Копытные ( / ˈ ʌ ŋ ɡ j ʊ l eɪ t s , - ɡ j ə - , - l ɪ t s , - l ə t s / UNG -гюу-лайтс, -gyə-, -lits, -ləts ) являются членами разнообразной клады Euungulata («настоящие копытные»), которая в основном состоит из крупных млекопитающих с копытами . Когда-то часть клады «Ungulata» вместе с кладой Paenungulata , «Ungulata» с тех пор была определена как полифилетическая и, следовательно, недействительная клада на основе молекулярных данных. В результате в 2001 году настоящие копытные были реклассифицированы в более новую кладу Euungulata внутри клады Laurasiatheria , а Paenungulata была реклассифицирована в далекую кладу Afrotheria . [4] Современные копытные делятся на два отряда: Perissodactyla, включая лошадей , носорогов и тапиров ; и парнокопытные, включая крупный рогатый скот , антилоп , свиней , жирафов , верблюдов , овец , оленей и бегемотов , среди других. Китообразные, такие как киты , дельфины и морские свиньи , также классифицируются как парнокопытные, хотя у них нет копыт. Большинство наземных копытных используют копытные кончики пальцев ног, чтобы поддерживать вес своего тела, когда они стоят или двигаются. Два других отряда копытных, Notoungulata и Litopterna , обитающие в Южной Америке, вымерли в конце плейстоцена, около 12 000 лет назад.
Этот термин примерно означает «копытное» или «копытное животное». В качестве описательного термина «копытное» обычно исключает китообразных, поскольку они не обладают большинством типичных морфологических характеристик других копытных, но недавние открытия показывают, что они также произошли от ранних парнокопытных . [5] Копытные, как правило, травоядны, и многие из них используют специализированные кишечные бактерии , позволяющие им переваривать целлюлозу, хотя некоторые представители могут отклоняться от этого: несколько видов свиней и вымершие энтелодонты всеядны, в то время как китообразные и вымершие мезонихи плотоядны.
Копытное происходит от позднелатинского прилагательного ungulatus , «копытный». Ungulatus — уменьшительная форма латинского unguis , «ноготь» (ноготь на пальце руки, ногте). [6]
Euungulata — клада (или, в некоторых систематиках, большой отряд) млекопитающих. Два современных отряда копытных — Perissodactyla (непарнокопытные) и Artiodactyla (парнокопытные) . Hyracoidea (даманы) , Sirenia (морские коровы) (дюгони и ламантины) и Proboscidea (слоны) в прошлом были сгруппированы в кладу «Ungulata», которая позже оказалась полифилетической и теперь недействительной кладой. Три отряда Paenungulata теперь считаются кладой и сгруппированы в кладу Afrotheria, а Euungulata теперь сгруппированы в кладу Laurasiatheria . [1]
В 2009 году морфологические [7] [8] [9] [10] и молекулярные [11] [12] исследования показали, что трубкозубы, даманы, морские коровы и слоны были более тесно связаны друг с другом, а также с сенгисами , тенреками и слонами. золотые кроты , чем непарнокопытные и парнокопытные, и образуют кладу Afrotheria . Слоны, морские коровы и даманы были сгруппированы вместе в кладе Paenungulata , в то время как трубкозуб считался либо их близким родственником, либо близким родственником сенгиса в кладе Afroinsectiphilia . [13] Это яркий пример конвергентной эволюции . [14]
В настоящее время существует некоторый спор относительно того, является ли эта меньшая Euungulata кладистической (основанной на эволюции) группой, или просто фенетической группой ( форма таксона ) или народным таксоном (похожим, но не обязательно родственным). Некоторые исследования действительно обнаружили, что мезаксонские копытные и параксонские копытные образуют монофилетическую линию, [15] [16] [17] тесно связанную либо с Ferae ( хищниками и панголинами ) [18] [19] из клады Fereuungulata , либо с к летучим мышам . [20] Другие исследования показали, что эти два отряда не так уж тесно связаны, поскольку некоторые считают непарнокопытных близкими родственниками летучих мышей и Ferae у Pegasoferae [21] , а другие считают парнокопытных близкими родственниками летучих мышей. [22]






Ниже приведена упрощенная систематика (при условии, что копытные действительно образуют естественную группу) с существующими семействами в порядке взаимоотношений. Имейте в виду, что все еще существовали некоторые серые зоны конфликта, например, в случае с отношениями между семьями пекоранов и семьями усатых китов . См. каждое семейство, чтобы узнать о взаимоотношениях видов, а также о разногласиях в соответствующих статьях.
Ниже приводится общее мнение о филогении семейств копытных. [22] [23]


Perissodactyla и Artiodactyla включают большинство крупных наземных млекопитающих. Эти две группы впервые появились в позднем палеоцене , быстро распространившись среди самых разнообразных видов на многочисленных континентах, и с тех пор развивались параллельно. Некоторые ученые полагали, что современные копытные произошли от эволюционной группы млекопитающих, известной как кондиларты ; [24] самым ранним известным членом группы был крошечный Protungulatum , [25] копытное животное, которое сосуществовало с последним из нептичьих динозавров 66 миллионов лет назад; однако многие авторитеты не считают его настоящим плацентой, не говоря уже о копытном. [26] Загадочные диноцераты были одними из первых крупных травоядных млекопитающих, хотя их точная связь с другими млекопитающими все еще обсуждается, причем одна из теорий заключается в том, что они могут быть просто дальними родственниками ныне живущих копытных; самое последнее исследование выявило, что они входят в состав настоящего сообщества копытных, наиболее близкого к Кароднии . [27]
В Австралии у сумчатых Chaeropus также развились копыта, похожие на копыта парнокопытных, [28] — пример конвергентной эволюции.

Считалось, что непарнокопытные произошли от Phenacodontidae , небольших животных размером с овцу, которые уже демонстрировали признаки анатомических особенностей, которые унаследуют их потомки (например, уменьшение пальцев I и V). [30] К началу эоцена , 55 миллионов лет назад (Mya), они диверсифицировались и распространились, заняв несколько континентов. Лошади и тапиры эволюционировали в Северной Америке; [31] Носороги, по-видимому, развились в Азии от тапироподобных животных, а затем колонизировали Америку в среднем эоцене (около 45 млн лет назад). Из примерно 15 семей выживают только три (McKenna and Bell, 1997; Hooker, 2005). Эти семьи были очень разнообразны по форме и размеру; в их число входили огромные бронтотерии и причудливые халикотерии . Самый крупный непарнокопытный, азиатский носорог по имени Paraceratherium , достигал 15 тонн (17 тонн), что более чем в два раза превышало вес слона . [32]
В ходе кладистического исследования было обнаружено, что антракобуниды и десмостилии - две линии, которые ранее были классифицированы как афротерийцы (точнее, ближе к слонам) - были классифицированы как клада, тесно связанная с непарнокопытными. [2] Демостилии были крупными четвероногими амфибиями с массивными конечностями и коротким хвостом. [33] [ отсутствует длинная цитата ] Они вырастали до 1,8 метра (6 футов) в длину и, как считалось, весили более 200 килограммов (440 фунтов). Их окаменелости были известны от северной части Тихоокеанского региона , [34] от южной Японии через Россию , от Алеутских островов и тихоокеанского побережья Северной Америки до южной оконечности Нижней Калифорнии . Их форма зубов и скелета позволяет предположить, что десмостилии были водными травоядными животными , зависящими от прибрежной среды обитания. Их название связано с их весьма характерными коренными зубами, у которых каждый бугорок был преобразован в полые колонны, так что типичный коренной зуб напоминал скопление труб или, в случае изношенных коренных зубов, вулканы. Они были единственными вымершими морскими млекопитающими.
Южноамериканские меридунгуляты содержат несколько тапироподобных пиротерий и астрапотерий , мезаксонные литоптерны и разнообразные нотунггуляты . В целом считалось, что меридионгуляты произошли от таких животных, как Hyopsodus . [30] Некоторое время их отношения с другими копытными оставались загадкой. Некоторые палеонтологи даже поставили под сомнение монофилию Meridiungulata , предполагая, что пиротерии могут быть более тесно связаны с другими млекопитающими, такими как Embrithopoda (африканский отряд, родственный слонам ), чем с другими южноамериканскими копытными. [35] Недавнее исследование, основанное на костном коллагене, показало, что, по крайней мере, литоптерны и нонггуляты были тесно связаны с непарнокопытными. [36]
Самые старые известные окаменелости , отнесенные к лошадиным, датируются ранним эоценом , 54 миллиона лет назад. Их отнесли к роду Hyracotherium , но типовой вид этого рода теперь не считается членом этого семейства, а остальные виды были выделены в разные роды. Эти ранние лошадиные были животными размером с лису, с тремя пальцами на задних лапах и четырьмя на передних. Они были травоядными браузерами относительно мягких растений и уже были приспособлены к бегу. Сложность их мозга позволяет предположить, что они уже были бдительными и умными животными. [37] Более поздние виды уменьшили количество пальцев на ногах и развили зубы, более подходящие для измельчения травы и другой жесткой растительной пищи.
К началу эоцена носороги отделились от других непарнокопытных . К этому периоду относятся окаменелости Hyracyus eximus , найденные в Северной Америке. Этот маленький безрогий предок больше напоминал тапира или небольшую лошадь, чем носорога. В позднем эоцене возникли три семейства, иногда объединяемые в надсемейство Rhinocerotoidea: Hyracodontidae , Amynodontidae и Rhinocerotidae , создав тем самым взрыв разнообразия, не имеющий себе равных на какое-то время, пока изменения окружающей среды не привели к радикальному уничтожению нескольких видов.
Первые тапириды, такие как гептодон , появились в раннем эоцене. [38] Они выглядели очень похожими на современные формы, но были примерно вдвое меньше и не имели хоботка. Первые настоящие тапиры появились в олигоцене . К миоцену такие роды, как Miotapirus , были почти неотличимы от современных видов. Считалось, что азиатские и американские тапиры разошлись примерно 20–30 миллионов лет назад; а тапиры мигрировали из Северной Америки в Южную Америку около 3 миллионов лет назад в рамках Великого американского обмена . [39]
Непарнокопытные были доминирующей группой крупных наземных животных на протяжении всего олигоцена. Однако с появлением трав в миоцене (около 20 млн лет назад) произошли серьезные изменения: виды парнокопытных с их более сложным желудком смогли лучше адаптироваться к грубой диете с низким содержанием питательных веществ и вскоре приобрели известность. Тем не менее, многие виды непарнокопытных выживали и процветали до позднего плейстоцена (около 10 000 лет назад), когда они столкнулись с давлением человеческой охоты и изменения среды обитания.

Считалось, что парнокопытные произошли от небольшой группы мыщелков Arctocyonidae , которые были неспециализированными, внешне похожими на енотов или медведей, всеядными из раннего палеоцена (около 65–60 миллионов лет назад). У них были относительно короткие конечности, лишенные специализаций, присущих их родственникам (например, уменьшенные боковые пальцы, сросшиеся кости и копыта), [30] и длинные тяжелые хвосты. Их примитивная анатомия делает маловероятным, что они были в состоянии преследовать добычу, но благодаря своим мощным пропорциям, когтям и длинным клыкам они, возможно, были в состоянии одолеть более мелких животных во внезапных атаках. [30] Очевидно, эти млекопитающие вскоре разделились на две отдельные линии: мезонихи и парнокопытные.

Первые парнокопытные выглядели как сегодняшние шевротаны или свиньи: маленькие коротконогие существа, питавшиеся листьями и мягкими частями растений . К позднему эоцену (46 миллионов лет назад) уже сложились три современных подотряда: суина ( группа свиней ); Tylopoda ( отряд верблюдов ); и Ruminantia ( группа коз и крупного рогатого скота ). Тем не менее парнокопытные в то время были далеко не доминирующими: непарнокопытные были гораздо успешнее и гораздо более многочисленными. Парнокопытные выжили в своих нишах, обычно занимая маргинальные места обитания , и предположительно именно в это время у них развилась сложная пищеварительная система , которая позволяла им выживать на пище более низкого качества. В то время как большинство парнокопытных заняли ниши, оставленные несколькими вымершими непарнокопытными, одна линия парнокопытных начала выходить в моря.

Традиционная теория эволюции китообразных заключалась в том, что китообразные были родственниками мезонихий . У этих животных были необычные треугольные зубы, очень похожие на зубы примитивных китообразных. Вот почему ученые долгое время считали, что китообразные произошли от формы мезонихий. Сегодня многие учёные полагают, что китообразные произошли от того же вида, который дал начало гиппопотамам. Эта гипотетическая предковая группа, вероятно, разделилась на две ветви около 54 миллионов лет назад . [5] Одна ветвь эволюционировала в китообразных , возможно, начиная примерно 52 миллиона лет назад с прото-кита Pakicetus и других ранних предков китообразных, известных под общим названием Archaeoceti , которые в конечном итоге подверглись водной адаптации и превратились в полностью водных китообразных . [40] Другой ветвью стали антракотеры , большое семейство четвероногих зверей, самые ранние из которых в позднем эоцене напоминали тощих гиппопотамов со сравнительно маленькими и узкими головами. Все ветви антракотериев, за исключением той, которая развилась в Hippopotamidae , вымерли в плиоцене , не оставив потомков. [41]
Семейство Raoellidae считается самым близким семейством парнокопытных к китообразным. [42] [43] Следовательно, новые теории эволюции китообразных предполагают, что киты и их предки избежали хищничества, а не конкуренции, медленно адаптируясь к океану. [44] [45] [46]

Мезонихи изображались как «волки на копытах» и были первыми крупными хищниками среди млекопитающих, появившимися в палеоцене. [47] У ранних мезонихов на ногах было пять пальцев, которые, вероятно, опирались на землю во время ходьбы ( стопоходное передвижение), но у более поздних мезонихов было четыре пальца, которые заканчивались крошечными копытами на всех пальцах ног, и они все лучше адаптировались к бегу. Подобно бегущим представителям парнокопытных, мезонихи ( например, Pachyaena ) ходили на пальцах ( пальцеходная локомоция). [47] Мезонихи жили очень плохо в конце эпохи эоцена, и только один род, Mongolestes , [48] выжил в эпоху раннего олигоцена , поскольку климат изменился и возникла жесткая конкуренция со стороны более приспособленных креодонтов .


Копытные имели большое разнообразие в ответ на половой отбор и экологические события; у большинства копытных отсутствует ключица . [49] Наземные копытные были по большей части травоядными, а некоторые из них были травоядными . Однако из этого правила были исключения: известно, что свиньи, пекари, бегемоты и дукеры всеядны. Некоторые китообразные были единственными современными копытными, которые были хищниками; усатые киты потребляют значительно меньших по размеру тела животных, таких как мелкие виды рыб и криль ; Зубатые киты, в зависимости от вида, могут потреблять самые разные виды: кальмаров , рыб, акул и других видов млекопитающих, таких как тюлени и другие киты. В экосистемном отношении копытные заселили все уголки планеты, от гор до океанских глубин ; от лугов до пустынь , а некоторые из них были приручены людьми .
Копытные развили специализированные адаптации, особенно в области черепных придатков, зубных рядов и морфологии ног, включая модификацию таранной кости ( одной из костей лодыжки на конце голени) с короткой крепкой головой.
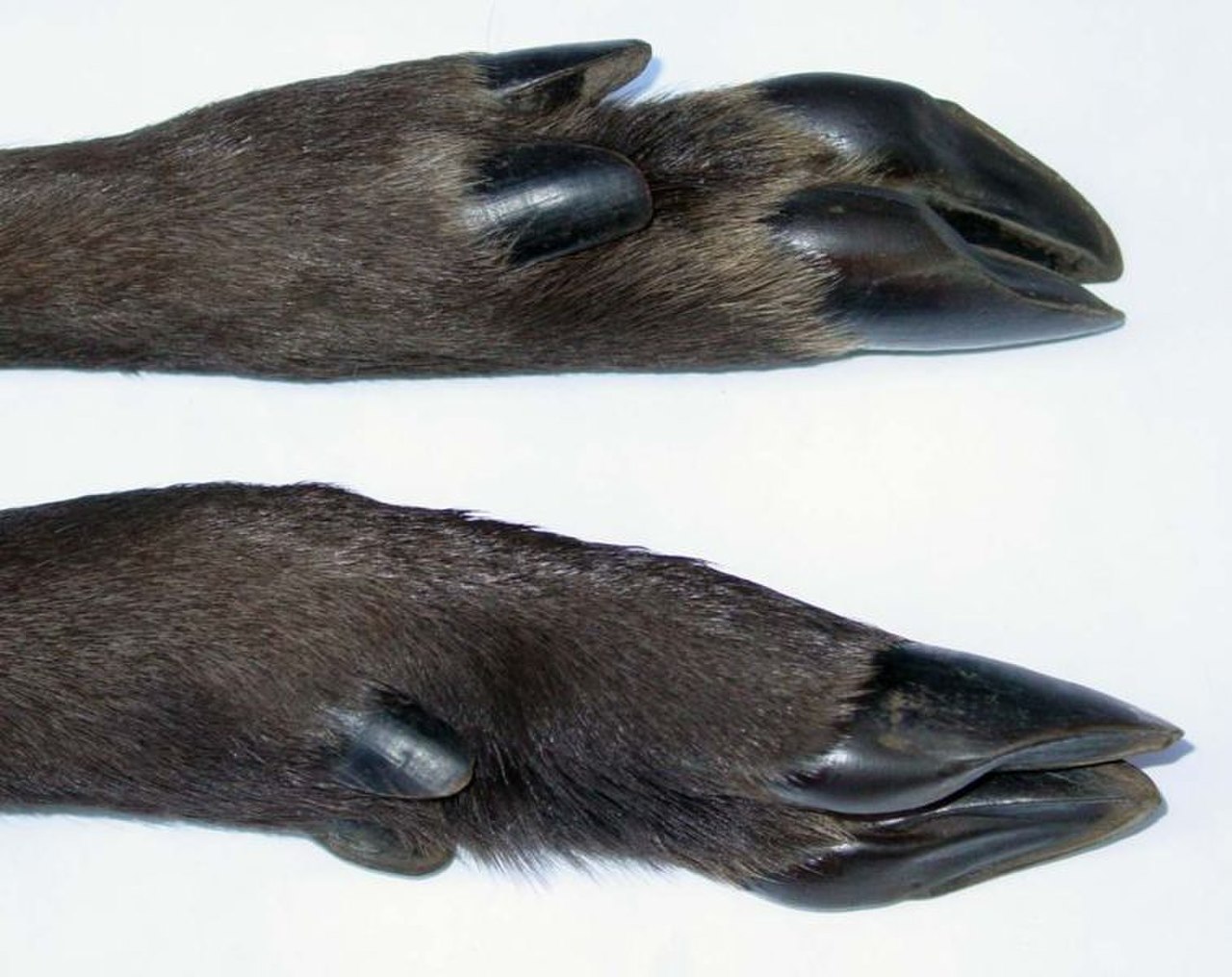
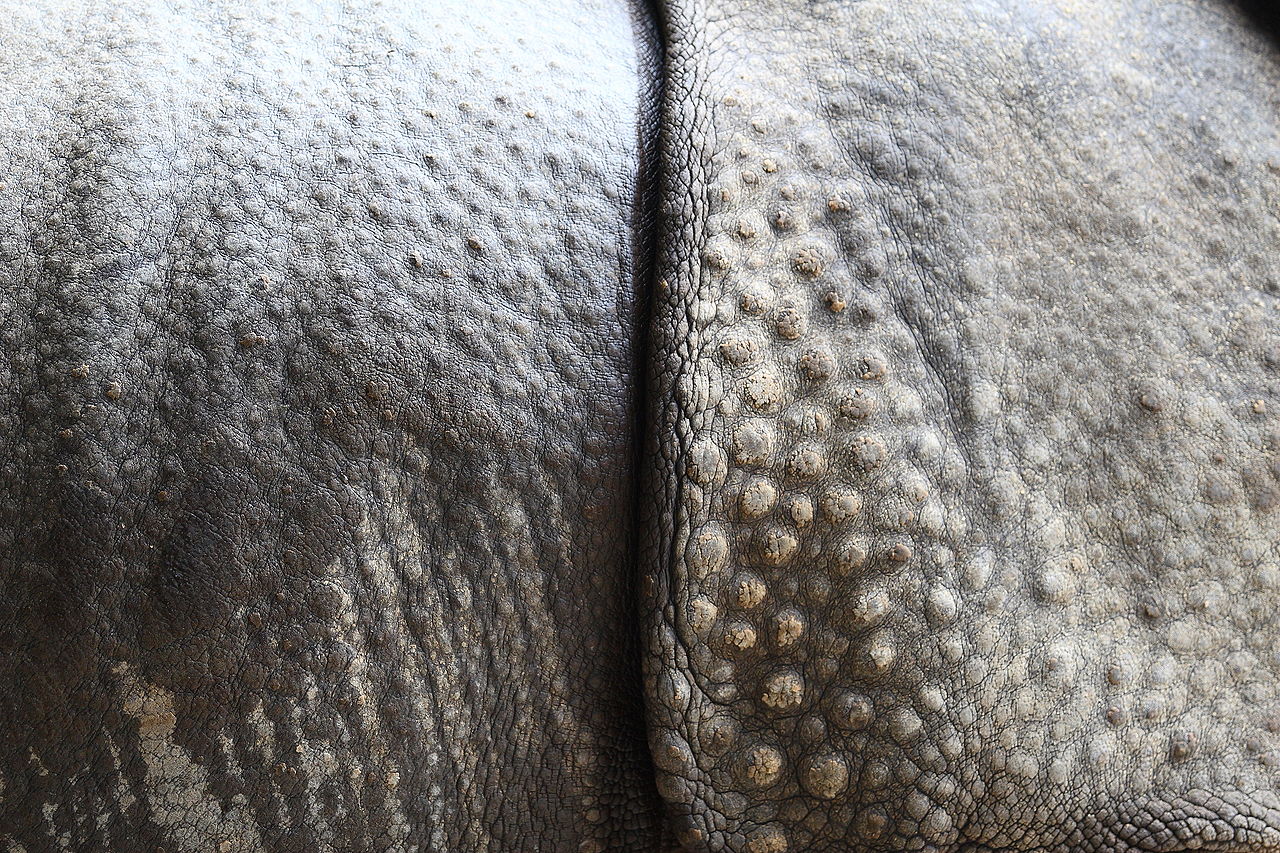
Копыто — кончик пальца копытного млекопитающего , укреплённый толстым роговым ( кератиновым ) покрытием. Копыто состоит из твердой или эластичной подошвы и твердой стенки, образованной толстым ногтем , обернутым вокруг кончика пальца. И подошва, и край копытной стенки обычно выдерживают вес животного. Копыта постоянно растут и постоянно изнашиваются от использования. У большинства современных копытных лучевая и локтевая кости срослись по длине передней конечности; ранние копытные, такие как арктоциониды , не имели этой уникальной структуры скелета. [50] Сращение лучевой и локтевой костей не позволяет копытному вращать переднюю конечность. Поскольку эта скелетная структура не имеет специфической функции у копытных, она считается гомологичной характеристикой, общей для копытных и других млекопитающих. Эта черта могла быть передана от общего предка. Хотя разговорные названия двух отрядов копытных основывались на количестве пальцев у их членов («непарнокопытные» для непарнокопытных и «парнокопытные» для наземных парнокопытных), это не точная причина, по которой они были сгруппированы. У тапиров четыре пальца спереди, однако они принадлежали к отряду «непарнокопытных»; пекари и современные китообразные были представителями отряда «парнокопытных», однако у пекари три пальца спереди, а киты были крайним примером, поскольку у них вместо копыт были ласты. Ученые классифицировали их по распределению веса на пальцы ног.
У периссодактилей мезаксоническая стопа, а это означает, что вес распределяется на третий палец на всех ногах благодаря плоской симметрии стоп. У общего предка произошло сокращение пальцев ног, классическим примером являются лошади с одинарными копытами. В результате у непарнокопытных появилось альтернативное название — почти устаревшая Mesaxonia. Непарнокопытные были не единственной линией млекопитающих, у которой развилась эта черта; у меридунгулятов неоднократно развивались меаксонные стопы.
Наземные парнокопытные имеют параксоническую стопу, то есть вес распределяется на третий и четвертый пальцы всех ног. У большинства этих млекопитающих копыта раздвоены, а два меньших по размеру, известные как прибылые пальцы , расположены выше на ноге. Самые ранние китообразные ( археоцеты ) также обладали этой характеристикой, а также имели таранную кость и кубовидную кость в лодыжке, которые были дополнительными диагностическими признаками парнокопытных. [51]

У современных китообразных передние конечности превратились в грудные плавники , а задние — внутренние и редуцированные. Иногда гены, кодирующие более длинные конечности, заставляют современных китообразных развивать миниатюрные ноги (так называемый атавизм ). Основным способом передвижения является движение вверх и вниз хвостовым плавником, называемым лапой, который используется для движения , а грудные плавники вместе со всей хвостовой частью обеспечивают управление направлением движения. Все современные китообразные по-прежнему сохраняют свои пальцы, несмотря на то, что внешний вид говорит об обратном.
У большинства копытных развились редуцированные клыки и специализированные коренные зубы , в том числе бунодонт (низкие закругленные бугры) и гипсодонт (высокие корончатые зубы). Развитие гипсодонтии представляло особый интерес, поскольку эта адаптация была тесно связана с распространением лугов в миоцене, примерно 25 миллионов лет назад. По мере того как лесные биомы сокращались, пастбища расширялись, открывая новые ниши для млекопитающих. Многие копытные перешли от рациона пасущихся животных к пастбищному рациону, и, возможно, из-за абразивного кремнезема в траве гипсодонтия стала обычным явлением. Однако недавние данные связывают эволюцию гипсодонтии с открытой, песчаной средой обитания, а не с самой травой. Это называется гипотезой «твердости, а не травы» . [52]
У некоторых копытных полностью отсутствуют верхние резцы, вместо них есть зубная подушечка , помогающая при поиске пищи. [53] [54] Его можно найти у верблюдов, жвачных животных и некоторых зубатых китов; Современные усатые киты примечательны тем, что вместо этого у них есть усатые киты , которые отфильтровывают криль из воды. С другой стороны, зубы развились как оружие или средство сексуального проявления, наблюдаемое у свиней и пекари, некоторых видов оленей, кабарги, гиппопотамов, клюворылых китов и нарвала с его длинным клыком. [55]

У копытных развились разнообразные черепные придатки, которые можно найти у цервоидов (за исключением кабарги). У быков и антилоп размер и форма рогов сильно различаются, но основная структура всегда представляет собой пару простых костных выступов без ветвей, часто имеющих спиральную, скрученную или рифленую форму, каждый из которых покрыт постоянной кератиновой оболочкой . Уникальное строение рогов — единственная однозначная морфологическая особенность быков, отличающая их от других пекоран . [56] [57] Развитие рогов у самцов связано с половым отбором, [58] [59] в то время как наличие рогов у самок, вероятно, связано с естественным отбором. [58] [60] Рога самок обычно меньше, чем у самцов, и иногда имеют другую форму. Считается, что рога самок быков развились для защиты от хищников или для выражения территориальности, поскольку нетерриториальные самки, которые могут использовать крипсис для защиты от хищников, часто не имеют рогов. [60]
Рога носорога, в отличие от других рогатых млекопитающих, состоят только из кератина. Эти рога опираются на носовую часть черепа животного.
Рога уникальны для оленьих и встречаются в основном у самцов: единственные самки оленей, имеющие рога, - это карибу и северный олень , чьи рога обычно меньше, чем у самцов. Тем не менее, плодовитые самки других видов оленей способны время от времени производить рога, обычно из-за повышенного уровня тестостерона. [61] Каждый рог растет из точки крепления на черепе, называемой ножкой. Пока рог растет, он покрыт кожей с большим количеством сосудов , называемой бархатом, которая снабжает растущую кость кислородом и питательными веществами. [62] Рога считаются одним из наиболее преувеличенных случаев мужских вторичных половых признаков в животном мире, [63] и растут быстрее, чем любые другие кости млекопитающих. [64] Рост происходит на кончике, первоначально в виде хряща , который затем минерализуется и превращается в кость. Когда рог достигает своего полного размера, бархат теряется, и кость рога отмирает. Эта мертвая костная структура представляет собой зрелый рог. В большинстве случаев кость у основания разрушается остеокластами, и рога со временем отпадают. [62] В результате их быстрого роста рога предъявляют оленям значительную потребность в питательных веществах; таким образом, они могут представлять собой честный сигнал об эффективности метаболизма и способности собирать пищу. [65]
Оссиконы — это роговидные (или рогообразные) выступы, встречающиеся на головах жирафов и самцов окапи . Они похожи на рога антилоп и крупного рогатого скота , за исключением того, что они происходят из окостеневшего хряща [66] и что оссиконы остаются покрытыми кожей и мехом , а не рогом.
Черепные придатки вилорогов уникальны. Каждый «рог» вилорога состоит из тонкой, сплющенной с боков костной пластинки, которая растет из лобных костей черепа, образуя постоянное ядро. Как и у жирафовых, кожа покрывает сердцевину костей, но у вилорога она превращается в ороговевшую оболочку, которая ежегодно сбрасывается и отрастает заново. В отличие от рогов семейства Bovidae, роговые ножны вилорога разветвлены, каждая из ножен имеет направленный вперед зубец (отсюда и название вилорог). Рога у самцов хорошо развиты.
{{cite book}}: CS1 maint: несколько имен: список авторов ( ссылка ){{cite journal}}: CS1 maint: multiple names: authors list (link)