Превращение молекулярного азота в биологически доступные соединения азота.
Азотфиксация — это химический процесс, при котором молекулярный азот ( N 2), который имеет прочную тройную ковалентную связь , превращается в аммиак ( NH 3) или родственные азотистые соединения, обычно в почве или водных системах [1] , а также в промышленности . Азот в воздухе представляет собой молекулярный диазот , относительно нереактивную молекулу, которая метаболически бесполезна для всех микроорганизмов, за исключением нескольких. Биологическая фиксация азота, или диазотрофия, представляет собой важный микробный процесс, который превращает газообразный диазот (N 2 ) в аммиак (NH 3 ) с помощью белкового комплекса нитрогеназы (Nif). [2] [3]
Фиксация азота необходима для жизни, поскольку фиксированные неорганические соединения азота необходимы для биосинтеза всех азотсодержащих органических соединений , таких как аминокислоты и белки , нуклеозидтрифосфаты и нуклеиновые кислоты . Как часть азотного цикла , он необходим для сельского хозяйства и производства удобрений . Это также косвенно имеет отношение к производству всех азотистых химических соединений, к которым относятся некоторые взрывчатые вещества, фармацевтические препараты и красители.
Фиксация азота осуществляется естественным путем в почве микроорганизмами, называемыми диазотрофами , которые включают бактерии , такие как Azotobacter , и археи . Некоторые азотфиксирующие бактерии имеют симбиотические отношения с группами растений, особенно с бобовыми . [4] Более слабые несимбиотические отношения между диазотрофами и растениями часто называют ассоциативными, как видно из фиксации азота на корнях риса . Азотфиксация происходит между некоторыми термитами и грибами . [5] Он естественным образом возникает в воздухе в результате образования NO x при ударе молнии . [6] [7]
Все биологические реакции, связанные с процессом фиксации азота, катализируются ферментами, называемыми нитрогеназами . [8] Эти ферменты содержат железо , часто со вторым металлом, обычно молибденом , но иногда и ванадием .
История
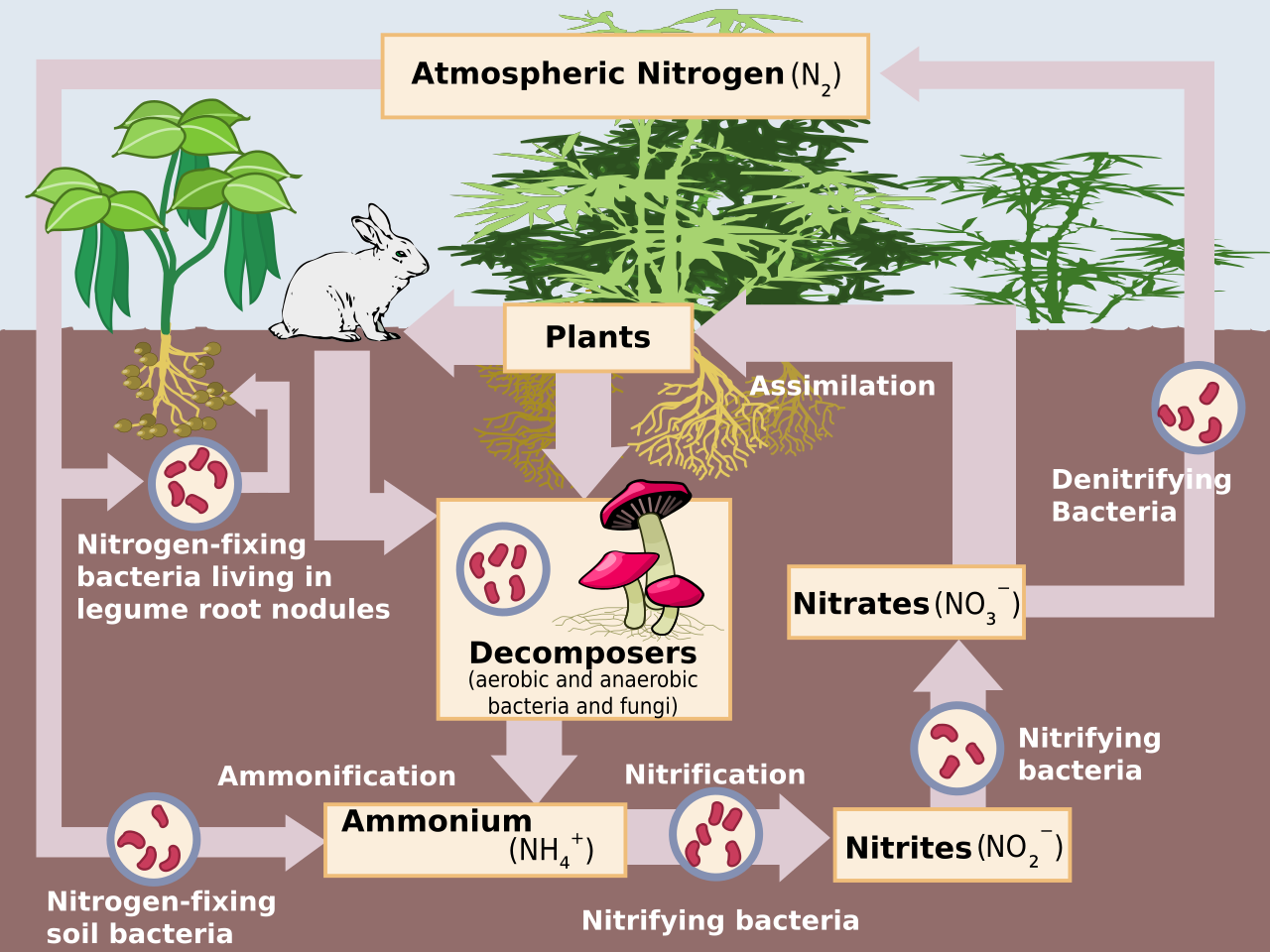
Схематическое изображение круговорота азота . Абиотическая азотфиксация исключена.
Биологическая фиксация азота была открыта Жаном-Батистом Буссенго в 1838 году. [9] [10] Позже, в 1880 году, процесс, посредством которого она происходит, был открыт немецкими агрономами Германом Хеллригелем и Германом Вильфартом [де] [11] и был полностью описан. голландский микробиолог Мартинус Бейеринк . [12]
«Длительные исследования связи растений с усвоением азота, начатые Соссюром , Виллем , Лоузом , Гилбертом и другими, завершились открытием Хеллригелем и Уилфартом в 1887 году симбиотической фиксации». [13]
«Эксперименты Боссенго в 1855 году и Пью, Гилберта и Лоуза в 1887 году показали, что азот не попадает в растение напрямую. Открытие Германом Хеллригелем и Германом Вильфартом в 1886-88 годах роли азотфиксирующих бактерий открыло новую эру Почвоведение». [14]
Биологическая фиксация азота (БНФ) происходит, когда атмосферный азот превращается в аммиак ферментом нитрогеназой . [1] Общая реакция на BNF такова:
Процесс сопряжен с гидролизом 16 эквивалентов АТФ и сопровождается совместным образованием одного эквивалента H. 2. [15] Преобразование N 2Преобразование аммиака происходит в металлический кластер под названием FeMoco , аббревиатура железомолибденового кофактора . Механизм протекает через серию стадий протонирования и восстановления , на которых активный центр FeMoco гидрирует N. 2субстрат. [16] У свободноживущих диазотрофов аммиак, вырабатываемый нитрогеназой, ассимилируется в глутамат через путь глутаминсинтетаза /глутаматсинтаза. Микробные гены nif, необходимые для фиксации азота, широко распространены в различных средах. [17]
Например, было показано, что разлагающаяся древесина, которая обычно имеет низкое содержание азота, является местом обитания диазотрофного сообщества. [18] [19] Бактерии обогащают древесный субстрат азотом посредством фиксации, тем самым обеспечивая разложение валежной древесины грибами. [20]
Нитрогеназы быстро разлагаются кислородом. По этой причине многие бактерии прекращают выработку фермента в присутствии кислорода. Многие азотфиксирующие организмы существуют только в анаэробных условиях, дыша, чтобы снизить уровень кислорода, или связывая кислород с помощью белка , такого как леггемоглобин . [1]
Важность азота
Атмосферный азот недоступен для большинства организмов [21] , поскольку его тройная ковалентная связь очень прочна. Большинство из них поглощают фиксированный азот из различных источников. На каждые 100 атомов углерода ассимилируется примерно от 2 до 20 атомов азота. Атомное соотношение углерода (C) : азот (N) : фосфор (P), наблюдаемое в среднем в планктонной биомассе, было первоначально описано Альфредом Редфилдом [22] , который определил стехиометрическое соотношение между атомами C:N:P, соотношение Редфилда. , чтобы быть 106:16:1. [22]
Нитрогеназа
Белковый комплекс нитрогеназа катализирует восстановление газообразного азота (N 2 ) до аммиака (NH 3 ). [23] У цианобактерий эта ферментная система расположена в специализированной клетке, называемой гетероцистой . [24] Производство нитрогеназного комплекса генетически регулируется, а активность белкового комплекса зависит от концентрации кислорода в окружающей среде, а также внутри- и внеклеточных концентраций аммиака и окисленных форм азота (нитратов и нитритов). [25] [26] [27] Кроме того, считается, что совокупные концентрации аммония и нитрата ингибируют N Fix , особенно когда внутриклеточные концентрации 2-оксоглутарата (2-OG) превышают критический порог. [28] Специализированная клетка-гетероциста необходима для работы нитрогеназы из-за ее чувствительности к кислороду окружающей среды. [29]
Нитрогеназа состоит из двух белков: каталитического железозависимого белка, обычно называемого белком MoFe, и восстанавливающего белка, содержащего только железо (белок Fe). Существует три различных железозависимых белка: молибден-зависимый, ванадий-зависимый и только железо, причем все три варианта белка нитрогеназы содержат компонент железосодержащего белка. Молибдензависимая нитрогеназа является наиболее часто встречающейся нитрогеназой. [23] Различные типы нитрогеназы можно определить по конкретному железосодержащему белковому компоненту. [30] Нитрогеназа высоко консервативна. Экспрессия генов посредством секвенирования ДНК позволяет определить, какой белковый комплекс присутствует в микроорганизме и потенциально экспрессируется. Чаще всего ген nif H используется для идентификации присутствия молибдензависимой нитрогеназы, за которым следуют близкородственные нитрогеназоредуктазы (компонент II) vnf H и anf H, представляющие ванадий-зависимую и железо-нитрогеназу соответственно. [31] При изучении экологии и эволюции азотфиксирующих бактерий ген nifH является наиболее широко используемым биомаркером. [32] nif H имеет два сходных гена anf H и vnfH, которые также кодируют нитрогеназоредуктазный компонент нитрогеназного комплекса. [33]
Цианобактерии , широко известные как сине-зеленые водоросли, обитают почти во всех освещенных средах на Земле и играют ключевую роль в круговороте углерода и азота в биосфере . В общем, цианобактерии могут использовать различные неорганические и органические источники связанного азота, такие как нитрат , нитрит , аммоний , мочевина или некоторые аминокислоты . Некоторые штаммы цианобактерий также способны к диазотрофному росту - способность, которая, возможно, присутствовала у их последнего общего предка в архейском эоне. [38] Азотфиксация естественным образом происходит не только в почвах, но и в водных системах, включая как пресноводные, так и морские. [39] [40] Действительно, количество азота, зафиксированного в океане, по крайней мере, такое же, как и на суше. [41] Считается , что колониальная морская цианобактерия Trichodesmium фиксирует азот в таких масштабах, что на ее долю приходится почти половина фиксации азота в морских системах во всем мире. [42] Морские поверхностные лишайники и нефотосинтезирующие бактерии, принадлежащие к Proteobacteria и Planctomycetes, фиксируют значительное количество атмосферного азота. [43] Виды азотфиксирующих цианобактерий в пресных водах включают: Aphanizomenon и Dolichospermum (ранее Anabaena). [44] У таких видов есть специализированные клетки, называемые гетероцитами , в которых фиксация азота происходит с помощью фермента нитроциты. [45] [46]
Симбиозы корневых клубеньков
Семья бобовых
На корне этой фасоли видны узелки.
К растениям, которые способствуют фиксации азота, относятся растения семейства бобовых — Fabaceae — с такими таксонами , как кудзу , клевер , соя , люцерна , люпин , арахис и ройбуш . [35] Они содержат симбиотические бактерии -ризобии в клубеньках своей корневой системы , производящие соединения азота, которые помогают растению расти и конкурировать с другими растениями. [47] Когда растение умирает, фиксированный азот высвобождается, делая его доступным для других растений; это помогает удобрить почву . [1] [48] Подавляющее большинство бобовых имеют эту ассоциацию, но некоторые роды (например, Styphnolobium ) этого не делают. Во многих традиционных методах ведения сельского хозяйства на полях поочередно выращивают различные типы культур, среди которых обычно есть культура, состоящая в основном или полностью из клевера . [ нужна цитата ]
Эффективность фиксации в почве зависит от многих факторов, в том числе от состава бобовых , воздушно-почвенных условий. Например, фиксация азота красным клевером может составлять от 50 до 200 фунтов на акр (от 56 до 224 кг/га). [49]
Незерновые
Разрезанный корневой клубень ольхи
Способность фиксировать азот в клубеньках имеется у актиноризных растений, таких как ольха и ольха , с помощью бактерий Frankia . Они встречаются в 25 родах в отрядах Cucurbitales , Fagales и Rosales , которые вместе с Fabales образуют азотфиксирующую кладу евросид . Способность фиксировать азот не всегда присутствует в этих семьях. Например, из 122 родов Rosaceae только четыре фиксируют азот. Fabales были первой линией, ответвившейся от этой азотфиксирующей клады; таким образом, способность фиксировать азот может быть плезиоморфной и впоследствии утрачиваться у большинства потомков исходного азотфиксирующего растения; однако возможно, что основные генетические и физиологические потребности присутствовали в зачаточном состоянии у самых недавних общих предков всех этих растений, но развились до полной функции лишь у некоторых из них. [50]
Кроме того, трема ( Parasponia ), тропический род семейства Cannabaceae , необычайно способен взаимодействовать с ризобиями и образовывать азотфиксирующие клубеньки. [51]
Другие симбионты растений
Некоторые другие растения живут вместе с цианобионтами (цианобактериями, такими как Nostoc ), которые фиксируют для них азот:
Метод фиксации азота был впервые описан Генри Кавендишем в 1784 году с использованием электрических дуг, реагирующих с азотом и кислородом воздуха. Этот метод был реализован в процессе Биркеланда-Эйда в 1903 году. [56] Фиксация азота молнией - очень похожий естественный процесс.
Возможность реакции атмосферного азота с некоторыми химическими веществами впервые была обнаружена Дефоссом в 1828 году. Он заметил, что смеси оксидов щелочных металлов и углерода реагируют с азотом при высоких температурах. С использованием карбоната бария в качестве исходного материала в 1860-х годах стал доступен первый коммерческий процесс, разработанный Маргаритой и Сурдевалем. Образующийся цианид бария реагирует с водяным паром, образуя аммиак. В 1898 году Франк и Каро разработали так называемый процесс Франка-Каро для фиксации азота в форме цианамида кальция . Процесс затмил процесс Габера , открытый в 1909 году. [57] [58]
Габеровский процесс
Аппаратура для исследования фиксации азота альфа-лучами (Лаборатория исследования фиксированного азота, 1926 г.)
Доминирующим промышленным методом производства аммиака является процесс Габера, также известный как процесс Габера-Боша. [59] Производство удобрений в настоящее время является крупнейшим источником антропогенного фиксированного азота в наземной экосистеме . Аммиак является необходимым предшественником удобрений , взрывчатых веществ и других продуктов. Процесс Габера требует высокого давления (около 200 атм) и высоких температур (не менее 400 °C), которые являются обычными условиями для промышленного катализа. В этом процессе используется природный газ в качестве источника водорода и воздух в качестве источника азота. Производство аммиака привело к интенсификации производства азотных удобрений во всем мире [60] , и ему приписывают поддержку роста численности населения с примерно 2 миллиардов человек в начале 20-го века до примерно 8 миллиардов человек сейчас. [61]
Гомогенный катализ
Было проведено много исследований по открытию катализаторов фиксации азота, часто с целью снижения энергетических затрат. Однако такие исследования до сих пор не смогли приблизиться к эффективности и простоте процесса Габера. Многие соединения реагируют с атмосферным азотом с образованием динитрогенных комплексов . Первым сообщенным диазотистым комплексом был Ru(NH 3) 5( Н 2) 2+ . [62] Некоторые растворимые комплексы катализируют азотфиксацию. [63]
Молния
Молния нагревает воздух вокруг себя, разрывая связи N. 2начинается образование азотистой кислоты.
Азот можно фиксировать с помощью молнии, превращая газообразный азот ( N 2) и газообразный кислород ( O 2) в атмосфере на NO x ( оксиды азота ). Затем _ 2Молекула очень стабильна и инерционна из-за тройной связи между атомами азота. [64] Молния производит достаточно энергии и тепла, чтобы разорвать эту связь [64] , позволяя атомам азота вступать в реакцию с кислородом, образуя NO. Икс. Эти соединения не могут быть использованы растениями, но по мере охлаждения молекула реагирует с кислородом с образованием NO. 2, [65] который, в свою очередь, реагирует с водой с образованием HNO 2( азотистая кислота ) или HNO 3( азотная кислота ). Когда эти кислоты просачиваются в почву, они образуют NO. 3(нитрат) , который полезен растениям. [66] [64]
^ Захран Х.Х. (декабрь 1999 г.). «Ризобиально-бобовый симбиоз и азотфиксация в суровых условиях и в засушливом климате». Обзоры микробиологии и молекулярной биологии . 63 (4): 968–89, оглавление. дои :10.1128/MMBR.63.4.968-989.1999. ПМК 98982 . ПМИД 10585971.
^ Сапунцис П., де Вержес Дж., Руск К., Сильерс М., Форстер Б.Дж., Поульсен М. (2016). «Возможность фиксации азота в симбиозе термитов, выращивающих грибы». Границы микробиологии . 7 : 1993. doi : 10.3389/fmicb.2016.01993 . ПМК 5156715 . ПМИД 28018322.
^ Слоссон Э (1919). Творческая химия. Нью-Йорк, штат Нью-Йорк: The Century Co., стр. 19–37.
^ Хилл Р.Д., Ринкер Р.Г., Уилсон HD (1979). «Фиксация атмосферного азота молнией». Дж. Атмос. Наука . 37 (1): 179–192. Бибкод : 1980JAtS...37..179H. doi : 10.1175/1520-0469(1980)037<0179:ANFBL>2.0.CO;2 .
^ Вагнер СК (2011). «Биологическая азотфиксация». Знания о природном образовании . 3 (10): 15. Архивировано из оригинала 13 сентября 2018 года . Проверено 29 января 2019 г.
^ Буссенго (1838). «Recherches chimiques sur la растительности, предприятия dans le, но d'examiner si les plantes prennent de l'azote à l'atmography» [Химические исследования растительности, предпринятые с целью изучения того, поглощают ли растения азот из атмосферы]. Annales de Chimie et de Physique . 2-я серия (на французском языке). 67 : 5–54.и 69: 353–367.
^ Смил В. (2001). Обогащение Земли . Массачусетский Институт Технологий.
^ Хеллригель Х, Уилфарт Х (1888). Untersuchungen über die Stickstoffnahrung der Gramineen und Leguminosen [ Исследования потребления азота злаками и бобовыми ] (на немецком языке). Берлин, Германия: Buchdruckerei der "Post" Kayssler & Co.
^ Бейеринк М.В. (1901). «Убер олигонитрофильный микробен». Centralblatt für Bakteriologie, Parasitenkunde, Infektionskrankheiten und Hygiene (на немецком языке). 7 (16): 561–582.
^ Говард С. Рид (1942) Краткая история науки о растениях , страница 230, Chronic Publishing
^ Ринне К.Т., Раджала Т., Пелтониеми К., Чен Дж., Смоландер А., Мякипаа Р. (2017). «Скорость накопления и источники внешнего азота в гниющей древесине в лесу, где преобладает ель обыкновенная». Функциональная экология . 31 (2): 530–541. Бибкод : 2017FuEco..31..530R. дои : 10.1111/1365-2435.12734 . ISSN 1365-2435. S2CID 88551895.
^ Хоппе Б., Каль Т., Караш П., Вубет Т., Баухус Дж., Бускот Ф., Крюгер Д. (2014). «Сетевой анализ выявляет экологические связи между азотфиксирующими бактериями и дереворазрушающими грибами». ПЛОС ОДИН . 9 (2): e88141. Бибкод : 2014PLoSO...988141H. дои : 10.1371/journal.pone.0088141 . ПМЦ 3914916 . ПМИД 24505405.
^ Тласкал В., Брабцова В., Ветровски Т., Джомура М., Лопес-Мондехар Р., Оливейра Монтейро Л.М. и др. (январь 2021 г.). «Взаимодополняющая роль обитающих в древесине грибов и бактерий способствует разложению валежной древесины». mSystems . 6 (1). doi : 10.1128/mSystems.01078-20. ПМЦ 7901482 . ПМИД 33436515.
^ Делвич CC (1983). «Круговорот элементов в биосфере». В Ляухли А., Биелески Р.Л. (ред.). Неорганическое питание растений . Энциклопедия физиологии растений. Берлин, Гейдельберг: Springer. стр. 212–238. дои : 10.1007/978-3-642-68885-0_8. ISBN978-3-642-68885-0.
^ ab Редфилд AC (1958). «Биологический контроль химических факторов окружающей среды». Американский учёный . 46 (3): 230А–221. ISSN 0003-0996. JSTOR 27827150.
^ ab Burgess BK, Lowe DJ (ноябрь 1996 г.). «Механизм нитрогеназы молибдена». Химические обзоры . 96 (7): 2983–3012. дои : 10.1021/cr950055x. ПМИД 11848849.
^ Петерсон Р.Б., Волк CP (декабрь 1978 г.). «Высокое восстановление нитрогеназной активности и Fe-меченной нитроцисты в гетероцистах, выделенных из Anabaena variabilis». Труды Национальной академии наук Соединенных Штатов Америки . 75 (12): 6271–6275. Бибкод : 1978PNAS...75.6271P. дои : 10.1073/pnas.75.12.6271 . ПМЦ 393163 . ПМИД 16592599.
^ Беверсдорф Л.Дж., Миллер Т.Р., МакМахон К.Д. (6 февраля 2013 г.). «Роль фиксации азота в токсичности цветения цианобактерий в эвтрофном озере с умеренным климатом». ПЛОС ОДИН . 8 (2): e56103. Бибкод : 2013PLoSO...856103B. дои : 10.1371/journal.pone.0056103 . ПМК 3566065 . ПМИД 23405255.
^ Галлон JR (1 марта 2001 г.). «Фиксация N2 у фототрофов: адаптация к специализированному образу жизни». Растение и почва . 230 (1): 39–48. дои : 10.1023/А: 1004640219659. ISSN 1573-5036. S2CID 22893775.
^ Паерл Х (9 марта 2017 г.). «Парадокс цианобактериальной азотфиксации в природных водах». F1000Исследования . 6 : 244. дои : 10.12688/f1000research.10603.1 . ПМЦ 5345769 . ПМИД 28357051.
^ Ли Дж. Х., Лоран С., Конде В., Беду С., Чжан CC (ноябрь 2003 г.). «Повышение уровня 2-оксоглутарата способствует развитию гетероцист у цианобактерии Anabaena sp., штамма PCC 7120». Микробиология . 149 (Часть 11): 3257–3263. дои : 10.1099/mic.0.26462-0 . ПМИД 14600238.
^ Волк CP, Эрнст А, Эльхай Дж (1994). «Метаболизм и развитие гетероцист». В Брайанте Д.А. (ред.). Молекулярная биология цианобактерий . Достижения в области фотосинтеза. Дордрехт: Springer Нидерланды. стр. 769–823. дои : 10.1007/978-94-011-0227-8_27. ISBN978-94-011-0227-8.
^ Шнайдер К., Мюллер А. (2004). «Железная нитрогеназа: исключительные каталитические, структурные и спектроскопические особенности». Смит Б.Е., Ричардс Р.Л., Ньютон В.Е. (ред.). Катализаторы азотфиксации . Фиксация азота: происхождение, применение и прогресс исследований. Дордрехт: Springer Нидерланды. стр. 281–307. дои : 10.1007/978-1-4020-3611-8_11. ISBN978-1-4020-3611-8.
^ Кноче К.Л., Аояма Э., Хасан К., Минтир С.Д. (2017). «Роль нитрогеназы и ферредоксина в механизме биоэлектрокаталитической фиксации азота мутантом Cyanobacteria Anabaena variabilis SA-1, иммобилизованным на электродах из оксида индия и олова (ITO)». Electrochimica Acta (на корейском языке). 232 : 396–403. doi :10.1016/j.electacta.2017.02.148.
^ Раймонд Дж., Зиферт Дж.Л., Стейплс CR, Бланкеншип RE (март 2004 г.). «Естественная история фиксации азота». Молекулярная биология и эволюция . 21 (3): 541–554. дои : 10.1093/molbev/msh047 . ПМИД 14694078.
^ Шюддекопф К., Хеннеке С., Лизе У., Куче М., Клипп В. (май 1993 г.). «Характеристика генов anf, специфичных для альтернативной нитрогеназы, и идентификация генов nif, необходимых для обеих нитрогеназ у Rhodobacter capsulatus». Молекулярная микробиология . 8 (4): 673–684. doi :10.1111/j.1365-2958.1993.tb01611.x. PMID 8332060. S2CID 42057860.
^ Поступление азота в древний океан - SciTechDaily
^ ab Мус Ф., Крук М.Б., Гарсия К., Гарсия Костас А., Геддес Б.А., Кури Э.Д. и др. (июль 2016 г.). Келли Р.М. (ред.). «Симбиотическая фиксация азота и проблемы ее распространения на небобовые». Прикладная и экологическая микробиология . 82 (13): 3698–3710. Бибкод : 2016ApEnM..82.3698M. дои : 10.1128/AEM.01055-16. ПМЦ 4907175 . ПМИД 27084023.
^ Дхамад А.Е., диджей Лесснер (октябрь 2020 г.). Атоми Х (ред.). «Система CRISPRi-dCas9 для архей и ее использование для изучения функции генов во время фиксации азота Methanosarcina acetivorans». Прикладная и экологическая микробиология . 86 (21): e01402–20. Бибкод : 2020ApEnM..86E1402D. дои : 10.1128/AEM.01402-20. ПМЦ 7580536 . ПМИД 32826220.
^ Бэ Х.С., Моррисон Э., Шантон Дж.П., Ограм А (апрель 2018 г.). «Метаногены вносят основной вклад в фиксацию азота в почвах Эверглейдс Флориды». Прикладная и экологическая микробиология . 84 (7): e02222–17. Бибкод : 2018ApEnM..84E2222B. дои : 10.1128/AEM.02222-17. ПМЦ 5861825 . ПМИД 29374038.
^ Латышева Н., Юнкер В.Л., Палмер В.Дж., Кодд Г.А., Баркер Д. (март 2012 г.). «Эволюция фиксации азота у цианобактерий». Биоинформатика . 28 (5): 603–606. doi : 10.1093/биоинформатика/bts008 . ПМИД 22238262.
^ Пьерелла Карлусич Дж. Дж., Пеллетье Э., Ломбард Ф., Карсик М., Дворжак Э., Колин С. и др. (июль 2021 г.). «Глобальные закономерности распределения морских азотфиксаторов по данным визуализации и молекулярным методам». Природные коммуникации . 12 (1): 4160. Бибкод : 2021NatCo..12.4160P. дои : 10.1038/s41467-021-24299-y. ПМК 8260585 . ПМИД 34230473.
↑ Эш С (13 августа 2021 г.). Эш С., Смит Дж. (ред.). «Немного света на диазотрофов». Наука . 373 (6556): 755,7–756. Бибкод : 2021Sci...373..755A. doi : 10.1126/science.373.6556.755-g. ISSN 0036-8075. S2CID 238709371.
^ Бергман Б., Санд Г., Лин С., Ларссон Дж., Карпентер Э.Дж. (май 2013 г.). «Триходезмий - широко распространенная морская цианобактерия с необычными свойствами фиксации азота». Обзоры микробиологии FEMS . 37 (3): 286–302. дои : 10.1111/j.1574-6976.2012.00352.x. ПМЦ 3655545 . ПМИД 22928644.
^ «Крупномасштабное исследование указывает на появление новых, обильных азотфиксирующих микробов на поверхности океана». ScienceDaily . Архивировано из оригинала 8 июня 2019 года . Проверено 8 июня 2019 г.
↑ Рольф С., Альмесё Л., Элмгрен Р. (5 марта 2007 г.). «Азотфиксация и численность диазотрофных цианобактерий Aphanizomenon sp. в Собственной Балтике». Серия «Прогресс в области морской экологии ». 332 : 107–118. Бибкод : 2007MEPS..332..107R. дои : 10.3354/meps332107 .
^ Кармайкл WW (12 октября 2001 г.). «Влияние на здоровье цианобактерий, продуцирующих токсины: «ЦианоHAB»". Оценка человеческого и экологического риска . 7 (5): 1393–1407. doi : 10.1080/20018091095087. ISSN 1080-7039. S2CID 83939897.
^ Боте Х., Шмитц О., Йейтс М.Г., Ньютон МЫ (декабрь 2010 г.). «Азотфиксация и водородный обмен у цианобактерий». Обзоры микробиологии и молекулярной биологии . 74 (4): 529–551. дои : 10.1128/MMBR.00033-10. ПМК 3008169 . ПМИД 21119016.
^ Op den Camp R, Streng A, De Mita S, Cao Q, Polone E, Liu W и др. (февраль 2011 г.). «Микоризный рецептор типа LysM, задействованный для ризобиального симбиоза у небобовой Parasponia». Наука . 331 (6019): 909–12. Бибкод : 2011Sci...331..909O. дои : 10.1126/science.1198181. PMID 21205637. S2CID 20501765.
^ «Биология саговников, Статья 1: Корралоидные корни саговников». www1.biologie.uni-hamburg.de . Проверено 14 октября 2021 г.
^ Рай АН (2000). «Симбиозы цианобактерий и растений». Новый фитолог . 147 (3): 449–481. дои : 10.1046/j.1469-8137.2000.00720.x . ПМИД 33862930.
^ Ван Дейнзе А., Самора П., Дело П.М., Хейтманн С., Джаяраман Д., Раджасекар С. и др. (август 2018 г.). «Азотфиксация в местных сортах кукурузы поддерживается диазотрофной микробиотой, связанной со слизью». ПЛОС Биология . 16 (8): e2006352. дои : 10.1371/journal.pbio.2006352 . ПМК 6080747 . ПМИД 30086128.
↑ Псковский М (16 июля 2019 г.). «Местная кукуруза: кому принадлежат права на «чудо-растение» Мексики?». Йель E360 .
^ Эйд С (1909). «Производство нитратов из атмосферы с помощью электрической дуги - процесс Биркеланда-Эйда». Журнал Королевского общества искусств . 57 (2949): 568–576. JSTOR 41338647.
^ Генрих Х, Невбнер Р (1934). «Die Umwandlungsgleichung Ba(CN)2 → BaCN2 + C im Temperaturgebiet von 500 bis 1000 °C» [Реакция превращения Ba(CN) 2 → BaCN 2 + C в интервале температур от 500 до 1000 °C]. З. Электрохим. Энджью. Физ. Хим . 40 (10): 693–698. дои : 10.1002/bbpc.19340401005. S2CID 179115181. Архивировано из оригинала 20 августа 2016 года . Проверено 8 августа 2016 г.
^ Кертис Х.А. (1932). Фиксированный азот.
^ Смил, В. 2004. Обогащение Земли: Фриц Хабер, Карл Бош и трансформация мирового производства продуктов питания, MIT Press.
^ Глиберт П.М. , Марангер Р. , Собота DJ, Бауман Л. (1 октября 2014 г.). «Связь Хабер-Бош – вредное цветение водорослей (HB – HAB)». Письма об экологических исследованиях . 9 (10): 105001. Бибкод : 2014ERL.....9j5001G. дои : 10.1088/1748-9326/9/10/105001 . ISSN 1748-9326. S2CID 154724892.
«Коллекция Трэвиса П. Хигнетта фотографий лаборатории исследования фиксированного азота // Цифровые коллекции Института истории науки». digital.sciencehistory.org . Проверено 16 августа 2019 г. Цифровые коллекции Института истории науки (фотографии, изображающие многочисленные этапы процесса фиксации азота, а также различное оборудование и аппараты, используемые для производства атмосферного азота, включая генераторы, компрессоры, фильтры, термостаты, вакуумные и доменные печи).
«Предлагаемый процесс фиксации атмосферного азота», историческая перспектива, Scientific American , 13 июля 1878 г., стр. 21
Снимок азотфиксаторов глобального океана путем сопоставления последовательностей с клетками в океане Тара.



.jpg/1280px-Lightning_Pritzerbe_01_(MK).jpg)

