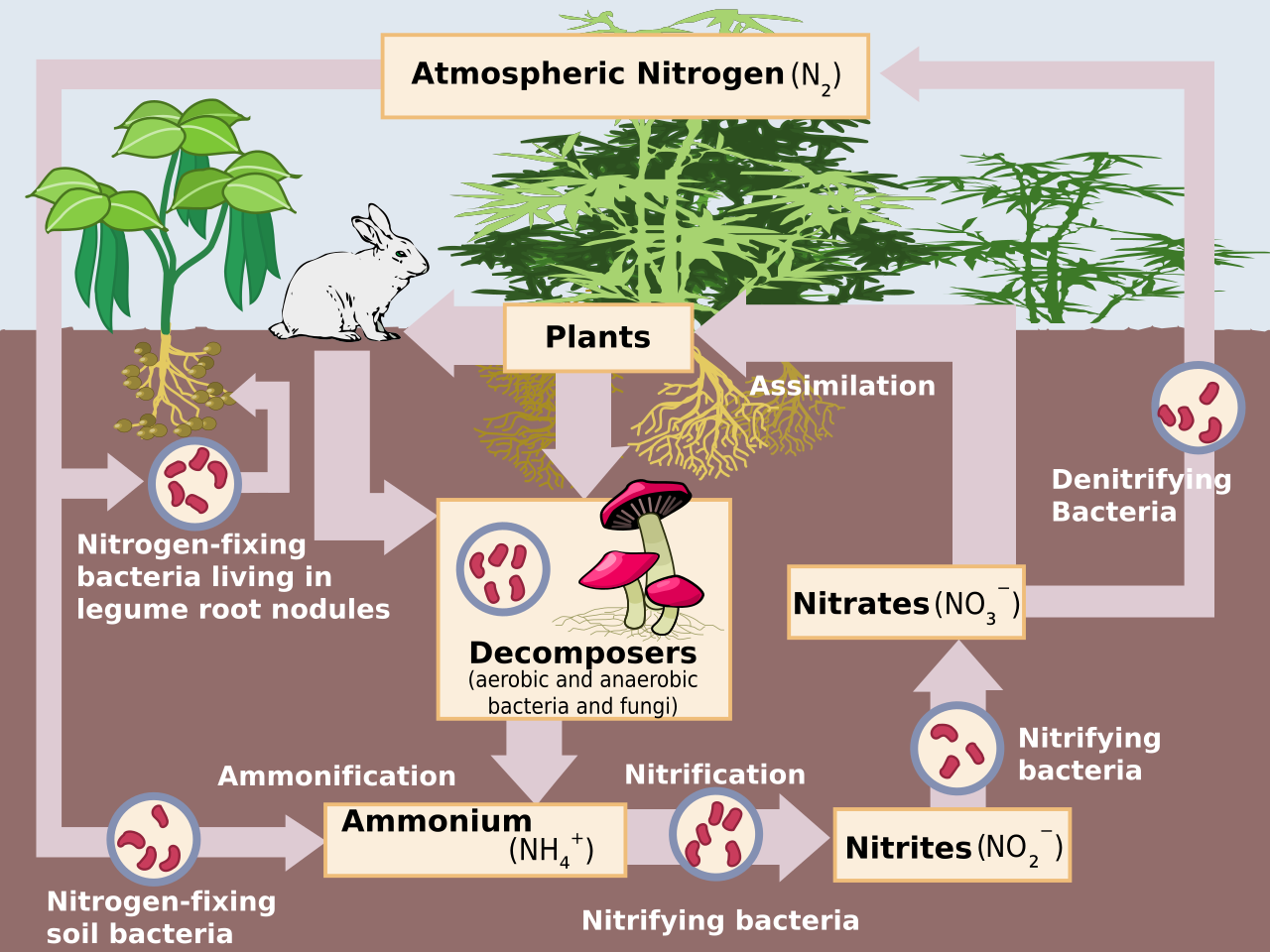
Нитрификация —это биологическое окисление аммиакадо нитрата через промежуточный нитрит . Нитрификация — важный этап круговорота азота в почве . Процесс полной нитрификации может происходить как через отдельные организмы [1], так и полностью внутри одного организма, как у комаммокс -бактерий. Превращение аммиака в нитрит обычно является стадией, лимитирующей скорость нитрификации. Нитрификация — аэробный процесс, осуществляемый небольшими группами автотрофных бактерий и архей .
Процесс нитрификации начинается с первой стадии окисления аммиака, при которой аммиак (NH 3 ) или аммоний (NH 4 + ) превращаются в нитрит (NO 2 - ). Эту первую стадию иногда называют нитрованием. Его осуществляют две группы организмов: аммиакокисляющие бактерии ( АОБ ) и аммиакокисляющие археи ( АОА [2] ).
Аммиакокисляющие бактерии (АОБ) обычно являются грамотрицательными бактериями и принадлежат к бетапротеобактериям и гаммапротеобактериям [3], включая широко изучаемые роды, включая Nitrosomonas и Nitrococcus . Они известны своей способностью использовать аммиак в качестве источника энергии и широко распространены в широком диапазоне сред, таких как почвы, водные системы и очистные сооружения.
AOB обладают ферментами, называемыми монооксигеназами аммиака (AMO), которые катализируют превращение аммиака в гидроксиламин (NH 2 OH), важнейший промежуточный продукт в процессе нитрификации. [4] Эта ферментативная активность чувствительна к факторам окружающей среды, таким как pH, температура и доступность кислорода.
AOB играют жизненно важную роль в нитрификации почвы, что делает их ключевыми игроками в круговороте питательных веществ. Они способствуют превращению аммиака, образующегося при разложении органических веществ или удобрений, в нитрит, который в дальнейшем служит субстратом для нитритокисляющих бактерий (НОБ).
До открытия архей, способных к окислению аммиака, аммиакоокисляющие бактерии (АОБ) считались единственными организмами, способными к окислению аммиака. С момента их открытия в 2005 году [5] культивировались два изолята АОА: Nitrosopumilus maritimus [6] и Nitrososphaera viennensis . [7] При сравнении AOB и AOA, АОА доминирует как в почвах, так и в морской среде, [2] [8] [6] [9] [10] [11] предполагая, что Nitrososphaerota (ранее Thaumarchaeota ) может вносить больший вклад в окисление аммиака. в этих средах. [2]
Кренархеол , который, как принято считать, вырабатывается исключительно АОА (в частности, Nitrososphaerota), был предложен в качестве биомаркера АОА и окисления аммиака. Было обнаружено, что численность кренархеолов связана с сезонным цветением AOA, что позволяет предположить, что было бы целесообразно использовать численность кренархеолов в качестве показателя популяций AOA [12] и, следовательно, окисления аммиака в более широком смысле. Однако открытие Nitrososphaerota, которые не являются обязательными окислителями аммиака [13] , усложняет этот вывод, [14] как и одно исследование, которое предполагает, что кренархеол может продуцироваться морскими эвриархеотами II группы. [15]
Второй этап нитрификации — окисление нитрита в нитрат. Этот процесс иногда называют нитратацией. Окисление нитрита осуществляется нитритокисляющими бактериями ( НОБ ) из таксонов Nitrospirota , [16] Nitrospinota , [17] Pseudomonadota [18] и Chloroflexota . [19] НОБ обычно присутствуют в почве, геотермальных источниках, пресноводных и морских экосистемах.
Окисление аммиака до нитрата в одну стадию внутри одного организма было предсказано в 2006 году [20] и обнаружено в 2015 году у вида Nitrospira inopinata . Чистая культура организма была получена в 2017 году [21] , что представляет собой революцию в нашем понимании процесса нитрификации.
Идея о том, что окисление аммиака до нитрата на самом деле является биологическим процессом, впервые была высказана Луи Пастером в 1862 году. [22] Позже в 1875 году Александр Мюллер, проводя оценку качества воды из колодцев в Берлине , отметил, что аммоний стабилен. в стерилизованных растворах, но нитрифицированы в природных водах. А. Мюллер предположил, что нитрификация при этом осуществляется микроорганизмами. [23] В 1877 году Жан-Жак Шлёзинг и Ахилл Мюнц , два французских агрохимика, работавшие в Париже , доказали, что нитрификация действительно является микробиологически опосредованным процессом, экспериментируя с жидкими сточными водами и искусственной матрицей почвы (стерилизованный песок с порошкообразным мелом). [24] Их выводы были вскоре подтверждены (в 1878 году) Робертом Уорингтоном , который исследовал нитрификационную способность садовой почвы на экспериментальной станции Ротамстед в Харпендене в Англии. [25] Р. Уорингтон также сделал первое наблюдение о том, что нитрификация представляет собой двухэтапный процесс, в 1879 году [26] , что было подтверждено Джоном Манро в 1886 году. [27] Хотя в то время считалось, что двухэтапная нитрификация разделены на отдельные фазы жизни или черты характера одного микроорганизма.
Первый чистый нитрификатор (окисляющий аммиак), скорее всего, был выделен в 1890 году Перси Франклендом и Грейс Франкленд , двумя английскими учеными из Шотландии. [28] До этого Уорингтон , [25] Сергей Виноградский [29] и Франкленды умели лишь обогащать культуры нитрификаторов. Франкланд и Франкланд добились успеха, применив систему серийных разведений с очень низким содержанием инокулята и длительным временем культивирования, исчисляемым годами. Сергей Виноградский в том же году (1890 г.) заявил о выделении чистой культуры [29] , но его культура по-прежнему представляла собой совместную культуру бактерий, окисляющих аммиак и нитрит. [30] С. Виноградскому это удалось всего лишь год спустя, в 1891 году. [31]
Фактически, во время серийных разведений окислители аммиака и окислители нитрита были неосознанно разделены, в результате чего образовалась чистая культура, обладающая только способностью к окислению аммиака. Таким образом, Франкланд и Франкланд заметили, что эти чистые культуры теряют способность выполнять оба этапа. Утрату способности к окислению нитритов наблюдал еще Р. Уорингтон . [26] Культивирование чистого нитритного окислителя произошло позже, в 20 веке, однако невозможно с уверенностью сказать, какие культуры были без примесей, поскольку все теоретически чистые штаммы имеют одинаковые черты (потребление нитрита, производство нитрата). [30]
Оба этапа производят энергию, которая будет связана с синтезом АТФ. Нитрифицирующие организмы являются хемоавтотрофами и используют углекислый газ в качестве источника углерода для роста. Некоторые АОБ обладают ферментом уреазой , который катализирует превращение молекулы мочевины в две молекулы аммиака и одну молекулу углекислого газа. Было показано, что Nitrosomonas europaea , а также популяции обитающих в почве AOB ассимилируют углекислый газ, выделяемый в результате реакции с образованием биомассы через цикл Кальвина , и собирают энергию путем окисления аммиака (другого продукта уреазы) до нитрита. Эта особенность может объяснить усиленный рост АОБ в присутствии мочевины в кислой среде. [32]
В большинстве сред присутствуют организмы, которые завершают оба этапа процесса, образуя нитрат в качестве конечного продукта. Однако возможно создание систем, в которых образуется нитрит ( процесс Шарона ).
Нитрификация важна в сельскохозяйственных системах, где удобрения часто применяются в виде аммиака. Превращение этого аммиака в нитрат увеличивает выщелачивание азота, поскольку нитрат более растворим в воде, чем аммиак.
Нитрификация также играет важную роль в удалении азота из городских сточных вод . Традиционное удаление – это нитрификация с последующей денитрификацией . Стоимость этого процесса заключается в основном в аэрации (подача кислорода в реактор) и добавлении внешнего источника углерода (например, метанола ) для денитрификации.
Нитрификация также может происходить в питьевой воде. В системах распределения, где хлорамины используются в качестве вторичного дезинфицирующего средства, присутствие свободного аммиака может выступать в качестве субстрата для микроорганизмов, окисляющих аммиак. Сопутствующие реакции могут привести к истощению остатков дезинфицирующего средства в системе. [33] Было показано, что добавление хлорит-иона в воду, обработанную хлорамином, контролирует нитрификацию. [34] [35]
Вместе с аммонификацией нитрификация образует процесс минерализации , который относится к полному разложению органического материала с выделением доступных соединений азота. Это восполняет круговорот азота .
В морской среде азот часто является лимитирующим питательным веществом , поэтому круговорот азота в океане представляет особый интерес. [36] [37] Этап цикла нитрификации представляет особый интерес в океане, поскольку он создает нитраты , основную форму азота, ответственную за «новое» производство . Более того, поскольку океан обогащается антропогенным CO 2 , возникающее в результате снижение pH может привести к снижению темпов нитрификации. Нитрификация потенциально может стать «узким местом» в круговороте азота. [38]
Нитрификация, как указано выше, формально представляет собой двухэтапный процесс; на первом этапе аммиак окисляется до нитрита , а на втором этапе нитрит окисляется до нитрата. Разнообразные микробы ответственны за каждый шаг в морской среде. В морской среде известно несколько групп аммиакоокисляющих бактерий (АОБ), в том числе Nitrosomonas , Nitrospira и Nitrosococcus . Все они содержат функциональный ген аммиакмонооксигеназы ( АМО ), который, как следует из названия, отвечает за окисление аммиака. [2] [37] Последующие метагеномные исследования и подходы к культивированию показали, что некоторые Thermoproteota (ранее Crenarchaeota) обладают АМО. Термопротеоты широко распространены в океане, и некоторые виды имеют в 200 раз большее сродство к аммиаку, чем AOB, что контрастирует с предыдущим мнением, что AOB в первую очередь ответственны за нитрификацию в океане. [39] [36] Более того, хотя классически считается, что нитрификация вертикально отделена от первичного производства , поскольку окисление нитрата бактериями ингибируется светом, нитрификация АОА, по-видимому, не ингибируется светом, а это означает, что нитрификация происходит на протяжении всего водяной столб , бросая вызов классическим определениям «нового» и «переработанного» производства . [36]
На втором этапе нитрит окисляется до нитрата. В океанах этот этап не так хорошо изучен, как первый, но известно, что бактерии Nitrospina [17] [40] и Nitrobacter выполняют этот этап в океане. [36]
Нитрификация — это процесс окисления соединений азота (фактически, потеря электронов от атома азота к атомам кислорода ), который поэтапно катализируется рядом ферментов.
ИЛИ
У Nitrosomonas europaea первая стадия окисления (аммиак до гидроксиламина ) осуществляется ферментом аммиакмонооксигеназой (АМО).
Второй этап (гидроксиламин в нитрит) катализируется двумя ферментами. Гидроксиламиноксидоредуктаза (ГАО) превращает гидроксиламин в оксид азота. [41]
Другой неизвестный в настоящее время фермент превращает оксид азота в нитрит.
Третий этап (от нитрита к нитрату) завершается в отдельном организме.
Из-за присущей ей микробной природы нитрификация в почвах сильно зависит от почвенных условий. В целом нитрификация почвы будет происходить с оптимальной скоростью, если условия для микробных сообществ будут способствовать здоровому росту и активности микробов. Почвенные условия, влияющие на скорость нитрификации, включают:
Ингибиторы нитрификации — это химические соединения, замедляющие нитрификацию аммиака , аммонийсодержащих или мочевиносодержащих удобрений , которые вносятся в почву в качестве удобрений. Эти ингибиторы могут помочь снизить потери азота в почве, который в противном случае использовался бы сельскохозяйственными культурами. Ингибиторы нитрификации широко используются: их добавляют примерно к 50% осеннего безводного аммиака в штатах США, таких как Иллинойс. [42] Они обычно эффективны в увеличении извлечения азотных удобрений в пропашных культурах, но уровень эффективности зависит от внешних условий, и их преимущества, скорее всего, будут видны при нормах азота, меньших, чем оптимальные. [43]
Экологические проблемы, связанные с нитрификацией, также способствуют интересу к использованию ингибиторов нитрификации: основной продукт, нитрат , вымывается в грунтовые воды, вызывая токсичность как для людей [44] , так и для некоторых видов дикой природы и способствуя эвтрофикации стоячих вод. Некоторые ингибиторы нитрификации также подавляют выработку метана , парникового газа.
Торможению процесса нитрификации способствуют, прежде всего, отбор и ингибирование/уничтожение бактерий, окисляющих соединения аммиака. Множество соединений, ингибирующих нитрификацию, которые можно разделить на следующие области: активный центр аммонооксигеназы (АМО), механистические ингибиторы и процесс N- гетероциклических соединений . Процесс последнего из трех еще не получил широкого понимания, но он известен. Присутствие АМО было подтверждено на многих субстратах, являющихся ингибиторами азота, таких как дициандиамид , тиосульфат аммония и нитрапирин .
Превращение аммиака в гидроксиламин является первым этапом нитрификации, где АН 2 представляет собой ряд потенциальных доноров электронов.
Эту реакцию катализирует АМО. Ингибиторы этой реакции связываются с активным центром АМО и предотвращают или замедляют процесс. Процессу окисления аммиака АМО придается большое значение в связи с тем, что другие процессы требуют совместного окисления NH 3 для получения восстанавливающих эквивалентов. Обычно его обеспечивает соединение гидроксиламиноксидоредуктаза (НАО), которое катализирует реакцию:
Это требование усложняет механизм торможения. Кинетический анализ ингибирования окисления NH 3 показал, что субстраты АМО демонстрируют кинетику от конкурентной до неконкурентной . Связывание и окисление могут происходить по двум сайтам АМО: у конкурентных субстратов связывание и окисление происходит по сайту NH 3 , а у неконкурентных субстратов — по другому сайту.
Ингибиторы, основанные на механизме действия, можно определить как соединения, которые прерывают нормальную реакцию, катализируемую ферментом. Этот метод осуществляется путем инактивации фермента посредством ковалентной модификации продукта, что в конечном итоге ингибирует нитрификацию. В ходе этого процесса АМО деактивируется, и один или несколько белков ковалентно связываются с конечным продуктом. Обнаружено, что это наиболее заметно в широком диапазоне соединений серы или ацетилена .
Установлено, что серосодержащие соединения, в том числе тиосульфат аммония (популярный ингибитор), производят летучие соединения с сильными ингибирующими эффектами, такие как сероуглерод и тиомочевина .
В частности, примечательным дополнением стал тиофосфорилтриамид, который имеет двойную цель: ингибировать как выработку уреазы , так и нитрификацию. [45] В исследовании ингибирующих эффектов окисления бактериями Nitrosomonas europaea использование тиоэфиров привело к окислению этих соединений до сульфоксидов , где атом S является основным местом окисления АМО. Наиболее сильно это коррелирует с областью конкурентного торможения.

N-гетероциклические соединения также являются высокоэффективными ингибиторами нитрификации и часто классифицируются по кольцевой структуре. Механизм действия этих соединений недостаточно изучен: хотя нитрапирин, широко используемый ингибитор и субстрат АМО, является слабым ингибитором указанного фермента, эффекты указанного механизма не могут напрямую коррелировать со способностью соединения ингибируют нитрификацию. Предполагается, что нитрапирин действует против фермента монооксигеназы внутри бактерий, предотвращая рост и окисление CH 4 /NH 4 . [46] Соединения, содержащие два или три соседних атома N в кольце ( пиридазин , пиразол , индазол ), как правило, оказывают значительно более высокий ингибирующий эффект, чем соединения, содержащие несмежные атомы N или единичные атомы N в кольце ( пиридин , пиррол ). [47] Это говорит о том, что наличие атомов N в кольце напрямую коррелирует с ингибирующим эффектом этого класса соединений.
Некоторые ферментативные ингибиторы нитрификации, такие как нитрапирин, также могут ингибировать окисление метана у метанотрофных бактерий. [48] АМО демонстрирует кинетическую скорость оборота, аналогичную метанмонооксигеназе (ММО), обнаруженной у метанотрофов, что указывает на то, что ММО является катализатором, аналогичным АМО, в целях окисления метана. Более того, метанотрофные бактерии имеют много общего с окислителями NH 3 , такими как Nitrosomonas . [49] Профиль ингибитора дисперсных форм ММО (pMMO) демонстрирует сходство с профилем АМО, что приводит к сходству свойств между ММО у метанотрофов и АМО у автотрофов .

Ингибиторы нитрификации также представляют интерес с экологической точки зрения из-за образования нитратов и закиси азота в процессе нитрификации. Закись азота (N 2 O), хотя ее концентрация в атмосфере намного ниже, чем у CO 2, имеет потенциал глобального потепления примерно в 300 раз больший, чем углекислый газ, и способствует 6% планетарного потепления за счет парниковых газов. Это соединение также известно тем, что катализирует распад озона в стратосфере . [50] Нитраты, токсичное соединение для дикой природы и домашнего скота и продукт нитрификации, также вызывают обеспокоенность.
Почва, состоящая из полианионных глин и силикатов , обычно имеет чистый анионный заряд. Следовательно, аммоний (NH 4 + ) прочно связывается с почвой, а ионы нитрата (NO 3 - ) — нет. Поскольку нитраты более подвижны, они попадают в грунтовые воды через сельскохозяйственные стоки . Нитраты в подземных водах могут влиять на концентрацию поверхностных вод либо посредством прямого взаимодействия подземных и поверхностных вод (например, приток ручьев, родников), либо в результате их извлечения для поверхностного использования. Например, большая часть питьевой воды в Соединенных Штатах поступает из грунтовых вод, но большинство очистных сооружений сбрасывают воду в поверхностные воды.
Дикие животные, такие как амфибии, пресноводные рыбы и насекомые, чувствительны к уровню нитратов и, как известно, вызывают смерть и аномалии развития у пораженных видов. [51] Уровни нитратов также способствуют эвтрофикации — процессу, при котором сильное цветение водорослей снижает уровень кислорода в водоемах и приводит к смерти потребляющих кислород существ из-за аноксии. Считается также, что нитрификация способствует образованию фотохимического смога, приземного озона, кислотных дождей , изменений видового разнообразия и других нежелательных процессов. Кроме того, было показано, что ингибиторы нитрификации подавляют окисление метана (CH 4 ), сильнодействующего парникового газа , до CO 2 . Показано, что и нитрапирин, и ацетилен являются особенно сильными супрессорами обоих процессов, хотя характер их действия неясен.






