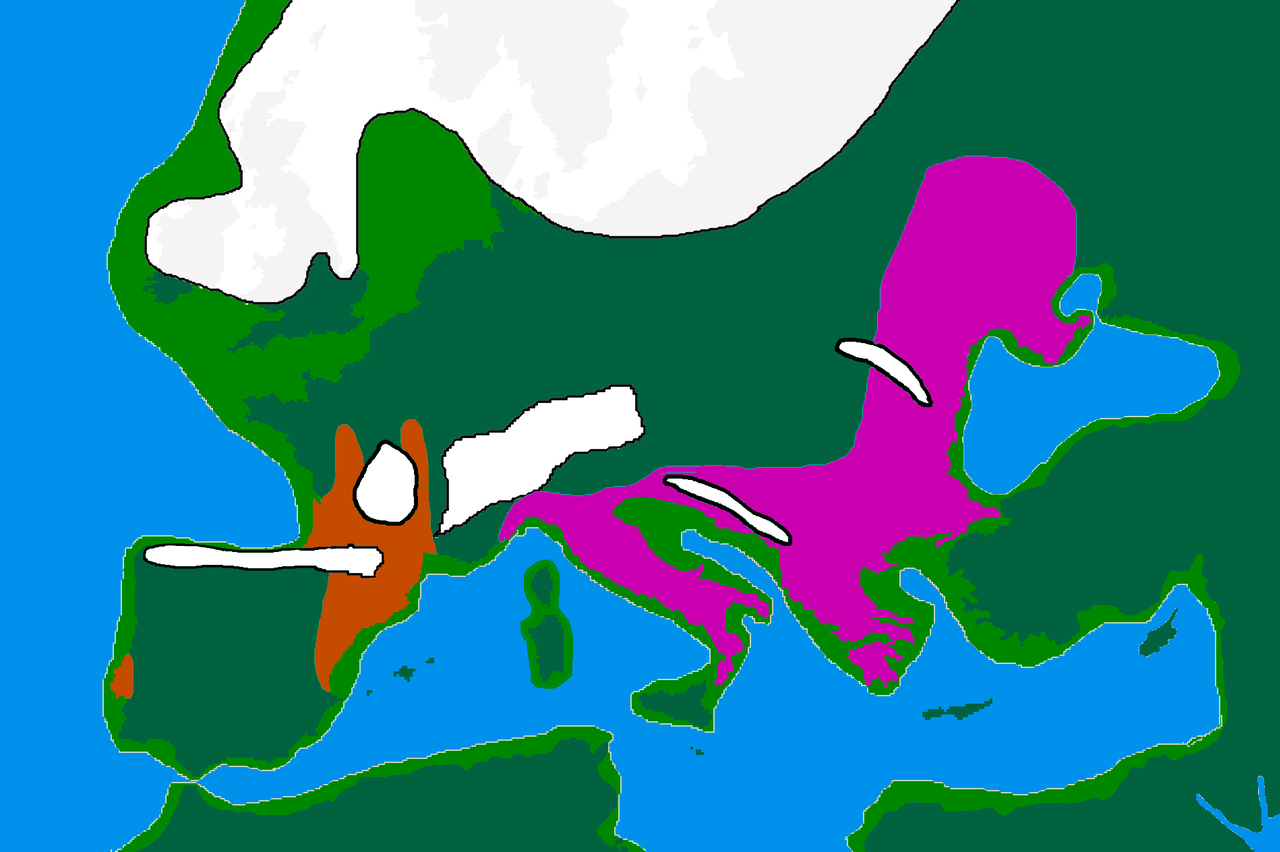
Гаплогруппа I, скорее всего, возникла в Европе, [1] [2] и до сих пор ее находят в палеолитических стоянках по всей Европе, но не за ее пределами. [3] Она отделилась от общего предка IJ* около 43 000 лет назад. [4] Ранние свидетельства существования гаплогруппы J были обнаружены на Кавказе и в Иране. [3] Кроме того, живые примеры предшественника гаплогруппы IJ* были обнаружены только в Иране, среди мазендерани и этнических персов из Фарса . [5] Это может указывать на то, что IJ возникла в Юго-Западной Азии .
Первоначально самым древним найденным образцом был Paglicci133 из Италии , возраст которого составляет не менее 31 000 лет [6] , однако в более позднем исследовании эта информация была изменена, и вместо этого самым древним был указан Dolní Věstonice (DV14) из Чешской Республики , возраст которого составляет не менее 30 800 лет [7] .
Гаплогруппа I была обнаружена у нескольких людей, принадлежащих к культуре Граветтиан. Граветтианцы распространились на запад из дальнего угла Восточной Европы, вероятно, России, в Центральную Европу. Они связаны с генетическим кластером, который обычно называют кластером Вестонице. [8] [9] [10]
Происхождение
Распространение кроманьонцевЕвропейские убежища LGM , 20 000 лет до н.э.
Солютрейские и протосолютрейские культуры
Эпиграветтская культура
Имеющиеся данные свидетельствуют о том, что I-M170 предшествовала в областях, в которых она позже стала доминирующей, гаплогруппам K2a (K-M2308) и C1 (гаплогруппа C-F3393). K2a и C1 были обнаружены в старейших секвенированных мужских останках из Западной Евразии (датируемых примерно от 45 000 до 35 000 лет до н. э. ), таких как: Усть-Ишимский человек (современная Западная Сибирь) K2a*, Оазе 1 (Румыния) K2a*, Костенки 14 (юго-запад России) C1b и Гойет Q116-1 (Бельгия) C1a. [3] [11] Самая старая найденная I-M170 принадлежит человеку, известному как Кремс WA3 (нижняя Австрия), и датируется примерно 33 000-24 000 лет до н. э. На том же месте были найдены два мальчика-близнеца, оба были отнесены к гаплогруппе I*. [12] [13]
Гаплогруппа IJ существовала на Ближнем Востоке и/или в Европе около 40 000 лет назад. [ необходима цитата ] TMRCA (время до самого последнего общего предка ) для I-M170 было оценено Карафетом и коллегами в 2008 году в 22 200 лет назад с доверительным интервалом от 15 300 до 30 000 лет назад. [4] Это сделало бы событие основания I-M170 приблизительно одновременным с последним ледниковым максимумом (LGM), который длился от 26 500 лет назад до примерно 19 500 лет назад. [14] TMRCA является оценкой времени расхождения субкладов. Рутси и коллеги в 2004 году также отмечают две другие даты для клада, возраст изменения STR и время с момента расхождения популяций. Эти последние две даты примерно связаны и происходят несколько позже расхождения субкладов. Для гаплогруппы I-M170 они оценивают время до вариации STR как 24 000 ± 7 100 лет назад и время до расхождения популяций как 23 000 ± 7 700 лет назад. [1] Эти оценки согласуются с оценками Karafet 2008, цитируемыми выше. Однако Андерхилл и его коллеги вычисляют время до расхождения субкладов I1 и I2 как 28 400 ± 5 100 лет назад, хотя они вычисляют возраст вариации STR I1 всего в 8 100 ± 1 500 лет назад. [15]
Семино (2000) предположил, что первоначальное распространение этой популяции соответствует распространению граветтской культуры. [16] Позднее гаплогруппа, наряду с двумя случаями гаплогруппы C , была обнаружена в человеческих останках, принадлежащих ранее упомянутой граветтской культуре, а также у людей мадленской и азильской культур. [17] Рутси и его коллеги в 2004 году предположили, что каждая из предковых популяций, в которых сейчас доминирует определенный субклад гаплогруппы I-M170, испытала независимое расширение популяции сразу после последнего ледникового максимума. [1]
Пять известных случаев гаплогруппы I в останках людей эпохи верхнего палеолита в Европе делают ее одной из самых частых гаплогрупп того периода. [17] В 2016 году были обнаружены останки охотника-собирателя возрастом 31 210–34 580 лет из пещеры Пагличчи , Апулия, Италия, в которых была обнаружена I-M170. [18] До сих пор только гаплогруппа F* и гаплогруппа C 1b были зарегистрированы, по одному разу каждая, на более старых останках в Европе. Субклад I2 I-M170 является основной гаплогруппой, обнаруженной на мужских останках в мезолитической Европе, примерно до 6000 г. до н. э., когда произошла массовая миграция в Европу анатолийских фермеров, несущих Y-ДНК G2a. [19]
Самые ранние документальные свидетельства об I1 относятся к неолитической Венгрии, хотя она, должно быть, отделилась от I2 в более ранний период времени.
В одном случае гаплогруппа I была обнаружена далеко от Европы, среди 2000-летних останков из Монголии. [20]
Похоже, что отдельные волны перемещения населения повлияли на Юго-Восточную Европу . Роль Балкан как давнего коридора в Европу из Анатолии и/или Кавказа подтверждается общим филогенетическим происхождением обеих гаплогрупп I и J в родительской гаплогруппе IJ (M429). Это общее происхождение предполагает, что субклады IJ проникли на Балканы из Анатолии или Кавказа, некоторое время до последнего ледникового максимума. Впоследствии I и J были распространены в Азии и Европе по дизъюнктивной филогеографической схеме, типичной для «родственных» гаплогрупп. Естественный географический коридор, такой как Балканы, вероятно, использовался позднее членами других субкладов IJ, а также другими гаплогруппами, включая те, которые связаны с ранними европейскими земледельцами.
Существование гаплогруппы IJK – предка обеих гаплогрупп IJ и K (M9) – и ее эволюционное расстояние от других субкладов гаплогруппы F (M89) подтверждает вывод о том, что гаплогруппы IJ и K обе возникли в Юго-Западной Азии. Живые носители F* и IJ* были зарегистрированы на Иранском нагорье . [5]
Распределение
Частоты гаплогруппы I:
Подгруппы
Субклады гаплогруппы I-M170 с их определяющими мутациями по состоянию на 2011 год. [ 86] Актуальные филогенетические деревья, перечисляющие все известные на данный момент субклады I, можно найти на сайтах Y-Full и FamilyTreeDNA.
I-M253 Гаплогруппа I1 (L64, L75, L80, L81, L118, L121/S62, L123, L124/S64, L125/S65, L157, L186, L187, M253, M307.2/P203.2, M450/S109, P30, P40, S63, S66, S107, S108, S110, S111) Типична для популяций Скандинавии и Северо-Западной Европы , с умеренным распространением по всей Восточной Европе. В Анатолии на уровне 1% [87]
Обратите внимание, что названия некоторых подгрупп изменились, поскольку были идентифицированы новые маркеры, а последовательность мутаций стала более ясной.
И-М170
Составной субклад I-M170 содержит особей, являющихся прямыми потомками самых ранних членов гаплогруппы I, не имеющих ни одной из последующих мутаций, которые идентифицируют остальные названные субклады.
Несколько индивидуумов I*, которые не попадают ни в один из известных субкладов, были обнаружены среди лакского народа Дагестана с частотой (3/21), [ 89] , а также в Турции (8/741), Адыгее на Кавказе (2/138) и Ираке (1/176), хотя I-M170 встречается только с очень низкой частотой среди современных популяций этих регионов в целом. Это согласуется с убеждением, что гаплогруппа впервые появилась в Юго-Западной Евразии.
(Ни одно из исследований, из которых были получены приведенные выше цифры, не исключало существующую кладу I2-M438 в целом, а только определенные субклады, поэтому эти предполагаемые случаи I*, возможно, относятся к I2.)
У ныне живущего мужчины- хазарейца из Афганистана также был обнаружен I*, при этом все известные субклады I1 (M253) и I2 (M438) были исключены. [91]
И1-М253
Гаплогруппа I1-M253 (M253, M307, P30, P40) демонстрирует очень четкий градиент частоты с пиковой частотой около 35% среди населения южной Норвегии, юго-западной Швеции и Дании и быстро уменьшающимися частотами к краям исторически германского мира. Заметным исключением является Финляндия, где частота у западных финнов достигает 40%, а в некоторых провинциях, таких как Сатакунта, более 50%. Считается, что I1 стала распространенной в результате эффекта основателя во время скандинавского бронзового века и впоследствии распространилась по всей Европе во время периода переселения народов , когда германские племена мигрировали из южной Скандинавии и северной Германии в другие места Европы. [92]
За пределами Фенноскандии распространение гаплогруппы I1-M253 тесно коррелирует с распространением гаплогруппы I2a2-M436; но среди скандинавов (включая как германские, так и уральские народы региона) почти все Y-хромосомы гаплогруппы I-M170 являются I1-M253. Другой характеристикой скандинавских Y-хромосом I1-M253 является их довольно низкое разнообразие гаплотипов (разнообразие STR): большее разнообразие Y-хромосом гаплогруппы I1-M253 было обнаружено среди французов и итальянцев , несмотря на гораздо более низкую общую частоту гаплогруппы I1-M253 среди современных французских и итальянских популяций. Это, наряду со структурой филогенетического дерева I1-M253, убедительно свидетельствует о том, что большинство ныне живущих мужчин I1 являются потомками изначально небольшой группы репродуктивно успешных мужчин, которые жили в Скандинавии во время скандинавского бронзового века. [93] [94]
И2-М438
Гаплогруппа I2-M438 , ранее I1b , возможно, возникла в Южной Европе (сейчас ее наибольшая частота встречается на западных Балканах и Сардинии ) около 15 000–17 000 лет назад и развилась в три основные подгруппы: I2-M438*, I2a-L460, I2b-L415 и I2c-L596.
I2a1a-M26
Гаплогруппа I2a1a-M26 примечательна своим сильным присутствием на Сардинии. Гаплогруппа I-M170 охватывает примерно 40% всех отцов среди сардинцев , и I2a1a-M26 является преобладающим типом I среди них.
Гаплогруппа I2a1a-M26 практически отсутствует к востоку от Франции и Италии [95] , в то время как она встречается с низкими, но значительными частотами за пределами Сардинии на Балеарских островах , в Кастилии-Леоне , Стране Басков , Пиренеях , на юге и западе Франции и в частях Магриба в Северной Африке, Великобритании и Ирландии. Гаплогруппа I2a1a-M26, по-видимому, является единственным субкладом гаплогруппы I-M170, обнаруженным среди басков , но, по-видимому, встречается с несколько более высокими частотами среди общего населения Кастилии-Леона в Испании и Беарна во Франции, чем среди населения этнических басков. [ необходима цитата ] Мутация M26 встречается у коренных мужчин, населяющих каждый географический регион, где могут быть найдены мегалиты, включая такие отдаленные и культурно разобщенные регионы, как Канарские острова, Балеарские острова, Корсика, Ирландия и Швеция. [95]
Распространение I2a1a-M26 также отражает распространение культур атлантического бронзового века , что указывает на потенциальное распространение через торговлю обсидианом или регулярный морской обмен некоторыми металлургическими продуктами. [95]
I2a1b-M423
Приблизительное распределение частот и дисперсии кластеров гаплогруппы I-P37, предкового «Днепровско-Карпатского» (DYS448=20) и производного «Балканского» (DYS448=19: представленного одним SNP I-PH908), в Восточной Европе по О. М. Утевской (2017).
Гаплогруппа I2a1b-M423 является наиболее частой гаплогруппой Y-хромосомы I-M170 в популяциях Центральной и Восточной Европы, достигая своего пика на Западных Балканах , особенно в Далмации (50–60% [33] ) и Боснии и Герцеговине (до 71%, [96] в среднем 40-50% [33] ). Ее субклад I-L161 имеет большую дисперсию в Ирландии и Великобритании, но общая частота очень низкая (2–3%), в то время как субклад I-L162 имеет самую высокую дисперсию и также высокую концентрацию в Восточной Европе (Украина, Юго-Восточная Польша, Беларусь). [97]
I2a2-M436
Распределение гаплогруппы I2a2-M436 (M436/P214/S33, P216/S30, P217/S23, P218/S32) тесно коррелирует с распределением гаплогруппы I1, за исключением Фенноскандии , что позволяет предположить, что она, вероятно, была распространена по крайней мере в одной из палеолитических популяций убежища, которая также распространяла гаплогруппу I1-M253; отсутствие корреляции между распределениями I1-M253 и I2a2-M436 в Фенноскандии может быть результатом того, что гаплогруппа I2a2-M436 была сильнее затронута в самом раннем заселении этого региона эффектами основателя и генетическим дрейфом из-за ее редкости, поскольку гаплогруппа I2a2-M436 составляет менее 10% от общего разнообразия Y-хромосомы всех популяций за пределами Нижней Саксонии . Гаплогруппа I2a2-M436 была обнаружена у более чем 4% населения только в Германии, Нидерландах, Бельгии, Дании, Англии (не включая Корнуолл ), Шотландии и южных оконечностях Швеции и Норвегии в Северо-Западной Европе; провинциях Нормандия , Мэн , Анжу и Перш на северо-западе Франции; провинции Прованс на юго-востоке Франции; регионах Тоскана , Умбрия и Лацио в Италии; и Молдавии и районе вокруг Рязанской области и Республики Мордовия в Восточной Европе. Один субклад гаплогруппы I2a2-M436, а именно I2a2a1a1-M284, был обнаружен почти исключительно среди населения Великобритании, что было принято как предположение о том, что клад может иметь очень долгую историю на этом острове. Однако примечательно, что распределение гаплогрупп I1-M253 и гаплогрупп I2a2-M436, по-видимому, довольно хорошо коррелирует со степенью исторического влияния германских народов . Точечное присутствие обеих гаплогрупп с низкой частотой в районе исторических регионов Вифиния и Галатия в Турции может быть связано с варяжской гвардией или, скорее, предполагает связь с древними галлами Фракии , несколько племен которых, как записано, иммигрировали в эти части Анатолии по приглашению Никомеда I Вифинского . Это предположение подтверждается недавними генетическими исследованиями в отношении Y-ДНК гаплогруппы I2b2-L38, которые пришли к выводу, что в позднем железном веке произошла некоторая миграция кельтских латенцев через Бельгию на Британские острова, включая северо-восточную Ирландию. [98]
Гаплогруппа I2a2-M436 также встречается примерно у 1% сардинцев и у хазарейцев из Афганистана у 3%. [99]
Характеристики мутации
Технические характеристики U179:
Нуклеотидная замена (rs2319818): G на A
Положение (пара оснований): 275
Общий размер (пар оснований): 220
Вперед 5′→ 3′:ааггггататгакгакгатгатт
Обратный 5′→ 3′:cagctcctcttttcaactctca
Высота
Эта гаплогруппа достигает своей максимальной частоты на Западных Балканах (с самой высокой концентрацией I2 в современной Герцеговине ). Это может быть связано с необычно высокими мужчинами, поскольку сообщается, что те, кто находится в Динарских Альпах, являются самыми высокими в мире, со средним ростом мужчин в диапазоне 180 см (5 футов 11 дюймов) - 182 см (6 футов 0 дюймов) в кантонах Босния , 184 см (6 футов 0 дюймов) в Сараево , 182 см (6 футов 0 дюймов) - 186 см (6 футов 1 дюйм) в кантонах Герцеговины, в основном населенных хорватами . [100] Исследование 2014 года, изучающее корреляцию между гаплогруппами Y-ДНК и ростом, обнаружило корреляцию между гаплогруппами I1 , R1b-U106, I2a1b-M423 и высокими мужчинами. [101] В исследовании были измерены средние показатели роста молодых немецких, шведских, голландских, датских, сербских и боснийских мужчин. Средний рост немецких мужчин составил 180,2 см, шведских мужчин — 181,4 см, голландских мужчин — 183,8 см, датских мужчин — 180,6 см, сербов — 180,9 см, а боснийских хорватов из Герцеговины — 185,2 см.
Barać L, Pericić M, Klarić IM, et al. (Июль 2003 г.). "Y-хромосомное наследие хорватской популяции и ее островных изолятов" (PDF) . Eur. J. Hum. Genet . 11 (7): 535–42. doi : 10.1038/sj.ejhg.5200992 . PMID 12825075. S2CID 15822710. Архивировано из оригинала (PDF) 2012-12-17 . Получено 2005-12-08 .
Беннетт, EA, Прат, S., Пеан, S., Крепен, L., Яневич, A., Пуо, S., ... и Гейгл, EM (2019). Происхождение граветтийцев: геномные данные из 36 000-летнего восточноевропейца. BioRxiv, 685404.
Rootsi, S, Kivisild, T, Benuzzi, G, Help, H, et al. (2019). «Филогеография гаплогруппы I Y-хромосомы выявляет отдельные домены доисторического потока генов в Европе». Am. J. Hum. Genet . 75 (1): 128–137. doi : 10.1086/422196 . hdl : 10400.13/3045 . PMID 12825075. S2CID 2834639.
Генографический проект, National Geographic, Атлас пути человека
^ Львович, Рожанский Игорь (2021). «ОБЗОР ДАННЫХ ИСКОПАЕМОЙ ДНК: ГАПЛОКАРТЫ G И I». Исторический формат (4 (28)): 125–140.
^ abc Fu, Qiaomei; et al. (2016). «Генетическая история ледникового периода в Европе». Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.
^ ab Karafet TM, Mendez FL, Meilerman MB, Underhill PA, Zegura SL, Hammer MF (2008). «Новые бинарные полиморфизмы изменяют форму и повышают разрешение древа гаплогруппы Y-хромосомы человека». Genome Research . 18 (5): 830–8. doi :10.1101/gr.7172008. PMC 2336805. PMID 18385274 .
^ ab Grugni (2012). «Древние миграционные события на Ближнем Востоке: новые подсказки из вариации Y-хромосомы современных иранцев». PLOS ONE . 7 (7): e41252. Bibcode : 2012PLoSO...741252G. doi : 10.1371/journal.pone.0041252 . PMC 3399854. PMID 22815981 .
^ Фу, Цяомей и др. (2016). «Генетическая история Европы ледникового периода». Nature . 534 (7606): 200–5. Bibcode :2016Natur.534..200F. doi :10.1038/nature17993. PMC 4943878 . PMID 27135931.
^ Пост, Козимо; Ю, Хэ; Галичи, Айшин; Ружье, Элен; Кревкер, Изабель; Хуан, Илей; Рингбауэр, Харальд; Рорлах, Адам Б.; Нэгеле, Катрин; Вильяльба-Моуко, Ванесса; Радзевичюте, Рита; Феррас, Тьяго; Стессель, Александр; Тухбатова, Резеда; Друкер, Дороти Г. (01 марта 2023 г.). «Палеогеномика от верхнего палеолита до неолита европейских охотников-собирателей». Природа . 615 (7950): 117–126. дои : 10.1038/s41586-023-05726-0. hdl : 10256/23099 . ISSN 1476-4687.
^ "Подробный просмотр публикаций". fgga.univie.ac.at . Получено 2020-12-15 .
^ Mounier, Aurélien; Heuzé, Yann; Samsel, Mathilde; Vasilyev, Sergey; Klaric, Laurent; Villotte, Sébastien (14.12.2020). "Граветтская черепная морфология и родственные связи между группами людей в европейском верхнем палеолите". Scientific Reports . 10 (1): 21931. Bibcode :2020NatSR..1021931M. doi : 10.1038/s41598-020-78841-x . ISSN 2045-2322. PMC 7736346 . PMID 33318530.
^ Беннетт, Э. Эндрю; Прат, Сандрин; Пеан, Стефан; Крепен, Лоран; Яневич, Александр; Пуо, Саймон; Гранж, Тьерри; Гейгль, Ева-Мария (2 июля 2019 г.). «Происхождение граветтов: геномные данные 36 000-летнего восточноевропейца». bioRxiv : 685404. doi : 10.1101/685404. S2CID 198249005.
^ Сегин-Орландо и др. (2014) «Геномная структура европейцев, датируемая по меньшей мере 36 200 лет назад»
^ Тешлер-Никола, Мария; Фернандес, Даниэль; Гендель, Марк; Эйнвегерер, Томас; Саймон, Ульрих; Нойгебауэр-Мареш, Кристина; Тангл, Стефан; Хеймель, Патрик; Добсак, Тони; Рецманн, Аника; Прохаска, Томас (06 ноября 2020 г.). «Древняя ДНК обнаруживает монозиготных новорожденных близнецов из верхнего палеолита». Коммуникационная биология . 3 (1): 650. дои : 10.1038/s42003-020-01372-8 . ISSN 2399-3642. ПМЦ 7648643 . ПМИД 33159107.
^ Clark PU, Dyke AS, Shakun JD и др. (август 2009 г.). «Последний ледниковый максимум». Science . 325 (5941): 710–4. Bibcode :2009Sci...325..710C. doi :10.1126/science.1172873. PMID 19661421. S2CID 1324559.
^ PA Underhill, NM Myres, S. Rootsi, CT Chow, AA Lin, RP Otillar, R. King, LA Zhivotovsky, O. Balanovsky, A. Pshenichnov, KH Ritchie, LL Cavalli-Sforza, T. Kivisild, R. Villems, SR Woodward, Новые филогенетические связи для гаплогруппы I Y-хромосомы: переоценка ее филогеографии и предыстории, в P. Mellars, K. Boyle, O. Bar-Yosef и C. Stringer (редакторы), Переосмысление эволюции человека (2007), стр. 33–42.
^ Semino O, Passarino G, Oefner PJ, et al. (Ноябрь 2000). «Генетическое наследие палеолитического Homo sapiens sapiens у ныне живущих европейцев: перспектива Y-хромосомы». Science . 290 (5494): 1155–9. Bibcode :2000Sci...290.1155S. doi :10.1126/science.290.5494.1155. PMID 11073453.
^ ab "Palaeolithic DNA from Eurasia". Архивировано из оригинала 2016-10-03 . Получено 5 октября 2016 .
^ «Древняя ДНК раскрывает «генетическую преемственность» между каменным веком и современным населением Восточной Азии». Февраль 2017 г.
^ Keyser-Tracqui, C; Crubézy, E; Ludes, B (август 2003 г.). «Анализ ядерной и митохондриальной ДНК некрополя возрастом 2000 лет в долине Эгьин-Гол в Монголии». Am. J. Hum. Genet . 73 (2): 247–60. doi :10.1086/377005. PMC 1180365. PMID 12858290 .[ мертвая ссылка ]
^ abc ИЗУЧЕНИЕ ГЕНЕТИЧЕСКОЙ СТРУКТУРЫ НАРОДОВ ЗАПАДНОГО КАВКАЗА ПО ДАННЫМ О ПОЛИМОРФИЗМЕ Y-ХРОМОСОМЫ, МИТОХОНДРИАЛЬНОЙ ДНК И АЛУ-ИНСЕРЦИЙ
^ "Комментарии к статье Клёсова 2014 года о еврейской ДНК-генеалогии, стр. 1 из 2 - ДНК левитов". Архивировано из оригинала 2016-11-12 . Получено 2016-10-09 .
^ ab Кутуев Ильдус Альбертович. Генетическая структура и молекулярная филогеография народов кавказа.
^ ВКЛАД ОТДЕЛЬНЫХ ПОЛЕССКИХ ПОПУЛЯЦИЙ И ПОПУЛЯЦИИ БЕЛОРУССКИХ ТАТАР В ГЕНОФОНД НАСЕЛЕНИЯ БЕЛАРУСИ [1]
^ abc Pericic M, Lauc LB, Klaric IM и др. (октябрь 2005 г.). «Высокоточный филогенетический анализ юго-восточной Европы прослеживает основные эпизоды отцовского потока генов среди славянских популяций». Mol. Biol. Evol . 22 (10): 1964–75. doi : 10.1093/molbev/msi185 . PMID 15944443.
^ аб Батталья, Винченца; Форнарино, Симона; Аль-Захери, Надя; Оливьери, Анна; Пала, Мария; Майрес, Натали М; Кинг, Рой Дж; Роотси, Сиири; и др. (24 декабря 2008 г.). «Y-хромосомные свидетельства культурного распространения сельского хозяйства в Юго-Восточной Европе» (PDF) . Европейский журнал генетики человека . 17 (6): 820–30. дои : 10.1038/ejhg.2008.249. ПМК 2947100 . ПМИД 19107149.
^ Карачанак, Сена; Форнарино, Симона; Грюни, Виола; Семино, Орнелла; Тончева, Драга; Галабов, Ангел; Атанасов, Борис (2009). «Y-хромосомные гаплогруппы у болгар». Comptes rendus de l'Académie Bulgare des Sciences . 62 (3): 393–400. ISSN 1310-1331. ИНИСТ 21359873.
^ Карачанак С, Груни В, Форнарино С, Нешева Д, Аль-Захери Н, Батталья В, Каросса В, Йорданов Ю, Торрони А, Галабов А.С., Тончева Д, Семино О (2013). Перейра Л.М. (ред.). «Разнообразие Y-хромосомы у современных болгар: новые сведения об их происхождении». ПЛОС ОДИН . 8 (3): e56779. Бибкод : 2013PLoSO...856779K. дои : 10.1371/journal.pone.0056779 . ПМК 3590186 . ПМИД 23483890.
^ "Болгарская_Y_Таблица.xlsx".
^ D. Primorac; et al. (2022). «Хорватское генетическое наследие : обновленная история Y-хромосомы». Хорватский медицинский журнал . 63 (3): 273–286. doi : 10.3325/cmj.2022.63.273 . PMC 9284021. PMID 35722696.
^ abcdefghijklm Географическая структура генетического ландшафта Y-хромосомы Леванта: контраст между прибрежными и внутренними районами
^ Y-хромосомная вариация в Чешской Республике
^ Альтена Э., Смединг Р., ван дер Гааг, К.Дж. и др. Голландский ландшафт Y-хромосомы. Eur J Hum Genet 28, 287–299 (2020). https://doi.org/10.1038/s41431-019-0496-0
^ ЗОНЕН ВАН АДАМ В НИДЕРЛАНДИИ Генетическая генеалогия: een zoektocht в ДНК-архиве ons
^ «Частоты гаплогруппы Y среди фламандского населения».
^ Лаппалайнен, Туули; Койвумяки, Сату; Салмела, Элина; Хуопонен, Кирси; Систонен, Пертти; Савонтаус, Марья-Лийза; Лаэрмо, Пяйви (19 июля 2006 г.). «Региональные различия среди финнов: взгляд на Y-хромосому». Джин . 376 (2): 207–215. дои : 10.1016/j.gene.2006.03.004. ПМИД 16644145.
^ Рамос-Луис, Э.; Бланко-Вереа, А.; Брион, М.; Ван Хаффель, В.; Карраседо, А.; Санчес-Дис, П. (декабрь 2009 г.). «Филогеография французских мужских линий». Forensic Science International: Серия дополнений по генетике . 2 (1): 439–441. дои : 10.1016/j.fsigss.2009.09.026 . S2CID 85134429.
^ Kayser, Manfred; Lao, Oscar; Anslinger, K; Augustin, Christa; Bargel, G.; Edelmann, J; Elias, Sahar; Heinrich, M; Henke, Jürgen (2005). «Значительная генетическая дифференциация между Польшей и Германией следует за современными политическими границами, как показал анализ Y-хромосомы». Human Genetics . 117 (5): 428–443. doi :10.1007/s00439-005-1333-9. ISSN 0340-6717. PMID 15959808. S2CID 11066186.
^ Санчес, Дж. Дж.; Бёрстинг, К.; Эрнандес, А.; Менгель-Йоргенсен, Дж.; Морлинг, Н. (апрель 2004 г.). «Гаплогруппы SNP Y-хромосомы у датчан, гренландцев и сомалийцев». Серия международных конгрессов . 1261 : 347–349. doi :10.1016/S0531-5131(03)01635-2.
^ Наличие трех различных отцовских линий среди северных индийцев: исследование 560 Y-хромосом (2009)
^ ab Влияние истории, географии и религии на генетическую структуру: марониты в Ливане
^ Тамбетс, К.; Рутси, С.; Кивисилд, Т.; и др. (апрель 2004 г.). «Западные и восточные корни саамов — история генетических «выбросов», рассказанная митохондриальной ДНК и Y-хромосомами». Am. J. Hum. Genet . 74 (4): 661–82. doi :10.1086/383203. PMC 1181943. PMID 15024688 .
^ AY Хромосомная перепись Британских островов
^ Однородительские маркеры в Италии выявляют генетическую структуру, обусловленную полом, и различные исторические слои
^ Клинальные паттерны человеческого Y-хромосомного разнообразия в континентальной Италии и Греции определяются дрейфом и эффектом основателя.
^ Следы забытых исторических событий в горных общинах Центральной Италии: генетическое понимание
^ Однородительские маркеры современного населения Италии раскрывают подробности его доримского наследия
^ Генетическая структура современных изолированных популяций Южного Тироля, выявленная с помощью анализа полиморфизмов Y-хромосомы, мтДНК и Alu
^ Различия в миграциях греков и североафриканцев на Сицилию подтверждаются генетическими данными Y-хромосомы.
^ Изоляты в коридоре миграций: высокоразрешающий анализ вариаций Y-хромосомы в Иордании
^ Вестник Московского университета. Серия XXIII АНТРОПОЛОГИЯ № 1/2014: 96–101СВЯЗЬ ИЗМЕНЧИВОСТИ И ХРОМОСОМЫ И РОДОВОЙ СТРУКТУРЫ: ГЕНОФОНД СТЕПНОЙ АРИСТОКРАТИИИ ДУХОВЕНСТВА КАЗАХОВ
^ ab Y-хромосомы у иранцев и таджиков
^ ab MtDNA и вариации Y-хромосомы в курдских группах
^ Двойственное происхождение носителей татского языка из Дагестана, как записано в генеалогии однородительских вариантов
^ Плисс и др. Y-хромосомные линии латышей в контексте генетической изменчивости Восточно-Балтийского региона
^ Разнообразие Y-хромосомы в Ливане структурировано недавними историческими событиями
^ Отцовские линии в Ливии, выведенные из гаплогрупп Y-хромосомы
^ Данные Y-хромосомы об ограниченном греческом вкладе в пуштунское население Пакистана (2006)
^ Микрофилогеографическая и демографическая история португальских мужских родословных
^ Мартинес-Круз Б, Иоана М, Калафель Ф, Арауна ЛР, Санс П, Ионеску Р, Боэнгиу С, Калайджиева Л, Памджав Х, Макух Х, Плантинга Т, ван дер Меер Дж.В., Комас Д., Нетеа М.Г. (2012). Кивисилд Т (ред.). «Анализ Y-хромосомы у лиц, носящих имя Басараб первой династии валашских королей». ПЛОС ОДИН . 7 (7): е41803. Бибкод : 2012PLoSO...741803M. дои : 10.1371/journal.pone.0041803 . ПМК 3404992 . ПМИД 22848614.
^ Разнообразие Y-хромосомы в Саудовской Аравии и его связь с близлежащими регионами.
^ Згонянин Д., Альгафри Р., Антов М., Стоилькович Г., Петкович С., Вукович Р., Драшкович Д. (ноябрь 2017 г.). «Генетическая характеристика 27 локусов Y-STR с помощью набора Yfiler ® Plus в популяции Сербии». Международная судебно-медицинская экспертиза. Генетика . 31 : е48–е49. doi :10.1016/j.fsigen.2017.07.013. ПМИД 28789900.
^ Mihajlovic, Milica; Tanasic, Vanja; Markovic, Milica Keckarevic; Kecmanovic, Miljana; Keckarevic, Dusan (2022-11-01). "Распределение гаплогрупп Y-хромосомы в сербских группах населения, происходящих из исторически и географически значимых отдельных частей Балканского полуострова". Forensic Science International: Genetics . 61 : 102767. doi : 10.1016/j.fsigen.2022.102767. ISSN 1872-4973. PMID 36037736. S2CID 251658864.
^ Генетическая структура словацкого населения, выявленная с помощью полиморфизма Y-хромосомы
^ АНАЛИЗ Й-ДНК ХАПЛОТИПОВ СЛОВЕНЦЕВ
^ Адамс и др. Генетическое наследие религиозного разнообразия и нетерпимости: отцовские линии христиан, иудеев и мусульман на Пиренейском полуострове
^ Изменчивость Y-хромосомы среди суданцев: ограниченный поток генов, соответствие языку, географии и истории
^ Карлссон, Андреас О; Валлерстром, Томас; Готерстром, Андерс; Холмлунд, Гунилла (2006). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива». Европейский журнал генетики человека . 14 (8): 963–970. doi : 10.1038/sj.ejhg.5201651 . PMID 16724001.
^ Rootsi S, Magri C, Kivisild T и др. (июль 2004 г.). «Филогеография гаплогруппы Y-хромосомы I-M170 выявляет отдельные домены доисторического потока генов в Европе». Am. J. Hum. Genet . 75 (1): 128–37. doi :10.1086/422196. PMC 1181996 . PMID 15162323.
^ "Шведская база данных гаплогрупп (Stats haplopie)". Архивировано из оригинала 2016-10-10 . Получено 2016-10-08 .
^ Распределение Y-хромосомы среди популяций на северо-западе Китая выявляет значительный вклад скотоводов Центральной Азии и меньшее влияние западных евразийцев (2010)
^ Bosch, E.; Calafell, F.; Gonzalez-Neira, A.; Flaiz, C; Mateu, E; Scheil, HG; Huckenbeck, W; Efremovska, L; et al. (2006). «Отцовские и материнские линии на Балканах демонстрируют однородный ландшафт, несмотря на языковые барьеры, за исключением изолированных аромунов». Annals of Human Genetics . 70 (Pt 4): 459–87. doi :10.1111/j.1469-1809.2005.00251.x. PMID 16759179. S2CID 23156886.
^ Лаппалайнен, Т.; Лайтинен, В.; Салмела, Э.; Андерсен, П.; Хуопонен, К.; Савонтаус, М.-Л.; Лаэрмо, П. (май 2008 г.). «Миграционные волны в регион Балтийского моря». Энн. Хм. Жене . 72 (Часть 3): 337–48. дои : 10.1111/j.1469-1809.2007.00429.x . PMID 18294359. S2CID 32079904.
^ ISOGG 2011
^ Cinnioğlu, Cengiz; King, Roy; Kivisild, Toomas; Kalfoğlu, Ersi; Atasoy, Sevil; Cavalleri, Gianpiero L.; Lillie, Anita S.; Roseman, Charles C.; Lin, Alice A. (январь 2004 г.). «Раскопки слоев гаплотипа Y-хромосомы в Анатолии». Human Genetics . 114 (2): 127–148. doi :10.1007/s00439-003-1031-4. ISSN 0340-6717. PMID 14586639. S2CID 10763736.
^ "ISOGG 2017 Y-ДНК Гаплогруппа I". isogg.org .
^ ab Caciagli, Laura; Bulayeva, Kazima; Bulayev, Oleg; Bertoncini, Stefania; Taglioli, Luca; Pagani, Luca; Paoli, Giorgio; Tofanelli, Sergio (2009). «Ключевая роль патрилинейного наследования в формировании генетической изменчивости горцев Дагестана». Journal of Human Genetics . 54 (12): 689–694. doi : 10.1038/jhg.2009.94 . ISSN 1434-5161. PMID 19911015.
^ Роотси, Сиири; Магри, Кьяра; Кивисилд, Тоомас; Бенуцци, Джорджия; Помогите, Хела; Бермишева Марина; Кутуев, Ильдус; Барач, Ловорка; Перичич, Марияна (2004). «Филогеография гаплогруппы I Y-хромосомы раскрывает различные области доисторического потока генов в Европе». Американский журнал генетики человека . 75 (1): 128–137. дои : 10.1086/422196. ISSN 0002-9297. ПМК 1181996 . ПМИД 15162323.
^ Cristofaro J, et al. (октябрь 2013 г.). «Афганский Гиндукуш: где сходятся потоки генов Евразийского субконтинента». PLOS ONE . 8 (10): e76748. Bibcode : 2013PLoSO...876748D. doi : 10.1371/journal.pone.0076748 . PMC 3799995. PMID 24204668 .
^ Мальмстрём, Хелена; Гилберт, М. Томас П.; Томас, Марк Г.; Брандстрём, Микаэль; Стора, Ян; Молнар, Петра; Андерсен, Пернилле К.; Бендиксен, Кристиан; Холмлунд, Гунилла; Гётерстрём, Андерс; Виллерслев, Эске (03.11.2009). «Древняя ДНК обнаруживает отсутствие преемственности между неолитическими охотниками-собирателями и современными скандинавами». Current Biology . 19 (20): 1758–1762. doi : 10.1016/j.cub.2009.09.017 . ISSN 0960-9822. PMID 19781941.
^ Карлссон, Андреас О.; Валлерстрём, Томас; Гётерстрём, Андерс; Холмлунд, Гунилла (август 2006 г.). «Разнообразие Y-хромосомы в Швеции – долгосрочная перспектива». European Journal of Human Genetics . 14 (8): 963–970. doi : 10.1038/sj.ejhg.5201651 . ISSN 1476-5438. PMID 16724001.
^ abc Rootsi; et al. "Филогеография гаплогруппы I Y-хромосомы выявляет отдельные домены доисторического потока генов в Европе, рисунок 1" (PDF) . Архивировано из оригинала (PDF) 2007-03-03.
^ Марьянович Д., Форнарино С., Монтанья С. и др. (ноябрь 2005 г.). «Заселение современной Боснии и Герцеговины: гаплогруппы Y-хромосомы в трех основных этнических группах». Ann. Hum. Genet . 69 (Pt 6): 757–63. doi :10.1111/j.1529-8817.2005.00190.x. PMID 16266413. S2CID 36632274.[ мертвая ссылка ]
^ О. М. Утевская (2017). Генофонд українців за различными последовательностями генетических маркеров: поход и смесь по европейскому генетическому пространству (PhD) (на украинском языке). Национальный исследовательский центр радиационной медицины Национальной академии наук Украины . стр. 219–226, 302.
^ МакЭвой и Брэдли, Брайан П. и Дэниел Г. (2010). Кельты с Запада Глава 5: Ирландская генетика и кельты . Oxbow Books, Оксфорд, Великобритания. стр. 117. ISBN978-1-84217-410-4.
^ Хабер, Марк; Платт, Дэниел Э.; Ашрафиан Бонаб, Мазиар; Юханна, Соня С.; Сория-Эрнанц, Дэвид Ф.; Мартинес-Крус, Бегонья; Дуэи, Бушра; Гассибе-Саббах, Мишелла; Рафатпанах, Хошанг; Ганбари, Мохсен; Кит, Джон; Балановский Олег; Уэллс, Р. Спенсер; Комас, Дэвид; Тайлер-Смит, Крис; Заллуа, Пьер А. (2012). «Этнические группы Афганистана имеют общее Y-хромосомное наследие, структурированное историческими событиями». ПЛОС ОДИН . 7 (3): e34288. Бибкод : 2012PLoSO...734288H. doi : 10.1371/journal.pone.0034288 . PMC 3314501. PMID 22470552 .
^ Грагрубер, Павел; Попович, Стево; Бокувка, Доминик; Давидович, Иван; Гржебичкова, Сильва; Ингрова, Павлина; Потпара, Предраг; Прце, Стипан; Страчарова, Никола (2017). «Горы гигантов: антропометрическое обследование молодых мужчин в Боснии и Герцеговине». Королевское общество открытой науки . 4 (4): 161054. Бибкод : 2017RSOS....461054G. дои : 10.1098/rsos.161054. ПМЦ 5414258 . ПМИД 28484621.
^ Grasgruber, P.; Cacek, J.; Kalina, T.; Sebera, M. (2014-12-01). «Роль питания и генетики как ключевых факторов положительной тенденции роста». Economics & Human Biology . 15 : 81–100. doi : 10.1016/j.ehb.2014.07.002 . ISSN 1570-677X. PMID 25190282.
Внешние ссылки
На Викискладе есть медиафайлы по теме Гаплогруппа I Y-ДНК .
Филогенетическое дерево и карты распространения
Y-ДНК гаплогруппа I-M170 и ее субклады по версии Международного общества генетической генеалогии (ISOGG) 2013 г.
Филогеография Y-хромосомы гаплогруппы I
Частотное распределение Y-ДНК гаплогруппы I и ее субкладов – с видеоуроком
Частота и дисперсия I1b (теперь считается I2a2-M26)
Карта «I1a» (теперь считается I-M253)
Карта «I1b», архивированная 01.05.2015 на Wayback Machine (теперь считается I2a-P37.2)
Карта «I1c», архив 2008-08-16 на Wayback Machine (теперь считается I2b-M223)
Вспомнили дерево гаплогруппы I (К. Нордтведт, 2011).
Проекты
Проект I в FTDNA
Проект I1 в FTDNA
Проект I2* в FTDNA
Проект I2a в FTDNA
Проект I2b в FTDNA
Проект I2b2 L38+ в FTDNA
Скандинавский генеалогический проект yDNA в FTDNA
Генеалогический проект Финляндии в FTDNA
Другой
Изучение Y-гаплогруппы I и модальных гаплотипов
Консорциум Y-хромосомы (YCC)
Пример гаплотипов из I1* "y-кластера"
Страница YCC Haplogroup I – I1a (теперь считается I-M253), I1b (теперь считается I-P37.2) и I1c (теперь считается I-M223)
Предсказатель субклада Haplo-I Архивировано 2022-04-02 на Wayback Machine