesiosaurus dolichodeirus . [8]

В течение восемнадцатого века число открытий английских плезиозавров быстро увеличивалось, хотя все они носили более или менее фрагментарный характер. Важными коллекционерами были преподобные Уильям Маунси и баптист Ноэль Тернер, работавшие в долине Бельвуар , чьи коллекции в 1795 году были описаны Джоном Николсом в первой части его «Истории и древностей графства Лестершир» . [12] Один из частичных скелетов плезиозавров Тернера до сих пор хранится как образец NHMUK PV R.45 (ранее BMNH R.45) в Британском музее естественной истории; сегодня он называется Thalassiodracon . [8]

В начале девятнадцатого века плезиозавры были еще плохо известны, и их особое строение не было понято. Систематического различия между ихтиозаврами не проводилось , поэтому окаменелости одной группы иногда объединяли с окаменелостями другой, чтобы получить более полный образец. В 1821 году частичный скелет, обнаруженный в коллекции полковника Томаса Джеймса Бирча, [13] был описан Уильямом Конибером и Генри Томасом Де ла Бешем и признан представителем особой группы. Был назван новый род Plesiosaurus . Родовое название произошло от греческого πλήσιος, plèsios , «ближе к» и латинизированного saurus , в значении «ящер», чтобы выразить, что Plesiosaurus был в Цепи Бытия более близко расположен к Sauria , в частности к крокодилу, чем Ichthyosaurus , который имел форму более низшей рыбы. [14] Таким образом, название следует читать скорее как «приближающийся к Sauria» или «почти рептилия», чем как «почти ящерица». [15] Части образца все еще присутствуют в Музее естественной истории Оксфордского университета . [8]
Вскоре после этого морфология стала гораздо более известной. В 1823 году Томас Кларк сообщил о почти полном черепе, вероятно, принадлежащем Thalassiodracon , который сейчас хранится в Британской геологической службе как образец BGS GSM 26035. [8] В том же году коммерческий коллекционер окаменелостей Мэри Эннинг и ее семья обнаружили почти полный скелет в Лайм-Реджисе в Дорсете , Англия, на том, что сегодня называется Юрским побережьем . Его приобрел герцог Бекингем , который предоставил его геологу Уильяму Бакленду . Он, в свою очередь, позволил Кониберу описать его 20 февраля 1824 года в докладе, прочитанном в Геологическом обществе Лондона , [16] во время того же заседания, на котором впервые был назван динозавр — Megalosaurus . Две находки выявили уникальное и причудливое строение животных, которое в 1832 году профессор Бакленд сравнил с «морской змеей, пронзившей черепаху». В 1824 году Конибер также дал специфическое название плезиозавру : dolichodeirus , что означает «длинношеий». В 1848 году скелет был куплен Британским музеем естественной истории и каталогизирован как образец NHMUK OR 22656 (ранее BMNH 22656). [8] Когда лекция была опубликована , Конибер также назвал второй вид: Plesiosaurus giganteus . Это была короткошеяя форма, позже отнесенная к Pliosauroidea . [17]

Плезиозавры стали более известны широкой публике благодаря двум богато иллюстрированным публикациям коллекционера Томаса Хокинса : «Мемуары ихтиозавров и плезиозавров» 1834 года [18] и «Книга великих морских драконов» 1840 года. Хокинс придерживался весьма своеобразного взгляда на животных, [19] считая их чудовищными созданиями дьявола в доадамитовскую фазу истории. [20] В конце концов Хокинс продал свои ценные и красиво отреставрированные образцы Британскому музею естественной истории. [21]
В течение первой половины девятнадцатого века число находок плезиозавров неуклонно росло, особенно за счет открытий в морских скалах Лайм-Реджиса. Только сэр Ричард Оуэн назвал около сотни новых видов. Однако большинство их описаний были основаны на изолированных костях, без достаточной диагностики, чтобы иметь возможность отличить их от других видов, которые были описаны ранее. Многие из новых видов, описанных в это время, впоследствии были признаны недействительными . Род Plesiosaurus особенно проблематичен, поскольку большинство новых видов были помещены в него так, что он стал таксоном для мусора . Постепенно были названы другие роды. Хокинс уже создал новые роды, хотя они больше не считаются действительными. В 1841 году Оуэн назвал Pliosaurus brachydeirus . Его этимология ссылается на более раннего Plesiosaurus dolichodeirus , поскольку он происходит от πλεῖος, pleios , «более полно», отражая то, что, согласно Оуэну, он был ближе к Sauria, чем Plesiosaurus . Его видовое название означает «с короткой шеей». [22] Позже было признано, что Pliosauridae имеют морфологию, принципиально отличную от плезиозавридов. Семейство Plesiosauridae уже было придумано Джоном Эдвардом Греем в 1825 году . [23] В 1835 году Анри Мари Дюкроте де Бленвиль назвал сам отряд Plesiosauria. [24]
Во второй половине девятнадцатого века важные находки были сделаны за пределами Англии. Хотя сюда входили и некоторые немецкие открытия, в основном это были плезиозавры, найденные в отложениях американского мелового Западного Внутреннего морского пути , Ниобрарского мела . Одна окаменелость в частности ознаменовала начало Войн за кости между соперничающими палеонтологами Эдвардом Дринкером Коупом и Отниелом Чарльзом Маршем .

В 1867 году врач Теофил Тернер около Форт-Уоллеса в Канзасе обнаружил скелет плезиозавра, который он подарил Коупу. [25] Коуп попытался реконструировать животное, предполагая, что более длинная часть позвоночника была хвостом, более короткая — шеей. Вскоре он заметил, что скелет, формировавшийся под его руками, обладал некоторыми совершенно особыми качествами: шейные позвонки имели шевроны, а суставные поверхности хвостовых позвонков были ориентированы сзади наперед. [26] Взволнованный, Коуп пришел к выводу, что открыл совершенно новую группу рептилий: Streptosauria или «перевернутые ящеры», которые отличались бы перевернутыми позвонками и отсутствием задних конечностей, а хвост обеспечивал основное движение. [27] После публикации описания этого животного [28] с последующей иллюстрацией в учебнике по рептилиям и амфибиям [29] Коуп пригласил Марша и Джозефа Лейди полюбоваться его новым Elasmosaurus platyurus . Послушав некоторое время интерпретацию Коупа, Марш предположил, что более простым объяснением странного строения было бы то, что Коуп перевернул позвоночный столб относительно тела в целом. Когда Коуп с возмущением отреагировал на это предложение, Лейди молча взял череп и поместил его напротив предполагаемого последнего хвостового позвонка, к которому он идеально подходил: на самом деле это был первый шейный позвонок, к которому все еще был прикреплен кусок заднего черепа. [30] Униженный, Коуп попытался уничтожить все издание учебника и, когда это не удалось, немедленно опубликовал улучшенное издание с правильной иллюстрацией, но с той же датой публикации. [31] Он оправдывал свою ошибку, утверждая, что его ввел в заблуждение сам Лейди, который, описывая образец цимолиазавра , также перевернул позвоночный столб. [32] Марш позже утверждал, что это дело было причиной его соперничества с Коупом: «с тех пор он стал моим злейшим врагом». И Коуп, и Марш в своем соперничестве назвали много родов и видов плезиозавров, большинство из которых сегодня считаются недействительными. [33]
На рубеже веков большинство исследований плезиозавров было проведено бывшим студентом Марша, профессором Сэмюэлем Уэнделлом Уиллистоном . В 1914 году Уиллистоном была опубликована его работа «Водные рептилии прошлого и настоящего» . [34] Несмотря на то, что в ней рассматривались морские рептилии в целом, она на протяжении многих лет оставалась самым обширным общим текстом по плезиозаврам. [35] В 2013 году Оливье Риппель подготовил первый современный учебник. В середине двадцатого века США оставались важным центром исследований, в основном благодаря открытиям Сэмюэля Пола Уэллса .
В то время как в течение девятнадцатого и большей части двадцатого века новые плезиозавры описывались со скоростью три или четыре новых рода каждое десятилетие, темп внезапно ускорился в 1990-х годах, и за этот период было обнаружено семнадцать плезиозавров. Темпы открытий ускорились в начале двадцать первого века, и каждый год называлось около трех или четырех плезиозавров. [36] Это означает, что около половины известных плезиозавров являются относительно новыми для науки, что является результатом гораздо более интенсивных полевых исследований. Часть из них происходит вдали от традиционных областей, например, на новых участках, разработанных в Новой Зеландии , Аргентине , Чили , [37] Норвегии , Японии , Китае и Марокко , но места более оригинальных открытий оказались по-прежнему продуктивными, с важными новыми находками в Англии и Германии. Некоторые из новых родов являются переименованиями уже известных видов, которые были сочтены достаточно разными, чтобы оправдать отдельное название рода.
В 2002 году прессе было объявлено о « Монстре Арамберри ». Обнаруженный в 1982 году в деревне Арамберри , в северном мексиканском штате Нуэво-Леон , он изначально был классифицирован как динозавр . Образец на самом деле является очень большим плезиозавром, возможно, достигающим 15 м (49 футов) в длину. Средства массовой информации опубликовали преувеличенные сообщения, утверждая, что он был 25 метров (82 фута) в длину и весил до 150 000 килограммов (330 000 фунтов), что сделало бы его одним из крупнейших хищников всех времен. [38] [39]
В 2004 году в национальном природном заповеднике Бриджуотер-Бей в Сомерсете, Великобритания, местный рыбак обнаружил то, что, по всей видимости, было совершенно неповрежденным молодым плезиозавром . Окаменелость, возраст которой, как показывают аммониты , составляет 180 миллионов лет , достигала 1,5 метра (4 фута 11 дюймов) в длину и может быть связана с ромалеозавром . Вероятно, это наиболее хорошо сохранившийся образец плезиозавра, когда-либо обнаруженный. [40] [41] [42]
В 2005 году останки трех плезиозавров ( Dolichorhynchops herschelensis ), обнаруженные в 1990-х годах недалеко от Гершеля, Саскачеван, были признаны новым видом доктором Тамаки Сато, японским палеонтологом позвоночных. [43]
В 2006 году объединенная группа американских и аргентинских исследователей (последние из Аргентинского антарктического института и Музея Ла-Платы ) обнаружила скелет молодого плезиозавра длиной 1,5 метра (4 фута 11 дюймов) на острове Вега в Антарктиде. [44] В настоящее время окаменелость экспонируется в геологическом музее Школы горного дела и технологий Южной Дакоты . [45]
В 2008 году на Шпицбергене были обнаружены ископаемые останки неописанного плезиозавра, названного Predator X , ныне известного как Pliosaurus funkei . [46] Он имел длину 12 м (39 футов), а сила его укуса в 149 килоньютонов (33 000 фунтов силы ) является одной из самых мощных из известных. [47]
В декабре 2017 года на континенте Антарктида был найден большой скелет плезиозавра, самого древнего существа на континенте и первого представителя своего вида в Антарктиде. [48]
Не только увеличилось количество полевых открытий, но и с 1950-х годов плезиозавры стали предметом более обширной теоретической работы. Методология кладистики впервые позволила точно рассчитать их эволюционные связи. Было опубликовано несколько гипотез о том, как они охотились и плавали, включающих общие современные идеи о биомеханике и экологии . Многочисленные недавние открытия проверили эти гипотезы и дали начало новым. [ оригинальное исследование? ]

Плезиозавры происходят от Sauropterygia , группы, возможно, архелозавровых рептилий, которые вернулись в море. Продвинутая подгруппа завроптеригий, плотоядные Eusauropterygia с маленькими головами и длинными шеями, разделилась на две ветви во время верхнего триаса . Одна из них, Nothosauroidea , сохранила функциональные локтевые и коленные суставы; но другая, Pistosauria , стала более полно приспособленной к образу жизни в море. Их позвоночник стал жестче, и основная движущая сила во время плавания больше не исходила от хвоста, а от конечностей, которые превратились в ласты. [49] Pistosauria стали теплокровными и живородящими , рождая живых детенышей. [50] Ранние, базальные , члены группы, традиционно называемые « пистозавридами », все еще были в основном прибрежными животными. Их плечевые пояса оставались слабыми, их таз не мог выдерживать силу сильного гребка, а их ласты были тупыми. Позже откололась более продвинутая группа пистозавров: плезиозавры. У них были усиленные плечевые пояса, более плоский таз и более заостренные ласты. Другие адаптации, позволившие им колонизировать открытые моря, включали жесткие суставы конечностей; увеличение числа фаланг рук и ног; более плотное боковое соединение фаланг пальцев рук и ног и укороченный хвост. [51] [52]

Начиная с самого раннего юрского периода , геттангского яруса, известна богатая радиация плезиозавров, что подразумевает, что группа уже должна была диверсифицироваться в позднем триасе ; однако из этой диверсификации было обнаружено только несколько (очень) базальных форм, наиболее производный Rhaeticosaurus . Последующая эволюция плезиозавров весьма спорна. Различные кладистические анализы не привели к консенсусу относительно взаимоотношений между основными подгруппами плезиозавров. Традиционно плезиозавры делились на длинношеих Plesiosauroidea и короткошеих Pliosauroidea . Однако современные исследования показывают, что некоторые обычно длинношеие группы могли иметь короткошеих членов. Чтобы избежать путаницы между филогенией , эволюционными связями и морфологией , то есть строением животного, формы с длинной шеей называются «плезиозавроморфными», а формы с короткой шеей — «плиозавроморфными», при этом виды «плезиозавроморфных» не обязательно должны быть более тесно связаны друг с другом, чем с формами «плиозавроморфных». [53]

Последний общий предок Plesiosauria, вероятно, был довольно маленькой формой с короткой шеей. В раннем юрском периоде подгруппой с наибольшим количеством видов была Rhomaleosauridae , возможно, очень базальное ответвление видов, которые также были с короткой шеей. Плезиозавры в этот период были не более пяти метров (шестнадцать футов) в длину. К тоару , около 180 миллионов лет назад, другие группы, среди которых Plesiosauridae , стали более многочисленными, и некоторые виды развили более длинные шеи, в результате чего общая длина тела достигала десяти метров (33 фута). [54]
В середине юрского периода появились очень крупные Pliosauridae . Они характеризовались большой головой и короткой шеей, например, Liopleurodon и Simolestes . Эти формы имели черепа длиной до трех метров (десять футов), достигали длины до семнадцати метров (56 футов) и веса в десять тонн. У Pliosauridae были большие конические зубы, и они были доминирующими морскими плотоядными животными своего времени. В то же время, примерно 160 миллионов лет назад, присутствовали Cryptoclididae , более короткие виды с длинной шеей и маленькой головой. [55]
Leptocleididae распространились в течение раннего мелового периода . Это были довольно мелкие формы, которые, несмотря на свои короткие шеи, могли быть более тесно связаны с Plesiosauridae, чем с Pliosauridae. Позже в раннем меловом периоде появились Elasmosauridae ; они были одними из самых длинных плезиозавров, достигая до пятнадцати метров (пятидесяти футов) в длину из-за очень длинных шей, содержащих до 76 позвонков, больше, чем у любого другого известного позвоночного. Pliosauridae все еще присутствовали, как показывают крупные хищники, такие как Kronosaurus . [55]
В начале позднего мела ихтиозавры вымерли; возможно, группа плезиозавров эволюционировала, чтобы заполнить их ниши: Polycotylidae , у которых были короткие шеи и необычайно удлиненные головы с узкими мордами. В течение позднего мела эласмозавриды все еще имели много видов. [55]
Все плезиозавры вымерли в результате КТ-события в конце мелового периода, примерно 66 миллионов лет назад. [56]
В современной филогении клады — это определенные группы, которые содержат все виды, принадлежащие к определенной ветви эволюционного дерева . Один из способов определения клады — позволить ей состоять из последнего общего предка двух таких видов и всех его потомков. Такая клада называется « узловой кладой ». В 2008 году Патрик Дракенмиллер и Энтони Рассел таким образом определили Plesiosauria как группу, состоящую из последнего общего предка Plesiosaurus dolichocheirus и Peloneustes philarchus и всех его потомков. [57] Plesiosaurus и Peloneustes представляли собой основные подгруппы Plesiosauroidea и Pliosauroidea и были выбраны по историческим причинам; любой другой вид из этих групп был бы достаточен.
Другой способ определить кладу — позволить ей состоять из всех видов, более тесно связанных с определенным видом, который в любом случае хотят включить в кладу, чем с другим видом, который, наоборот, хотят исключить. Такая клада называется « стволовой кладой ». Такое определение имеет то преимущество, что в нее легче включить все виды с определенной морфологией . В 2010 году Хиллари Кетчум и Роджер Бенсон определили Plesiosauria как такой таксон на основе стебля : «все таксоны, более тесно связанные с Plesiosaurus dolichodeirus и Pliosaurus brachydeirus , чем с Augustasaurus hagdorni ». Кетчум и Бенсон (2010) также ввели новую кладу Neoplesiosauria, таксон на основе узла , который был определен как « Plesiosaurus dolichodeirus , Pliosaurus brachydeirus , их самый последний общий предок и все его потомки». [55] Клада Neoplesiosauria, скорее всего, материально идентична Plesiosauria sensu Druckenmiller & Russell, поэтому она будет обозначать тот же самый вид, и этот термин должен был заменить эту концепцию.
Бенсон и др. (2012) обнаружили, что традиционные Pliosauroidea парафилетичны по отношению к Plesiosauroidea. Было обнаружено, что Rhomaleosauridae находятся за пределами Neoplesiosauria, но все еще внутри Plesiosauria. Ранний карнийский пистозавр Bobosaurus оказался на один шаг более продвинутым, чем Augustasaurus, по отношению к Plesiosauria, и поэтому он по определению представлял собой самого базального известного плезиозавра. Этот анализ был сосредоточен на базальных плезиозаврах, и поэтому были включены только один производный плиозавр и один криптоклидий , в то время как эласмозавриды не были включены вообще. Более подробный анализ, опубликованный как Бенсоном, так и Дракенмиллером в 2014 году, не смог разрешить отношения между линиями в основе Plesiosauria. [58]

Следующая кладограмма следует анализу Бенсона и Дракенмиллера (2014). [58]


В целом, длина взрослой особи плезиозавров варьировалась от 1,5 метров (4,9 футов) до примерно 15 метров (49 футов). Таким образом, группа включала некоторых из крупнейших морских хищников в палеонтологической летописи , примерно равных по размеру самым длинным ихтиозаврам , мозазавридам , акулам и зубатым китам . Некоторые останки плезиозавров, такие как набор длиной 2,875 метра (9,43 фута) хорошо реконструированных и фрагментарных нижних челюстей, хранящихся в Музее Оксфордского университета и относящихся к Pliosaurus rossicus (ранее относимых к Stretosaurus [59] и Liopleurodon ), указали длину 17 метров (56 футов). Однако недавно было высказано мнение, что его размер в настоящее время не может быть определен из-за их плохой реконструкции, и более вероятной является длина 12,7 метра (42 фута) или меньше. [60] MCZ 1285, образец, в настоящее время относимый к Kronosaurus queenslandicus , из раннего мела Австралии , по оценкам имел длину черепа 2,21–2,85 м (7,3–9,4 фута). [60] [61] Серия шейных позвонков из глиняной формации Киммеридж указывает на плиозавра, вероятно, Pliosaurus , который мог быть длиной до 14,4 метра (47 футов). [62]
Типичный плезиозавр имел широкое, плоское тело и короткий хвост . Плезиозавры сохранили свои предковые две пары конечностей, которые эволюционировали в большие ласты . [63] Плезиозавры были связаны с более ранними Nothosauridae , [64] которые имели более крокодилоподобное тело. Расположение ласт необычно для водных животных, поскольку, вероятно, все четыре конечности использовались для продвижения животного по воде движениями вверх и вниз. Хвост, скорее всего, использовался только для помощи в управлении направлением. Это контрастирует с ихтиозаврами и более поздними мозазаврами , у которых хвост обеспечивал основное движение. [65]
Для приведения в действие ласт плечевой пояс и таз были значительно модифицированы, превратившись в широкие костные пластины на нижней стороне тела, которые служили поверхностью прикрепления для больших групп мышц, способных тянуть конечности вниз. В плече коракоид стал самым крупным элементом, покрывающим большую часть груди. Лопатка была намного меньше, образуя внешний передний край туловища. К середине она продолжалась в ключицу и, наконец, в небольшую межключичную кость. Как и у большинства четвероногих , плечевой сустав был образован лопаткой и коракоидом. В тазу костная пластина была образована седалищной костью сзади и более крупной лобковой костью спереди нее. Подвздошная кость , которая у наземных позвоночных несет вес задней конечности, стала небольшим элементом сзади, больше не прикрепленным ни к лобковой, ни к бедренной кости. Тазобедренный сустав был образован седалищной костью и лобковой костью. Грудные и тазовые пластины были соединены пластроном , костной клеткой, образованной парными брюшными ребрами , каждое из которых имело среднюю и внешнюю часть. Такое расположение обездвиживало все туловище. [65]
Чтобы стать ластами, конечности значительно изменились. Конечности были очень большими, каждая примерно такой же длины, как туловище. Передние и задние конечности сильно напоминали друг друга. Плечевая кость в верхней части руки и бедренная кость в верхней части ноги стали большими плоскими костями, расширенными на своих внешних концах. Локтевые и коленные суставы больше не функционировали: нижняя часть руки и голень не могли сгибаться относительно элементов верхней конечности, а образовывали плоское продолжение их. Все внешние кости стали плоскими опорными элементами ласт, плотно соединенными друг с другом и едва способными вращаться, сгибаться, разгибаться или расправляться. Это касалось локтевой кости , лучевой кости , пястных костей и пальцев, а также большеберцовой кости , малоберцовой кости , плюсневых костей и пальцев ног. Кроме того, для удлинения ласт число фаланг увеличилось до восемнадцати в ряду, явление, называемое гиперфалангией . Ласты не были идеально плоскими, а имели слегка выпуклый изогнутый верхний профиль, как у аэродинамического профиля , чтобы иметь возможность «летать» в воде. [65]

В то время как плезиозавры мало различались по строению туловища и в этом отношении могут быть названы «консервативными», между подгруппами имелись значительные различия в форме шеи и черепа. Плезиозавров можно разделить на два основных морфологических типа, которые различаются по размеру головы и шеи . «Плезиозавроморфы», такие как Cryptoclididae , Elasmosauridae и Plesiosauridae , имели длинные шеи и маленькие головы. «Плиозавроморфы», такие как Pliosauridae и Rhomaleosauridae , имели более короткие шеи с большой удлиненной головой. Изменения длины шеи были вызваны не удлинением отдельных шейных позвонков, а увеличением их количества. У Elasmosaurus семьдесят два шейных позвонка; известный рекорд принадлежит эласмозавриду Albertonectes с семьюдесятью шестью шейными позвонками. [66] Большое количество суставов навело ранних исследователей на мысль, что шея должна была быть очень гибкой; действительно, предполагалось, что изгиб шеи, подобный лебедю, возможен - на исландском языке плезиозавры даже называются Svaneðlur , «лебединые ящеры». Однако современные исследования подтвердили более раннюю гипотезу Уиллистона о том, что длинные пластинчатые шипы на вершине позвонков, processus spinosi , сильно ограничивали вертикальное движение шеи. Хотя горизонтальное изгибание было менее ограничено, в целом шея должна была быть довольно жесткой и, безусловно, не могла сгибаться в змеевидные кольца. Это еще более верно в отношении короткошеих «плиозавромофов», у которых было всего одиннадцать шейных позвонков. У ранних форм амфицельные или амфиплатные шейные позвонки несли двуглавые шейные ребра; у более поздних форм ребра были одноглавыми. В остальной части позвоночника число спинных позвонков варьировалось от девятнадцати до тридцати двух; крестцовых позвонков — от двух до шести, а хвостовых позвонков — от двадцати одного до тридцати двух. Эти позвонки все еще обладали исходными отростками, унаследованными от наземных предков Sauropterygia, и не были редуцированы до рыбоподобных простых дисков, как это произошло с позвонками ихтиозавров. Хвостовые позвонки обладали шевронными костями. Спинные позвонки плезиозавров легко узнаваемы по двум большим субцентральным отверстиям , парным сосудистым отверстиям на нижней стороне. [65]
Череп плезиозавров показал « эвриапсидное » состояние, не имея нижних височных окон , отверстий на нижних задних сторонах. Верхние височные окна образовывали большие отверстия по бокам задней крыши черепа, место прикрепления мышц, закрывающих нижние челюсти. Как правило, теменные кости были очень большими, с гребнем по средней линии, в то время как чешуйчатые кости обычно образовывали дугу, исключая теменные кости из затылка . Глазницы были большими, в целом направленными косо вверх; у плиозавридов глаза были более направлены вбок. Глаза поддерживались склеральными кольцами , форма которых показывает, что они были относительно плоскими, что было адаптацией к нырянию. Расположенные спереди внутренние ноздри, хоаны , имеют небные бороздки для направления воды, поток которой поддерживался гидродинамическим давлением над расположенными сзади, перед глазницами, внешними ноздрями во время плавания. Согласно одной из гипотез, во время прохождения через носовые протоки вода «обонялась» обонятельным эпителием. [67] [68] Однако, ближе к задней части, в нёбе присутствует вторая пара отверстий; более поздняя гипотеза утверждает, что это настоящие хоаны , а передняя пара на самом деле представляла собой парные солевые железы . [69] Расстояние между глазницами и ноздрями было настолько ограничено, потому что носовые кости были сильно редуцированы, даже отсутствовали у многих видов. Предчелюстные кости непосредственно касались лобных костей ; у эласмозавридов они даже доходили до теменных костей . Часто также отсутствовали слезные кости . [52]

Форма и количество зубов были очень изменчивы. Некоторые формы имели сотни игловидных зубов. Большинство видов имели более крупные конические зубы с круглым или овальным поперечным сечением. Таких зубов было от четырех до шести в предчелюстной кости и около четырнадцати-двадцати пяти в верхней челюсти ; количество в нижней челюсти примерно равнялось количеству в черепе. Зубы располагались в зубных гнездах, имели вертикально морщинистую эмаль и не имели настоящего режущего края или киля . У некоторых видов передние зубы были заметно длиннее, чтобы хватать добычу. [70]
Остатки мягких тканей плезиозавров редки, но иногда, особенно в сланцевых отложениях, они частично сохранились, например, показывая очертания тела. Ранним открытием в этом отношении был голотип Plesiosaurus conybeari (в настоящее время Attenborosaurus ). Из таких находок известно, что кожа была гладкой, без видимых чешуек, но с небольшими морщинами, что задняя кромка ласт значительно выступала за кости конечностей; [71] и что хвост имел вертикальный плавник, как сообщал Вильгельм Деймс в своем описании Plesiosaurus guilelmiimperatoris (в настоящее время Seeleyosaurus ). [72] Возможность хвостового плавника была подтверждена недавними исследованиями формы хвостового неврального позвоночника Pantosaurus , Cryptoclidus и Rhomaleosaurus zetlandicus . [73] [74] [75] Исследование 2020 года утверждает, что хвостовой плавник имел горизонтальную конфигурацию. [76]


Вероятный источник пищи плезиозавров варьировался в зависимости от того, принадлежали ли они к длинношеим «плезиозавроморфным» формам или к короткошеим «плиозавроморфным» видам.
Чрезвычайно длинные шеи «плезиозавроморфов» стали причиной спекуляций относительно их функции с того самого момента, как стало очевидным их особое строение. Конибер предложил три возможных объяснения. Шея могла служить для перехвата быстро движущейся рыбы во время преследования. В качестве альтернативы плезиозавры могли отдыхать на дне моря, в то время как голова была отправлена на поиски добычи, что, казалось, подтверждалось тем фактом, что глаза были направлены относительно вверх. Наконец, Конибер предположил возможность того, что плезиозавры плавали на поверхности, позволяя своим шеям нырять вниз, чтобы искать пищу на более низких уровнях. Все эти интерпретации предполагали, что шея была очень гибкой. Современное понимание того, что шея была на самом деле довольно жесткой, с ограниченным вертикальным движением, потребовало новых объяснений. Одна из гипотез заключается в том, что длина шеи позволяла застигать врасплох косяки рыб, голова прибывала до того, как вид или волна давления туловища могли предупредить их. «Плезиозавроморфы» охотились визуально, о чем свидетельствуют их большие глаза, и, возможно, использовали направленное чувство обоняния. Твердые и мягкотелые головоногие моллюски, вероятно, составляли часть их рациона. Их челюсти , вероятно, были достаточно сильными, чтобы прокусывать твердые панцири этого типа добычи. Были найдены ископаемые образцы с панцирями головоногих моллюсков, все еще находящимися в их желудках. [77] Костистые рыбы ( Osteichthyes ), которые еще больше разнообразились в юрский период, вероятно, также были добычей. Совершенно другая гипотеза утверждает, что «плезиозавроморфы» были донными питающимися. Жесткие шеи использовались для пропахивания морского дна, поедая бентос . Это было бы доказано длинными бороздами, присутствующими на древнем морском дне. [78] [79] Такой образ жизни в 2017 году был предложен для Morturneria . [80] «Плезиозавроморфы» не были хорошо приспособлены к ловле крупной быстро движущейся добычи, так как их длинные шеи, хотя и кажутся обтекаемыми, вызывали огромное трение кожи . Санкар Чаттерджи предположил в 1989 году, что некоторые Cryptocleididae были суспензионными питателями, фильтрующими планктон . Например, у Aristonectes были сотни зубов, что позволяло ему отсеивать мелких ракообразных из воды. [81]
Короткошеие «плиозавроморфы» были верхними плотоядными животными, или хищниками высшего порядка , в своих пищевых цепях . [82] Они были хищниками-преследователями [83] или хищниками-засадчиками, охотящимися на добычу разного размера, и оппортунистическими едоками; их зубы могли использоваться для прокалывания мягкотелой добычи, особенно рыбы. [84] Их головы и зубы были очень большими, подходящими для того, чтобы хватать и разрывать крупных животных. Их морфология позволяла им плавать с большой скоростью. Они также охотились визуально.
Плезиозавры сами были добычей других плотоядных животных, о чем свидетельствуют следы укусов, оставленные акулой, которые были обнаружены на окаменелом плавнике плезиозавра [85] , а также окаменелые останки содержимого желудка мозазавра, которые, как полагают, являются останками плезиозавра. [86]
Также были обнаружены скелеты с гастролитами , камнями, в желудках, хотя не было установлено, помогали ли они переваривать пищу, особенно головоногих, в мускулистом желудке или изменяли плавучесть , или и то, и другое. [87] [88] Однако общий вес гастролитов, обнаруженных в различных образцах, по-видимому, недостаточен для изменения плавучести этих крупных рептилий. [89] Первые гастролиты плезиозавров, обнаруженные у Mauisaurus gardneri (a nomen nudum [90] ), были зарегистрированы Гарри Гоувером Сили в 1877 году. [91] Количество этих камней на особь часто очень велико. В 1949 году ископаемое Alzadasaurus (образец SDSM 451, позже переименованный в Styxosaurus ) показало 253 из них. [92] Размер отдельных камней часто бывает значительным. В 1991 году был исследован образец эласмозаврида, KUVP 129744, содержащий гастролит диаметром семнадцать сантиметров и весом 1300 граммов; и несколько более короткий камень весом 1490 граммов. Всего было обнаружено сорок семь гастролитов общим весом 13 килограммов. Размер камней рассматривался как указание на то, что они были проглочены не случайно, а преднамеренно, животное, возможно, преодолело большие расстояния в поисках подходящего типа породы. [93] Типовой образец Scalamagnus (MNA V10046) связан с 289 гастролитами , что необычно по сравнению с большинством скелетов поликотилид, у которых гастролиты обычно отсутствуют. Общая масса гастролитов составляла от менее 0,1 грамма до 18,5 грамма, а общая масса составляла около 518 граммов. Около трех четвертей камней весили менее 2 граммов, при этом средняя масса и медианная масса камней соответственно оценивались в 1,9 грамма и 0,8 грамма. Гастролиты имели высокое среднее значение и изменчивость сферичности, что позволяет предположить, что этот человек получал свои камни из рек, расположенных вдоль западной стороны Западного внутреннего морского пути. [94]
Отличительная форма тела с четырьмя плавниками вызвала немало спекуляций о том, какой тип гребка использовали плезиозавры. Единственная современная группа с четырьмя плавниками — это морские черепахи, которые используют только переднюю пару для движения. Конибер и Бакленд уже сравнивали ласты с крыльями птиц. Однако такое сравнение было не очень информативным, поскольку механика полета птиц в этот период была плохо изучена. К середине девятнадцатого века обычно предполагалось, что плезиозавры использовали гребковое движение. Ласты были бы перемещены вперед в горизонтальном положении, чтобы минимизировать трение, а затем повернуты в осевом направлении в вертикальное положение, чтобы быть оттянутыми назад, вызывая максимально возможную реактивную силу . На самом деле, такой метод был бы очень неэффективным: обратный гребок в этом случае не создает тяги, а задний гребок создает огромную турбулентность. В начале двадцатого века недавно открытые принципы полета птиц навели нескольких исследователей на мысль, что плезиозавры, как черепахи и пингвины, совершали летательные движения во время плавания. Это было, например, предложено Эберхардом Фраасом в 1905 году [95] и Отенио Абелем в 1908 году [96] . Во время полета движение плавника более вертикальное, его кончик описывает овал или «8». В идеале плавник сначала движется наклонно вперед и вниз, а затем, после небольшого втягивания и вращения, пересекает этот путь снизу, чтобы быть вытянутым вперед и вверх. Во время обоих гребков, вниз и вверх, согласно принципу Бернулли , прямая и восходящая тяга создается выпукло изогнутым верхним профилем плавника, передний край слегка наклонен относительно потока воды, в то время как турбулентность минимальна. Однако, несмотря на очевидные преимущества такого способа плавания, в 1924 году первое систематическое исследование мускулатуры плезиозавров, проведенное Дэвидом Мередитом Сирсом Уотсоном, пришло к выводу, что они все же выполняли гребковые движения. [97]
В середине двадцатого века «модель гребли» Уотсона оставалась доминирующей гипотезой относительно плавательного гребка плезиозавров. В 1957 году Ламберт Беверли Холстед , в то время носивший фамилию Тарло, предложил вариант: задние конечности гребли бы в горизонтальной плоскости, а передние конечности гребли бы, перемещаясь вниз и назад. [98] [99] В 1975 году традиционная модель была оспорена Джейн Энн Робинсон , которая возродила «летящую» гипотезу. Она утверждала, что основные группы мышц были оптимально расположены для вертикального движения ласт, а не для горизонтального подтягивания конечностей, и что форма плечевых и тазобедренных суставов исключала бы вертикальное вращение, необходимое для гребли. [100] В последующей статье Робинсон предположил, что кинетическая энергия , генерируемая силами, прикладываемыми к туловищу при гребках, могла сохраняться и высвобождаться в виде упругой энергии в грудной клетке, что позволяло создать особенно эффективную и динамичную двигательную систему. [101]
В модели Робинсон и нисходящий, и восходящий гребки были бы мощными. В 1982 году ее критиковали Сэмюэль Тарситано, Эберхард Фрей и Юрген Рисс, которые утверждали, что, хотя мышцы на нижней стороне плечевых и тазовых пластин были явно достаточно мощными, чтобы тянуть конечности вниз, сопоставимые группы мышц на верхней части этих пластин для подъема конечностей просто отсутствовали, и, если бы они присутствовали, не могли бы быть задействованы с силой, их выпячивание несло опасность повреждения внутренних органов. Они предложили более ограниченную модель полета, в которой мощный нисходящий гребок сочетался с в значительной степени немощным восстановлением, при этом плавник возвращался в исходное положение под действием импульса движущегося вперед и временно тонущего тела. [102] [103] Эта модифицированная модель полета стала популярной интерпретацией. Меньше внимания было уделено альтернативной гипотезе Стивена Годфри, выдвинутой в 1984 году. Он предположил, что и передние, и задние конечности совершают глубокие гребковые движения назад в сочетании с мощным гребком вперед, что напоминает движение передних конечностей морских львов. [104]
В 2010 году Фрэнк Сандерс и Кеннет Карпентер опубликовали исследование, в котором пришли к выводу, что модель Робинсона была верной. Фрей и Рисс ошиблись бы, утверждая, что плечевые и тазовые пластины не имеют мышц, прикрепленных к их верхним сторонам. Хотя эти группы мышц, вероятно, не были очень мощными, это легко могло быть компенсировано большими мышцами на спине, особенно широчайшей мышцей спины , которая была бы хорошо развита ввиду высоких шипов на позвоночнике. Кроме того, плоское строение плечевых и тазобедренных суставов четко указывало на то, что основное движение было вертикальным, а не горизонтальным. [105]

Как и все четвероногие с конечностями, плезиозавры должны были иметь определенную походку , скоординированную схему движения, в данном случае ласт. Из всех возможностей, на практике внимание в основном было направлено на вопрос о том, двигались ли передняя и задняя пары одновременно, так что все четыре ласта были задействованы в один и тот же момент, или поочередно, когда каждая пара использовалась по очереди. Фрей и Рисс в 1991 году предложили альтернативную модель, которая имела бы преимущество более непрерывного движения. [106] В 2000 году Теагартен Лингхэм-Солиар уклонился от ответа, заключив, что, как и морские черепахи, плезиозавры использовали только переднюю пару для мощного гребка. Задняя пара использовалась бы только для рулевого управления. Лингхэм-Солиар вывел это из формы тазобедренного сустава, который допускал бы только ограниченное вертикальное движение. Более того, разделение функций движения и рулевого управления облегчило бы общую координацию тела и предотвратило бы слишком экстремальный наклон . Он отверг гипотезу Робинсона о том, что упругая энергия хранится в грудной клетке, посчитав ребра слишком жесткими для этого. [107]
Интерпретация Фрея и Рисса стала доминирующей, но была оспорена в 2004 году Сандерсом, который экспериментально показал, что, в то время как альтернативное движение могло вызвать чрезмерную тангажировку, одновременное движение вызвало бы лишь небольшую тангажировку, которую можно было бы легко контролировать задними ластами. Из других осевых движений, крен мог бы контролироваться попеременным включением ласт правой или левой стороны, а рыскание — длинной шеей или вертикальным хвостовым плавником. Сандерс не верил, что задняя пара не использовалась для движения, заключив, что ограничения, налагаемые тазобедренным суставом, были весьма относительными. [108] В 2010 году Сандерс и Карпентер пришли к выводу, что при чередующемся аллюре турбулентность, вызванная передней парой, препятствовала бы эффективному действию задней пары. Кроме того, длительная фаза скольжения после одновременного включения была бы очень энергоэффективной. [105] Также возможно, что походка была необязательной и была адаптирована к обстоятельствам. Во время быстрого равномерного преследования альтернативное движение было бы полезным; в засаде одновременный удар сделал бы пиковую скорость возможной. При поиске добычи на большем расстоянии сочетание одновременного движения с скольжением стоило бы меньше всего энергии. [109] В 2017 году исследование Люка Маскатта с использованием модели робота пришло к выводу, что задние ласты были активно задействованы, что позволило увеличить движущую силу на 60% и эффективность на 40%. Не было бы единой оптимальной фазы для всех условий, походка, вероятно, менялась в зависимости от ситуации. [110]

В целом, трудно определить максимальную скорость вымерших морских существ. Для плезиозавров это осложняется отсутствием консенсуса относительно их гребка ластами и походки. Точных расчетов их числа Рейнольдса нет . Ископаемые отпечатки показывают, что кожа была относительно гладкой, не чешуйчатой, и это могло уменьшить сопротивление формы . [105] На коже присутствуют небольшие морщины, которые могли предотвратить разделение ламинарного потока в пограничном слое и тем самым уменьшить трение кожи .
Устойчивую скорость можно оценить, вычислив сопротивление упрощенной модели тела, к которому может приблизиться вытянутый сфероид , и устойчивый уровень выработки энергии мышцами . Первое исследование этой проблемы было опубликовано Джуди Массаре в 1988 году. [111] Даже при предположении низкой гидродинамической эффективности 0,65 модель Массаре, казалось, указывала на то, что плезиозавры, если бы они были теплокровными, могли бы двигаться со скоростью четыре метра в секунду, или около четырнадцати километров в час, что значительно превышает известные скорости современных дельфинов и китов. [112] Однако в 2002 году Рёсукэ Мотани показал, что формулы, которые использовал Массаре, были несовершенны. Пересчет с использованием исправленных формул дал скорость в полметра в секунду (1,8 км/ч) для холоднокровного плезиозавра и полтора метра в секунду (5,4 км/ч) для эндотермического плезиозавра. Даже самая высокая оценка примерно на треть ниже скорости современных китообразных . [113]
Массаре также попыталась сравнить скорости плезиозавров со скоростями двух других основных групп морских рептилий, ихтиозавров и мозазаврид . Она пришла к выводу, что плезиозавры были примерно на двадцать процентов медленнее, чем продвинутые ихтиозавры, которые использовали очень эффективное туннеобразное движение, колеблясь только хвостом, но на пять процентов быстрее, чем мозазаврид, которые, как предполагалось, плавали с помощью неэффективного ангуллиформного, похожего на угорь, движения тела. [112]
Многие виды плезиозавров могли значительно различаться по скорости плавания, отражая различные формы тела, присутствующие в группе. В то время как короткошеие «плиозавроморфы» (например, Liopleurodon ) могли быть быстрыми пловцами, длинношеие «плезиозавроморфы» были созданы больше для маневренности, чем для скорости, замедляемые сильным трением кожи, но способные к быстрому движению качения. Некоторые длинношеие формы, такие как Elasmosauridae , также имеют относительно короткие короткие ласты с низким соотношением сторон , что еще больше снижает скорость, но улучшает качение. [114]
Мало данных, которые точно показывают, насколько глубоко ныряли плезиозавры. То, что они ныряли на значительную глубину, доказано следами декомпрессионной болезни . Головки плечевых и бедренных костей со многими окаменелостями показывают некроз костной ткани, вызванный слишком быстрым подъемом после глубокого погружения. Однако это не позволяет вывести некоторую точную глубину, поскольку повреждение могло быть вызвано несколькими очень глубокими погружениями или, в качестве альтернативы, большим количеством относительно неглубоких спусков. Позвонки не показывают таких повреждений: они, вероятно, были защищены превосходным кровоснабжением, которое стало возможным благодаря артериям, входящим в кость через два foramina subcentralia , больших отверстия в их нижней части. [115]
Спуску помогала бы отрицательная сила Архимеда , т. е. плотность была бы выше, чем у воды. Конечно, это имело бы недостаток, затрудняя подъем. Молодые плезиозавры демонстрируют пахиостоз , экстремальную плотность костной ткани, что могло бы увеличить относительный вес. У взрослых особей более губчатая кость. Гастролиты были предложены как метод увеличения веса [116] или даже как средство достижения нейтральной плавучести , проглатывая или выплевывая их снова по мере необходимости. [117] Они также могли использоваться для повышения устойчивости. [118]
Относительно большие глаза Cryptocleididae рассматриваются как приспособление к глубоководному нырянию. [119]
Исследование 2020 года показало, что зауроптеригии полагались на вертикальные удары хвоста, как и китообразные . У плезиозавров туловище было жестким, поэтому это действие было более ограниченным и происходило в сочетании с ластами. [76]
Традиционно считалось, что вымершие группы рептилий были холоднокровными, как и современные рептилии. Новые исследования последних десятилетий привели к выводу, что некоторые группы, такие как тероподы и птерозавры , были, скорее всего, теплокровными. Трудно определить, были ли плезиозавры также теплокровными. Одним из признаков высокого метаболизма является наличие быстрорастущей фиброламеллярной кости. Однако пахиостоз у молодых особей затрудняет установление того, обладали ли плезиозавры такой костью. Однако удалось проверить его наличие у более базальных членов более инклюзивной группы, к которой принадлежали плезиозавры, Sauropterygia . Исследование, проведенное в 2010 году, пришло к выводу, что фиброламеллярная кость изначально присутствовала у завроптеригий. [120] В последующей публикации 2013 года было обнаружено, что у Nothosauridae отсутствовал этот тип костной матрицы, но у базальных Pistosauria он был, что является признаком более высокого метаболизма. [121] Таким образом, более экономно предполагать, что более продвинутые пистозавры, плезиозавры, также имели более быстрый метаболизм. В статье, опубликованной в 2018 году, утверждалось, что у плезиозавров были показатели метаболизма в состоянии покоя (RMR) в диапазоне птиц на основе количественного остеогистологического моделирования. [122] Однако эти результаты проблематичны с точки зрения общих принципов физиологии позвоночных (см. закон Клейбера ); данные изотопных исследований зубной эмали плезиозавров действительно предполагают эндотермию при более низких RMR, с предполагаемой температурой тела около 26 °C (79 °F). [123]

Поскольку рептилии в целом являются яйцекладущими , до конца двадцатого века считалось возможным, что более мелкие плезиозавры могли выползать на пляж, чтобы откладывать яйца, как современные черепахи . Их сильные конечности и плоская нижняя часть, казалось, делали это осуществимым. Этот метод, например, защищал Холстед. Однако, поскольку эти конечности больше не имели функциональных локтевых или коленных суставов, а нижняя часть из-за своей плоской формы создавала бы большое трение, уже в девятнадцатом веке была выдвинута гипотеза, что плезиозавры были живородящими. Кроме того , было трудно представить, как самые крупные виды, такие большие, как киты, могли выжить, выброшенные на берег. Ископаемые находки эмбрионов ихтиозавров показали, что по крайней мере одна группа морских рептилий рожала живых детенышей. Первым, кто утверждал, что подобные эмбрионы были найдены у плезиозавров, был Гарри Говер Сили , который в 1887 году сообщил о приобретении узелка с четырьмя-восемью крошечными скелетами. [124] В 1896 году он описал это открытие более подробно. [125] Если бы они были подлинными, эмбрионы плезиозавров были бы очень маленькими, как у ихтиозавров. Однако в 1982 году Ричард Энтони Тулборн показал, что Сили был обманут «подделанной» окаменелостью гнезда раков. [126]
Фактический образец плезиозавра, найденный в 1987 году, в конечном итоге доказал, что плезиозавры рождали живых детенышей: [127] Эта окаменелость беременной Polycotylus latippinus показывает, что эти животные рожали одного крупного детеныша и, вероятно, вкладывали родительскую заботу в свое потомство, подобно современным китам. Детеныш был 1,5 метра (пять футов) в длину и, таким образом, был больше по сравнению со своей матерью длиной пять метров (шестнадцать футов), что указывает на K-стратегию в размножении. [128] Мало что известно о темпах роста или возможном половом диморфизме .
Из родительской заботы, на которую указывает большой размер детенышей, можно сделать вывод, что социальное поведение в целом было относительно сложным. [127] Неизвестно, охотились ли плезиозавры стаями. Относительный размер их мозга, по-видимому, типичен для рептилий. Из чувств важными были зрение и обоняние, слух — в меньшей степени; эласмозавриды полностью утратили стремя . Было высказано предположение, что у некоторых групп череп располагал электрочувствительными органами. [129] [130]
Некоторые окаменелости плезиозавров показывают патологии , вызванные болезнью или старостью. В 2012 году была описана нижняя челюсть плиозавра с суставом челюсти, явно пораженным артритом , типичным признаком старения . [131]
Окаменелости плезиозавров были найдены на всех континентах, включая Антарктиду . [132]
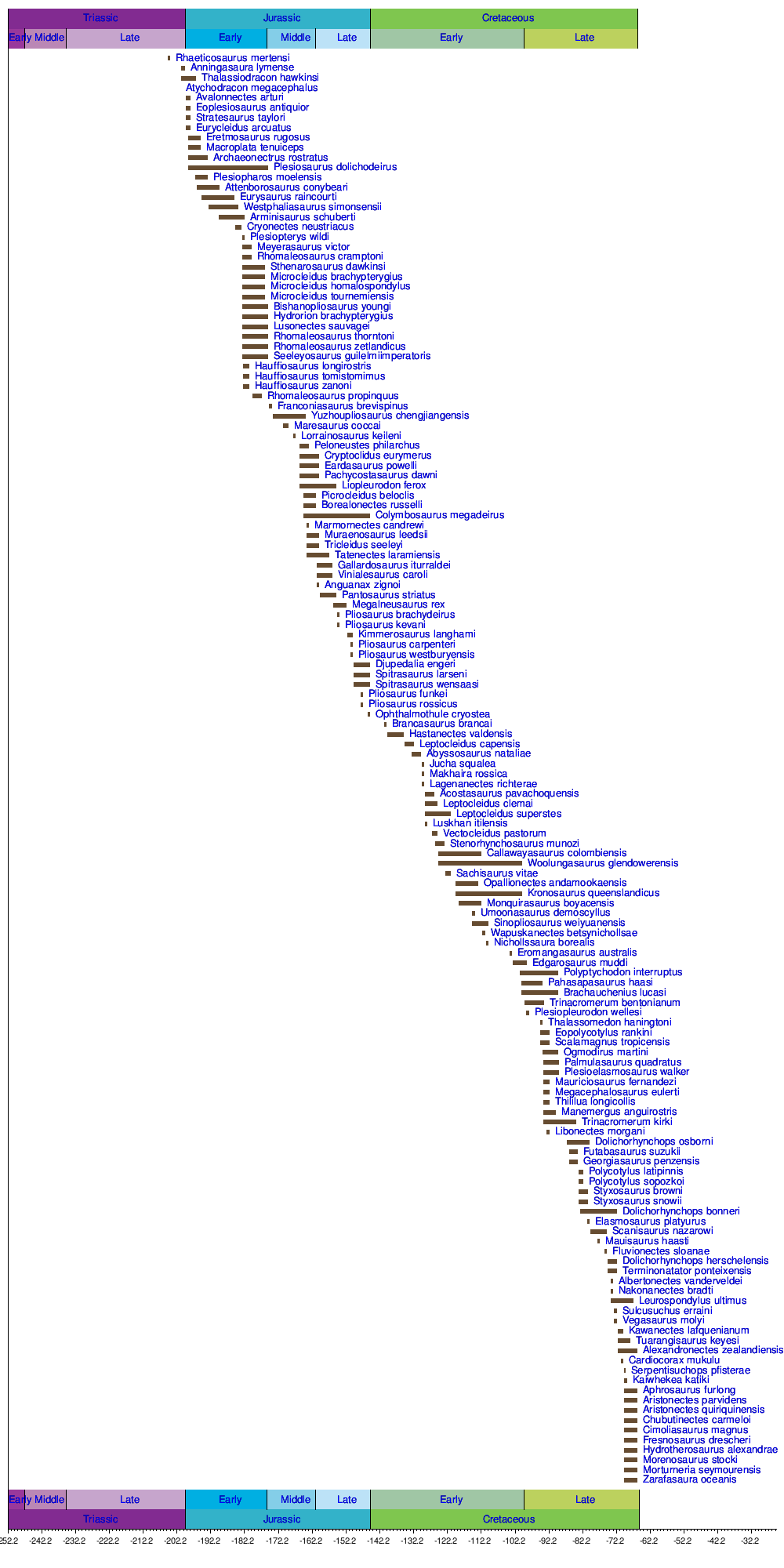
Ниже приведен список геологических формаций , в которых были обнаружены окаменелости плезиозавров.
_trailer_-_Plesiosaurus_1.png/1280px-When_Dinosaurs_Ruled_the_Earth_(1970)_trailer_-_Plesiosaurus_1.png)
Убеждение, что плезиозавры являются динозаврами, является распространённым заблуждением , и в популярной культуре плезиозавров часто ошибочно изображают как динозавров . [139] [140]
Было высказано предположение, что легенды о морских змеях и современные наблюдения предполагаемых монстров в озерах или море могут быть объяснены выживанием плезиозавров в наше время. Это криптозоологическое предложение было отвергнуто научным сообществом в целом, которое считает его основанным на фантазии и лженауке . Было показано, что предполагаемые туши плезиозавров являются частично разложившимися трупами гигантских акул . [141] [142] [143]
Хотя часто сообщается, что Лох-Несское чудовище похоже на плезиозавра, его также часто описывают как выглядящее совершенно иначе. Было представлено несколько причин, по которым оно вряд ли является плезиозавром. Они включают в себя предположение, что вода в озере слишком холодная для предполагаемой холоднокровной рептилии, чтобы она могла легко выжить, тот факт, что остеология шеи плезиозавра позволяет абсолютно с уверенностью сказать, что плезиозавр не мог поднять голову из воды, как лебедь, как это делает Лох-Несское чудовище, предположение, что дышащих воздухом животных было бы легко увидеть, когда бы они ни появлялись на поверхности, чтобы подышать, [144] тот факт, что озеро слишком мало и содержит недостаточно пищи, чтобы содержать размножающуюся колонию крупных животных, и, наконец, тот факт, что озеро образовалось всего 10 000 лет назад в конце последнего ледникового периода , а последнее ископаемое появление плезиозавров датируется более 66 миллионами лет назад. [145] Частые объяснения наблюдений включают волны , плавающие неодушевленные предметы, игру света, плавание известных животных и розыгрыши. [146] Тем не менее, в народном воображении плезиозавры стали отождествляться с чудовищем Лох-Несса. Это сделало плезиозавров более известными широкой публике. [147]