Некоторые подвижные одноклеточные простейшие [1] [2]
Одноклеточные протисты составляют обширную, разнообразную группу организмов, которая охватывает практически все среды и места обитания, демонстрируя зверинец форм и видов. Сотни видов инфузорий рода Paramecium [3] или жгутиконосцев Euglena [4] встречаются в морских, солоноватых и пресноводных водоемах; зеленые водоросли Chlamydomonas распространены в почве и пресной воде по всему миру; [5] паразиты из рода Giardia колонизируют кишечник нескольких видов позвоночных. [6] Одной из общих черт этих организмов является их подвижность, имеющая решающее значение для получения питательных веществ и избегания опасности. [7] В процессе эволюции одноклеточные организмы развивались в различных направлениях, и, таким образом, их богатая морфология приводит к широкому спектру способов плавания. [8] [2]
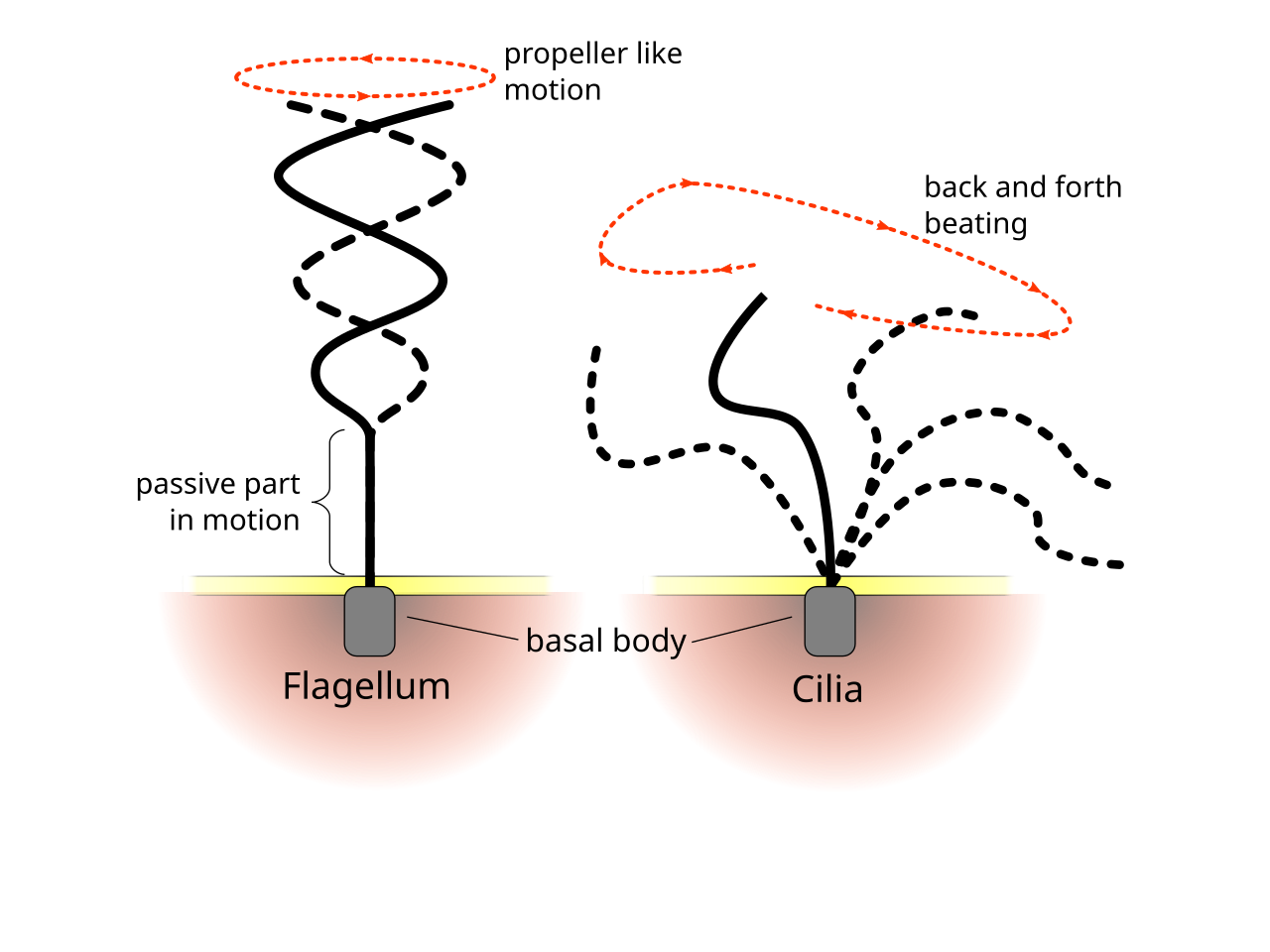
Многие плавающие простейшие приводят в действие хвостоподобные придатки, называемые жгутиками или ресничками , чтобы создать необходимую тягу. [9] Это достигается путем активного создания деформаций вдоль жгутика, что приводит к образованию сложной формы волны. Сама аксонема жгутика представляет собой пучок из девяти пар дублетов микротрубочек , окружающих две центральные микротрубочки, называемых аксонемой 9+2 , [10] и сшивающие динеиновые моторы , приводимые в действие гидролизом АТФ , выполняют механическую работу, способствуя относительному скольжению нитей , что приводит к изгибным деформациям . [2]
Хотя жгутики простейших имеют разнообразие форм и функций, [11] два больших семейства, жгутиконосцы и инфузории , можно различить по форме и характеру биения их жгутиков. [2]
На филогенетическом дереве справа водные организмы (живущие в морской, солоноватой или пресноводной среде) имеют свои ветви, нарисованные синим цветом, а паразитические организмы имеют свои ветви, нарисованные красным цветом. Инфузории обозначены звездочкой после их названий. Для каждого типа, отмеченного жирным шрифтом, репрезентативный организм был изображен рядом с его названием. [2]
Способы передвижения
Жгутиконосцы
Разница в характере биения жгутика и реснички
Жгутики используются как прокариотами (археями и бактериями), так и протистами. Кроме того, как жгутики, так и реснички широко используются в эукариотических клетках (растительных и животных), помимо протистов.
Регулярные паттерны биения эукариотических ресничек и жгутиков генерируют движение на клеточном уровне. Примеры варьируются от движения отдельных клеток, такого как плавание сперматозоидов , до транспортировки жидкости вдоль неподвижного слоя клеток, такого как в дыхательных путях . Хотя эукариотические жгутики и подвижные реснички ультраструктурно идентичны, паттерн биения двух органелл может быть разным. В случае жгутиков движение часто плоское и волнообразное, тогда как подвижные реснички часто выполняют более сложное трехмерное движение с силовым и восстановительным ходом.
Эукариотические жгутики — те, что у животных, растений и протистских клеток — это сложные клеточные выступы, которые хлещут вперед и назад. Эукариотические жгутики классифицируются вместе с эукариотическими подвижными ресничками как ундулиподии [17], чтобы подчеркнуть их отличительную роль волнистых придатков в клеточной функции или подвижности . Первичные реснички неподвижны и не являются ундулиподиями.
Морские жгутиконосцы из родов (слева направо) Cryptaulax , Abollifer , Bodo , Rhynchomonas , Kittoksia , Allas и Metromonas [18]
Жгутиконосцы обычно имеют небольшое количество длинных жгутиков, распределенных вдоль тела, и они приводят их в действие для создания тяги. Набор наблюдаемых последовательностей движений включает плоские волнообразные волны и бегущие спиральные волны, либо от основания к кончику, либо в противоположном направлении. [19] [20] Жгутики, прикрепленные к одному и тому же телу, могут следовать разным схемам биения, что приводит к сложной стратегии локомоции, которая часто также зависит от сопротивления, которое тело клетки оказывает жидкости. [2]
Инфузории
Реснички совершают мощные гребки вперед с помощью напряженного жгутика, за которыми следует относительно медленное возвратное движение с расслабленным жгутиком.
В отличие от жгутиконосцев, движение инфузорий происходит от движения слоя плотно упакованных и коллективно движущихся ресничек, которые представляют собой короткие волосовидные жгутики, покрывающие их тела. В основополагающей обзорной статье Бреннена и Винета (1977) перечислены несколько примеров из обеих групп, подчеркивающих их форму, форму удара, геометрические характеристики и плавательные свойства. [20] Реснички также могут использоваться для транспортировки окружающей жидкости, и их кооперативность может приводить к направленной генерации потока. У высших организмов это может иметь решающее значение для внутренних транспортных процессов, как при цитоплазматическом течении внутри растительных клеток, [21] или транспортировке яйцеклеток из яичника в матку у самок млекопитающих. [22] [2]
Инфузории обычно имеют сотни или тысячи ресничек, которые плотно упакованы вместе в массивы. Как и жгутики, реснички приводятся в действие специализированными молекулярными моторами . Эффективный прямой ход выполняется с помощью жесткого жгутика, за которым следует неэффективный обратный ход, выполняемый с помощью расслабленного жгутика. Во время движения отдельная ресничка деформируется, поскольку она использует силовые удары с высоким трением и восстановительные удары с низким трением. Поскольку на отдельном организме есть несколько ресничек, упакованных вместе, они демонстрируют коллективное поведение в метахрональном ритме . Это означает, что деформация одной реснички совпадает по фазе с деформацией ее соседки, вызывая волны деформации, которые распространяются по поверхности организма. Эти распространяющиеся волны ресничек позволяют организму использовать реснички скоординированным образом для движения. Типичным примером ресничного микроорганизма является парамеция , одноклеточное ресничное простейшее, покрытое тысячами ресничек. Совместное биение ресничек позволяет парамецию двигаться в воде со скоростью 500 микрометров в секунду. [23]
Реснички бьются в многоресничном микропловце [24]
Два изображения в разное время, одно за другим, показывающие разное расположение бьющихся ресничек. Различный цвет ресничек указывает на мгновенную стадию в цикле биения. Движение пловца слева направо
Третьей распространенной формой подвижности клеток простейших является актин -зависимая миграция клеток . Эволюция плавания на основе жгутиков хорошо изучена, и убедительные доказательства указывают на единое эволюционное происхождение эукариотического жгутика, произошедшее до диверсификации современных эукариот. С другой стороны, актин-зависимое ползание использует множество различных молекулярных механизмов, и изучение того, как они эволюционировали, только начинается. [25]
(a) Шестнадцатиклеточная колония. Каждая клетка имеет два жгутика длиной 30–40 мкм. Масштабная линейка составляет 10 мкм. (b) Схема колонии радиусом a: шестнадцать клеток (зеленые), каждая с одним глазком (оранжевая точка). Цис-жгутик находится ближе всего к глазку, транс-жгутик — дальше всего. [27] Жгутики центральных клеток бьются в встречном брассом, в то время как периферические жгутики бьются параллельно. Организация периферических жгутиков по типу вертушки приводит к левостороннему вращению тела со скоростью ω3.
Gonium — род колониальных водорослей , принадлежащих к семейству Volvocaceae . Типичные колонии имеют от 4 до 16 клеток, все одинакового размера, расположенных в плоской пластине, без передне-задней дифференциации. В колонии из 16 клеток четыре находятся в центре, а остальные 12 — по четырем сторонам, по три с каждой стороны. [28]
Начиная с работ Августа Вейсмана по теории зародышевой плазмы в биологии [29] и Джулиана Хаксли по природе индивидуума в эволюционной теории [30] , различные виды зеленых водорослей, принадлежащих к семейству Volvocaceae, были признаны важными в изучении эволюционных переходов от одноклеточной к многоклеточной жизни. С современной биологической точки зрения [31] эта значимость возникает из-за ряда специфических особенностей этих водорослей, включая тот факт, что они являются существующим семейством (что устраняет необходимость изучать микроископаемые ), легкодоступны в природе, изучались с различных точек зрения (биохимической, онтогенетической, генетической) и имели значительные экологические исследования. С точки зрения динамики жидкости [32] их относительно большой размер и простые условия культивирования позволяют проводить точные исследования их подвижности, потоков, которые они создают своими жгутиками, и взаимодействий между организмами, в то время как их высокая степень симметрии упрощает теоретические описания тех же самых явлений. [33] [26]
Поскольку они фотосинтезирующие, способность этих водорослей выполнять фототаксис является центральной для их жизни. Поскольку родословная охватывает от одноклеточных до крупных колониальных форм, ее можно использовать для изучения эволюции многоклеточной координации подвижности. Подвижность и фототаксис подвижных зеленых водорослей были предметом обширной литературы в последние годы, [34] [35] [36] [37] [38] [39] [40] уделяя основное внимание двум крайним случаям: одноклеточным Chlamydomonas и гораздо более крупным Volvox , с видами, состоящими из 1000–50 000 клеток. Chlamydomonas обычно плавает, активируя свои два жгутика брассом, сочетая продвижение и медленное вращение тела. У нее есть глазное пятно, небольшая область, очень чувствительная к свету, [41] [42], которая по-разному активирует два жгутика. [43] Эти реакции адаптивны, в масштабе времени, соответствующем периоду вращения тела клетки, [44] [45] [46] и позволяют клеткам сканировать окружающую среду и плыть к свету. [47] Многоклеточный Volvox демонстрирует более высокий уровень сложности с дифференциацией между внутренними зародышевыми клетками и соматическими клетками, предназначенными для движения. Несмотря на отсутствие центральной нервной системы для координации своих клеток, Volvox демонстрирует точный фототаксис. Это также достигается за счет адаптивной реакции на изменение уровня освещенности, при этом время реакции настроено на период вращения колонии, что создает дифференциальную реакцию между светлой и темной сторонами сфероида. [35] [48] [26]
В свете вышесказанного возникает естественный вопрос: как простейший дифференцированный организм достигает фототаксиса? В родословной Volvocine интересующий нас вид — Gonium. Эта 8- или 16-клеточная колония представляет собой один из первых шагов к настоящей многоклеточности, [49] предположительно произошедшей от одноклеточного общего предка раньше, чем другие водоросли Volvocine. [50] Она также является первой, показавшей клеточную дифференциацию. [26]
16-клеточная колония Gonium , показанная на диаграмме справа, организована в два концентрических квадрата из 4 и 12 клеток соответственно, каждая из которых двужгутиковая, удерживаемая вместе внеклеточным матриксом. [51] Все жгутики направлены в одну сторону: она демонстрирует гораздо более низкую симметрию, чем Volvox, не имея передне-задней симметрии. Тем не менее, она выполняет функции, аналогичные функциям ее одноклеточных и крупных колоний, поскольку она смешивает движение и вращение тела и эффективно плывет к свету. [34] [52] [53] Жгутиковая организация внутренних и периферических клеток сильно отличается: [54] [55] Центральные клетки похожи на Chlamydomonas, с двумя жгутиками, бьющимися в противоположных брассовых гребках, и вносят основной вклад в поступательное движение колонии. Однако у клеток на периферии жгутики бьются параллельно, что близко к клеткам Volvox . [27] Это сводит к минимуму стерические взаимодействия и позволяет избежать пересечения жгутиков друг с другом. [34] Более того, эти жгутики имплантированы под небольшим углом и организованы в виде вертушки [см. рис. 1(b)]: [54] Их биение вызывает левостороннее вращение колонии, выделенное на рис. 1(c) и 1(d) и в дополнительном фильме 1 [29]. Таким образом, структура жгутиков Gonium усиливает его ключевое положение как промежуточного звена в эволюции к многоклеточности и клеточной дифференциации. [26]
Эти небольшие плоские сборки демонстрируют интригующее плавание по спиральным траекториям — с плоскостью тела, почти перпендикулярной направлению плавания, — которые привлекали внимание натуралистов с 18 века. [52] [53] [56] Тем не менее, способ, которым колонии Gonium смещают свое плавание к свету, остается неясным. Ранние микроскопические наблюдения выявили дифференциальную активность жгутиков между освещенной и затененной сторонами колонии как источник фототактической переориентации. [52] [53] Тем не менее, полное описание динамики жидкости, количественно связывающее реакцию жгутиков на изменения света и гидродинамические силы и моменты, действующие на колонию, все еще отсутствует. С эволюционной точки зрения фототаксис у Gonium поднимает фундаментальные вопросы, такие как степень, в которой фототактическая стратегия одноклеточного предка сохраняется в колониальной форме, как фототактическая реакция жгутиков адаптируется к геометрии и симметрии колонии и как это приводит к эффективной переориентации. [26]
Такси протистов: Направленное движение
Фототаксис
Распределение трехмерного фототаксиса в древе эукариот [57]
Красные стрелки указывают на вероятную точку происхождения фототаксиса в данной группе. Вопросительные знаки указывают на неопределенности относительно независимого или общего происхождения.
Некоторые простейшие могут двигаться к стимулу или от него, движение называется таксис . Например, движение к свету, называемое фототаксисом , осуществляется путем соединения их стратегии передвижения с органом восприятия света. [58] Эукариоты впервые в истории жизни развили способность следовать направлению света в трех измерениях в открытой воде. Стратегия эукариотической сенсорной интеграции, сенсорной обработки, а также скорость и механика тактических ответов принципиально отличаются от таковых у прокариот. [59] [57]
Как одноклеточные, так и многоклеточные эукариотические фототактические организмы имеют фиксированную форму, поляризованы, плавают по спирали и используют реснички для плавания и фототактического управления. Передача сигналов может происходить через прямые ионные токи , активируемые светом , аденилатциклазы или тримерные G-белки . Используемые фоторецепторы также могут быть очень разными (см. ниже). Однако передача сигналов во всех случаях в конечном итоге изменяет активность биения ресничек. [57] Механика фототактической ориентации аналогична у всех эукариот. Фотосенсор с ограниченным углом обзора вращается, сканируя пространство, и периодически подает сигналы ресничкам, чтобы изменить их биение, что изменит направление спиральной траектории плавания. Трехмерный фототаксис можно обнаружить в пяти из шести основных групп эукариот ( opisthokonts , Amoebozoa , plants , chromalveolates , excavates , rhizaria ). [57]
Пелагический фототаксис присутствует у зеленых водорослей , но не присутствует у глаукофитовых или красных водорослей . [57] У зеленых водорослей есть «стигма», расположенная в самой внешней части хлоропласта , непосредственно под двумя мембранами хлоропласта . Стигма состоит из десятков или нескольких сотен липидных глобул, которые часто образуют гексагональные массивы и могут быть расположены в один или несколько рядов. Липидные глобулы содержат сложную смесь каротиноидных пигментов, которые обеспечивают функцию экранирования и оранжево-красный цвет, [60] а также белков, которые стабилизируют глобулы. [61] Стигма расположена латерально, в фиксированной плоскости относительно ресничек, но не непосредственно рядом с базальными тельцами. [62] [63] Фиксированное положение обеспечивается прикреплением хлоропласта к одному из ресничных корней. [64] Пигментированное стигма не следует путать с фоторецептором. Стигма обеспечивает только направленное затенение для соседних фоторецепторов, вставленных в мембрану (термин «глазное пятно» поэтому вводит в заблуждение). Стигматы также могут отражать и фокусировать свет как вогнутое зеркало, тем самым повышая чувствительность. [57]
У наиболее изученной зеленой водоросли Chlamydomonas reinhardtii фототаксис опосредован пигментом родопсином , что впервые было продемонстрировано при восстановлении нормального фотоповедения у слепого мутанта аналогами хромофора сетчатки . [65] Два родопсина архебактериального типа, каналородопсин -1 и -2, [66] [67] были идентифицированы как рецепторы фототаксиса у Chlamydomonas . [68] Оба белка имеют N-концевую 7-трансмембранную часть, похожую на архебактериальные родопсины, за которой следует приблизительно 400 остатков С-концевой мембранно-ассоциированной части. CSRA и CSRB действуют как светозависимые катионные каналы и запускают деполяризующие фототоки. [68] [69] Было показано, что CSRA локализуется в области стигмы с помощью иммунофлуоресцентного анализа (Suzuki et al. 2003). Индивидуальное истощение РНК-интерференции как CSRA, так и CSRB изменило токи, вызванные светом, и показало, что CSRA опосредует быстрый, высоконасыщающий ток, тогда как CSRB — медленный, низконасыщающий. Оба тока способны вызывать фотофобные реакции и могут играть роль в фототаксисе, [70] [69], хотя точный вклад двух рецепторов пока не ясен. [57]
Как и у всех биконтов (растений, хромальвеолят, экскаватов, ризариев), у зеленых водорослей есть две реснички, которые не идентичны. Передняя ресничка всегда моложе задней . [71] [72] В каждом клеточном цикле одна дочерняя клетка получает переднюю ресничку и трансформирует ее в заднюю. Другая дочь наследует заднюю, зрелую ресничку. Затем обе дочери выращивают новую переднюю ресничку. [57]
Как и все другие ресничные пловцы, зеленые водоросли всегда плавают по спирали. Рукояльность спирали надежна и гарантируется хиральностью ресничек. Две реснички зеленых водорослей имеют разные паттерны биения и функции. У Chlamydomonas каскад фототрансдукции изменяет паттерн удара и скорость биения двух ресничек по-разному в сложной схеме. [44] [45] Это приводит к переориентации спиральной траектории плавания до тех пор, пока спиральная ось плавания не выровнена с вектором света. [57]
Термотаксис
Chlamydomonas , род одноклеточных зеленых водорослей с двумя жгутиками, каждый из которых равен другому, [73] проявляет как фототаксис, так и термотаксис.
Температура является ключевым фактором окружающей среды для живых организмов, поскольку скорости химических реакций и физические характеристики биологических материалов могут существенно меняться в зависимости от температуры. Живые организмы приспосабливаются к холоду и тепловому стрессу, используя приобретенные механизмы, включая способность мигрировать в среду с температурой, подходящей для обитания. Одной из простейших форм поведения для миграции в подходящую тепловую среду является термотаксис . Термотаксис был обнаружен у многоклеточных организмов, таких как Caenorhabditis elegans и Drosophila melanogaster , а также у одноклеточных организмов, таких как Paramecium caudatum , Dictyostelium discoideum , Physarum polycephalum и Escherichia coli . [74] Отдельные клетки внутри многоклеточных организмов также демонстрируют термотаксис. Например, сперма млекопитающих мигрирует через яйцевод к месту оплодотворения, руководствуясь повышением температуры. [75] [76]
Исследование того, как одноклеточные организмы мигрируют в направлении предпочитаемых температур, началось более 100 лет назад. [74] В частности, термотактическое поведение клеток Paramecium было хорошо изучено. Клетки Paramecium скапливаются в местах, близких к температуре культивирования, то есть к температуре, при которой выращиваются клетки. [74] Накопление в этих местах происходит потому, что клетки часто меняют направление своего движения, когда сталкиваются с изменением температуры, которое отклоняется от температуры культивирования, и увеличивают скорость своего движения, когда они испытывают изменение температуры, приближающееся к температуре культивирования. [77] [78] Изменение направления движения вызывается деполяризующим рецепторным потенциалом, который запускает потенциал действия в ресничках. [79] Эти исследования клеток Paramecium выявили термотаксис у одноклеточных организмов более 30 лет назад, но молекулярные механизмы терморецепции и передачи сигнала до сих пор не изучены. [76]
Понимание молекулярных механизмов термотаксиса значительно продвинулось в последние годы благодаря исследованиям спермы млекопитающих . Человеческая сперма мигрирует в сторону более высоких температур, от 29 °C до 41 °C. [75] Сперма может обнаруживать температурный градиент всего лишь 0,014 °C/мм, что позволяет предположить, что сперма обнаруживает временные изменения температуры, а не пространственные различия. [75] Было предложено несколько молекул в качестве сенсорных молекул, включая каналы опсина и транзиторного рецепторного потенциала (TRP), такие как TRPV1 , TRPV4 и TRPM8 . [80] [81] [82] Каналы TRP являются мультимодальными сенсорами для тепловых, химических и механических стимулов, но функция опсинов как термосенсора еще не установлена. [76]
Температура является критическим фактором окружающей среды также для клеток Chlamydomonas , которые вырабатывают небольшие белки теплового шока , шаперонины и белки теплового шока HSP70 , а также подвергаются другим реакциям теплового шока, чтобы справиться с тепловым стрессом. [83] [84] [85] [86] В ответ на холодовой шок 4 °C клетки останавливают пролиферацию и накапливают крахмал и сахар. [87] Ожидается, что поведенческие реакции, направленные на избегание стрессовых теплых или холодных условий, присутствуют у Chlamydomonas . Хотя сообщается, что клетки C. moewusii мигрируют в сторону более теплых температур в градиенте от 10 °C до 15 °C, [88] не было ни одного сообщения, в котором температурный диапазон систематически изменялся бы для изучения связи с температурой культивирования. Исследование 2019 года продемонстрировало термотаксис у Chlamydomonas reinhardtii и обнаружило, что при температуре от 10 °C до 30 °C клетки Chlamydomonas мигрируют в сторону более низких температур независимо от температуры культивирования. [76]
В отличие от термотаксиса, фототаксис был широко изучен у Chlamydomonas . Два жгутика Chlamydomonas бьют в стиле гребка брассом во время плавания вперед, и во время фототаксиса клетки Chlamydomonas поворачиваются к источнику света или от него, контролируя баланс движущих сил, создаваемых двумя жгутиками. [41] [89] Баланс зависит от концентрации ионов кальция внутри жгутика; таким образом, потеря кальций-зависимого контроля у мутантов ptx1 приводит к дефекту фототаксиса. [43] [90] [91] Направление фототаксиса у Chlamydomonas зависит от интенсивности света, но также зависит от внутриклеточных окислительно-восстановительных (редокс) условий. [92] Клетки мигрируют к источнику света, когда интенсивность света слабая, но направление меняется на противоположное в восстановительных условиях. Напротив, клетки уплывают от источников света с высокой интенсивностью, но направление меняется на противоположное в окислительных условиях. [76]
Скорость плавания
Скорость плавания жгутиконосцев и инфузорий [2] Линейное распределение данных о скорости плавания
Подогнанных логнормальных кривых для скоростей плавания жгутиконосцев и инфузорий
Скорости плавания были нормализованы путем деления их на медиану распределения. Нормализованные распределения показывают замечательное сходство, как в подобранных кривых, так и в неопределенности оценки. [2]
У жгутиконосных водорослей резкие изменения интенсивности света или интенсивные световые стимулы вызывают быстрое изменение положения жгутиков и временное обратное плавание. [94] [95] У зеленых водорослей это действие может быть опосредовано сократительным корневым волокном, которое изменяет угол между базальными тельцами . [96] Клетки также могут быстро реагировать на неожиданные механические стимулы. Сокращения по принципу «все или ничего» у стебельчатой инфузории Vorticella могут происходить со скоростью 8 см/с. [97] У некоторых видов солнечников аксоподы могут полностью втягиваться в течение 20 мс , чтобы втянуть пойманную добычу для фагоцитоза . [97] [98] [93]
Эти быстрые реакции обычно вызываются потенциалами действия — однонаправленными электрическими импульсами, включающими быстрые, регенеративные изменения мембранного потенциала . Хотя все клетки демонстрируют некоторую электрическую активность, филогенетические данные свидетельствуют о том, что способность распространять потенциалы действия могла быть наследственной эукариотической чертой, поддерживаемой последним общим эукариотическим предком. Они могли возникнуть в ответ на случайное повреждение мембраны и внезапный приток кальция. [99] Биоэлектрическая сигнализация в форме потенциалов действия происходит на порядки быстрее, чем любые другие сигнальные модальности, например, химическая диффузия , фосфорилирование белков и т. д. [93]
Для того чтобы инициировать быстрые реакции побега , они могли быть напрямую связаны с аппаратом подвижности, в частности, с гибкими, мембранно-непрерывными структурами, такими как реснички и псевдоподии . Потеря потенциалзависимых натриевых / кальциевых каналов также коррелирует с потерей ресничек во многих таксонах. У простейших потенциалы действия «все или ничего» возникают почти исключительно в связи с цилиарными мембранами, [100] [101] [102] за исключением некоторых нересничных диатомовых водорослей . [103] [104] Градуированные потенциалы возникают у амеб , также для контроля движения. [105] [93]
У Chlamydomonas токи жгутиков, подобные потенциалу действия, вызывают фотофобные реакции и инверсию жгутиков (через потенциалзависимый кальциевый канал Cav2), в то время как токи фоторецепторов вызывают гораздо более мягкие реакции. [106] Здесь механосенсорный канал семейства транзиторных рецепторных потенциалов локализован в основании ресничек, тогда как Cav2 локализован только в дистальных областях ресничек. [107] [108] У Paramecium гиперполяризации увеличивают частоту биения ресничек, в то время как деполяризации оказывают противоположный эффект и в конечном итоге приводят к инверсии ресничек. Деполяризации выше определенного порога приводят к потенциалам действия из-за открытия каналов Cav, расположенных исключительно в мембране ресничек. [109] [110] Калиевые каналы , также находящиеся в мембране, помогают восстановить мембранный потенциал покоя . [93]
Эукариоты манипулируют своим мембранным потенциалом для достижения переходов между различными видами поведения. Сложные биоэлектрические последовательности были зарегистрированы в связи с интегрированным поведением питания и хищничества у Favella . [111] Повторяющееся поведение возникает из-за ритмических спайков. У инфузорий ритмические деполяризации контролируют быструю и медленную ходьбу с помощью щупальцеобразных составных ресничек, называемых цирри , [112] позволяя выходить из тупиков [113] и ритуалов ухаживания в конъюгирующих гаметах . [114] [115] У Stentor потенциалы действия вызывают сокращения всего тела. [116] Наконец, возбудимые системы, работающие вблизи бифуркаций, могут допускать предельные циклы, которые проявляются как повторяющиеся или ритмичные электрические спайки и повторяющееся поведение. В конечном итоге это может привести к привыканию. [117] [118] [93]
Вверху: Схемы этапов производства биогибридных C. reinhardtii . Внизу: СЭМ-изображения голых микроводорослей (слева) и биогибридных микроводорослей (справа), покрытых наночастицами оксида железа, покрытыми хитозаном (CSIONP). Изображения были псевдоокрашены. Более темный зеленый цвет на правом СЭМ-изображении представляет собой хитозановое покрытие на клеточной стенке микроводорослей. Частицы оранжевого цвета представляют собой CSIONP.
Биогибридные микропловцы могут быть определены как микропловцы, которые состоят как из биологических, так и из искусственных компонентов, например, одного или нескольких живых микроорганизмов, прикрепленных к одной или различным синтетическим частям. [120] [121] В 1999 году Монтеманьо и Башан опубликовали статью, в которой были определены конкретные стратегии прикрепления биологических молекул к наноизготовленным субстратам, что позволило подготовить гибридные неорганические/органические наноэлектромеханические системы (НЭМС). [122] Они описали производство больших количеств F1-АТФазы из термофильных бактерий Bacillus PS3 для приготовления биомолекулярных моторов F1-АТФазы , иммобилизованных на наномассиве золота, меди или никеля, полученном с помощью электронно-лучевой литографии . Эти белки были прикреплены к микросферам размером в один микрон, помеченным синтетическим пептидом . Следовательно, они завершили подготовку платформы с химически активными участками и разработку биогибридных устройств, способных преобразовывать энергию биомолекулярных моторов в полезную работу. [121]
За последнее десятилетие биогибридные микророботы, в которых живые мобильные микроорганизмы физически интегрированы с непривязанными искусственными структурами, приобрели растущий интерес для обеспечения активного передвижения и доставки грузов к целевому месту назначения. [123] [124] [125] [126] В дополнение к подвижности, внутренние возможности восприятия и вызова соответствующей реакции на искусственные и экологические изменения делают биогибридных микророботов на основе клеток привлекательными для транспортировки грузов в недоступные полости человеческого тела для локальной активной доставки диагностических и терапевтических агентов. [127] [128] [129] Активное передвижение, нацеливание и управление концентрированными терапевтическими и диагностическими агентами, встроенными в мобильных микророботов, к месту действия могут преодолеть существующие проблемы традиционных методов лечения. [130] [131] [132] С этой целью бактерии обычно использовались с прикрепленными шариками и теневыми клеточными телами. [133] [134] [135] [136] [137] [138] [139] [140] [119]
Chlamydomonas reinhardtii — одноклеточная зеленая микроводоросль . Дикий тип C. reinhardtii имеет сферическую форму, средний диаметр которой составляет около 10 мкм. [141] Этот микроорганизм может воспринимать видимый свет и управляться им (т. е. фототаксисом ) с высокой скоростью плавания в диапазоне 100–200 мкм/с. [129] Он обладает естественной автофлуоресценцией , что позволяет проводить флуоресцентную визуализацию без меток. [141] C. reinhardtii активно изучалась в качестве живого компонента биогибридных микророботов для активной доставки терапевтических средств. [129] Они биосовместимы со здоровыми клетками млекопитающих, не оставляют известных токсинов, подвижны в физиологически релевантных средах и допускают модификацию поверхности для переноса груза на клеточной стенке. [129] [142] [143] [144] [145] Для сборки C. reinhardtii были предложеныальтернативные стратегии прикрепления посредством модификации взаимодействующих поверхностей посредством электростатических взаимодействий [129] [142] и ковалентной связи. [146] [119]
^ Харрис, Элизабет Х. (7 марта 2009 г.). Справочник по хламидомонаде: Введение в хламидомонаду и ее лабораторное использование: Том 1. Academic Press. ISBN97-80080919553.
^ Адам, Родни Д. (2001). «Биология Giardia lamblia». Clinical Microbiology Reviews . 14 (3): 447–475. doi :10.1128/CMR.14.3.447-475.2001. PMC 88984. PMID 11432808 .
^ Брей, Деннис (2 ноября 2000 г.). Движения клеток: от молекул к подвижности. Garland Science. ISBN9781136844355.
^ Капучинелли, П. (1980). «Движение эукариотических клеток». Подвижность живых клеток . С. 59–74. doi :10.1007/978-94-009-5812-8_4. ISBN978-0-412-15770-7.
^ Сани, MA (1974) Реснички и жгутики . Academic Press.
^ Никастро, Д.; Макинтош, Дж. Р.; Баумейстер, В. (2005). «Трехмерная структура эукариотических жгутиков в состоянии покоя, выявленная с помощью криоэлектронной томографии». Труды Национальной академии наук . 102 (44): 15889–15894. Bibcode : 2005PNAS..10215889N. doi : 10.1073 /pnas.0508274102 . PMC 1276108. PMID 16246999.
^ Моран, Джонатан; МакКин, Пол Г.; Джинджер, Майкл Л. (2014). «Эукариотические жгутики: изменения формы, функции и состава в ходе эволюции». BioScience . 64 (12): 1103–1114. doi : 10.1093/biosci/biu175 . ISSN 1525-3244.
^ Доусон, Скотт С.; Паредес, Александр Р. (2013). «Альтернативные цитоскелетные ландшафты: цитоскелетная новизна и эволюция у базальных экскавационных протистов». Current Opinion in Cell Biology . 25 (1): 134–141. doi :10.1016/j.ceb.2012.11.005. PMC 4927265. PMID 23312067 .
^ ab Atkinson, A.; Polimene, L.; Fileman, ES; Widdicombe, CE; McEvoy, AJ; Smyth, TJ; Djeghri, N.; Sailley, SF; Cornwell, LE (2018). ""Комментарий. Что движет сезонностью планктона в стратифицирующемся шельфовом море? Некоторые конкурирующие и взаимодополняющие теории"]" (PDF) . Лимнология и океанография . 63 (6): 2877–2884. Bibcode :2018LimOc..63.2877A. doi :10.1002/lno.11036. S2CID 91380765.
^ Синглтон, Пол (2006). Словарь микробиологии и молекулярной биологии, 3-е издание, пересмотренное . Чичестер, Великобритания: John Wiley & Sons. стр. 32. ISBN978-0-470-03545-0.
^ Дэвид Дж. Паттерсон. «Амебы: простейшие, которые двигаются и питаются с помощью псевдоподий». Веб-проект «Дерево жизни».
^ "Амебы". Эдинбургский университет. Архивировано из оригинала 10 июня 2009 года.
↑ Биологический словарь, 2004, дата обращения 01.01.2011.
^ Паттерсон, Дэвид Дж. (2000) «Жгутиконосцы: гетеротрофные протисты со жгутиками» Древо жизни .
^ ab Brennen, C.; Winet, H. (1977). "Механика жидкости и движения с помощью ресничек и жгутиков". Annual Review of Fluid Mechanics . 9 : 339–398. Bibcode :1977AnRFM...9..339B. doi :10.1146/annurev.fl.09.010177.002011.
^ Аллен, NS; Аллен, RD (1978). «Цитоплазматический поток в зеленых растениях». Annual Review of Biophysics and Bioengineering . 7 : 497–526. doi :10.1146/annurev.bb.07.060178.002433. PMID 352247.
^ Lyons, RA; Saridogan, E.; Djahanbakhch, O. (2006). «Репродуктивное значение ресничек фаллопиевых труб человека». Human Reproduction Update . 12 (4): 363–372. doi : 10.1093/humupd/dml012 . PMID 16565155.
^ Лауга, Эрик; Томас Р. Пауэрс (25 августа 2009 г.). «Гидродинамика плавающих микроорганизмов». Reports on Progress in Physics . 72 (9): 096601. arXiv : 0812.2887 . Bibcode : 2009RPPh...72i6601L. doi : 10.1088/0034-4885/72/9/096601. S2CID 3932471.
^ ab Роде, Себастьян; Элгети, Йенс; Гомпер, Герхард (2021). «Многоресничные микропловцы–метахрональная координация и спиральное плавание». The European Physical Journal E . 44 (6): 76. arXiv : 2103.11447 . doi :10.1140/epje/s10189-021-00078-x. PMC 8187229 . PMID 34101070.
^ Фриц-Лейлин, Лиллиан К. (2020). «Эволюция подвижности животных клеток». Current Biology . 30 (10): R477–R482. Bibcode : 2020CBio...30.R477F. doi : 10.1016/j.cub.2020.03.026 . PMID 32428485. S2CID 218711237.
^ abcdef Де Малепрад, Элен; Моиси, Фредерик; Исикава, Такудзи; Голдштейн, Рэймонд Э. (2020). «Подвижность и фототаксис гониума, простейшей дифференцированной колониальной водоросли». Physical Review E. 101 ( 2): 022416. arXiv : 1911.08837 . Bibcode : 2020PhRvE.101b2416D. doi : 10.1103/PhysRevE.101.022416. PMID 32168596. S2CID 211858528.Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ ab Coleman, AW (2012). "Сравнительный анализ Volvocaceae (Chlorophyta)1". Журнал Phycology . 48 (3): 491–513. Bibcode :2012JPcgy..48..491C. doi :10.1111/j.1529-8817.2012.01168.x. PMID 27011065. S2CID 422091.
^ Пеннак, Роберт В. (1978). Пресноводные беспозвоночные Соединенных Штатов (Второе издание). John Wiley & Sons . С. 43. ISBN0-471-04249-8.
↑ Вейсман, Август (1889). «Очерки о наследственности и родственных биологических проблемах: Доктор Август Вейсман... Ред. Эдвард Б. Поултон... Сельмар Шёнланд... И Артур Э. Шипли... Авторизованный перевод».
^ Хаксли, Джулиан С. (22 марта 2012 г.). Индивид в царстве животных. Cambridge University Press. ISBN9781107606074.
^ Кирк, Дэвид Л. (8 сентября 2005 г.). Volvox: Поиск молекулярных и генетических истоков многоклеточности и клеточной дифференциации. Cambridge University Press. ISBN9780521019149.
^ Голдштейн, Рэймонд Э. (2015). «Зеленые водоросли как модельные организмы для биологической гидродинамики». Annual Review of Fluid Mechanics . 47 : 343–375. doi :10.1146/annurev-fluid-010313-141426. PMC 4650200. PMID 26594068 .
^ Goldstein, Raymond E. (2016). «Лекция на соискание премии бакалавра по динамике жидкости в масштабе клетки». Журнал механики жидкости . 807 : 1–39. Bibcode : 2016JFM...807....1G. doi : 10.1017/jfm.2016.586 . S2CID 55745525.
^ abc Hoops, HJ (1997). «Подвижность колониальных и многоклеточных Volvocales: структура, функция и эволюция». Protoplasma . 199 (3–4): 99–112. doi :10.1007/BF01294499. S2CID 22315728.
^ ab Дрешер, К.; Голдштейн, Р. Э.; Тувал, И. (2010). «Верность адаптивного фототаксиса». Труды Национальной академии наук . 107 (25): 11171–11176. doi : 10.1073/pnas.1000901107 . PMC 2895142. PMID 20534560 .
^ Tsang, Alan CH; Lam, Amy T.; Riedel-Kruse, Ingmar H. (2018). «Полигональное движение и адаптивный фототаксис посредством переключения биения жгутиков у микроплавунца Euglena gracilis». Nature Physics . 14 (12): 1216–1222. Bibcode :2018NatPh..14.1216T. doi :10.1038/s41567-018-0277-7. S2CID 126294173.
^ ab Foster, KW; Smyth, RD (1980). «Световые антенны в фототактических водорослях». Microbiological Reviews . 44 (4): 572–630. doi :10.1128/mr.44.4.572-630.1980. PMC 373196. PMID 7010112 .
^ Хегеманн, Питер (2008). «Сенсорные фоторецепторы водорослей». Ежегодный обзор биологии растений . 59 : 167–189. doi :10.1146/annurev.arplant.59.032607.092847. PMID 18444900.
^ ab Камия, Р.; Витман, ГБ (1984). «Субмикромолярные уровни кальция контролируют баланс биения между двумя жгутиками в демембранированных моделях хламидомонады». Журнал клеточной биологии . 98 (1): 97–107. doi :10.1083/jcb.98.1.97. PMC 2112995. PMID 6707098 .
^ ab Josef, Keith; Saranak, Jureepan; Foster, Kenneth W. (2005). «Поведение ресничек отрицательно фототаксической Chlamydomonas reinhardtii». Подвижность клеток и цитоскелет . 61 (2): 97–111. doi :10.1002/cm.20069. PMID 15849714.
^ ab Josef, Keith; Saranak, Jureepan; Foster, Kenneth W. (2006). «Анализ линейных систем поведения рулевого управления ресничек, связанного с отрицательным фототаксисом у Chlamydomonas reinhardtii». Cell Motility and the Cytoskeleton . 63 (12): 758–777. doi :10.1002/cm.20158. PMID 16986140.
^ Йошимура, Кендзиро; Камия, Ритсу (2001). «Чувствительность фоторецептора хламидомонады оптимизирована для частоты вращения тела клетки». Физиология растений и клеток . 42 (6): 665–672. doi : 10.1093/pcp/pce084 . PMID 11427687.
^ Лептос, Кириакос К.; Чиоччоли, Маурицио; Фурлан, Сильвано; Пеши, Адриана И .; Голдштейн, Рэймонд Э. (2018). «Адаптивный жгутиковый фотоответ определяет динамику точного фототаксического управления у хламидомонады ». doi : 10.1101/254714. S2CID 90374721.{{cite journal}}: Цитировать журнал требует |journal=( помощь )
^ Кирк, Дэвид Л. (2004). "Volvox". Current Biology . 14 (15): R599–R600. Bibcode : 2004CBio...14.R599K. doi : 10.1016/j.cub.2004.07.034 . PMID 15296767. S2CID 235312006.
^ abc Мур, AR (1916). «Механизм ориентации у гониума». Журнал экспериментальной зоологии . 21 (3): 431–432. Bibcode :1916JEZ....21..431M. doi :10.1002/jez.1400210306.
^ abc Mast, SO (1916). «Процесс ориентации в колониальном организме Gonium pectorale и изучение структуры и функции глазного пятна». Журнал экспериментальной зоологии . 20 (1): 1–17. Bibcode : 1916JEZ....20....1M. doi : 10.1002/jez.1400200102.
^ ab Greuel, Brian T.; Floyd, Gary L. (1985). «Развитие жгутикового аппарата и жгутиковой ориентации у колониальной зеленой водоросли Gonium Pectorale (Volvocales)1». Journal of Phycology . 21 (3): 358–371. Bibcode : 1985JPcgy..21..358G. doi : 10.1111/j.0022-3646.1985.00358.x. S2CID 85760904.
^ Харпер, РА (1912). «Структура и развитие колонии гониума». Труды Американского микроскопического общества . 31 (2): 65–83. doi :10.2307/3221328. JSTOR 3221328.
^ Мюллер OF (1782) Kleine Schriften Aus Der Naturhistorie , Дессау, herausgegeben von JAE Goeze, стр. 15–21.
^ abcdefghij Jékely, Gáspár (2009). «Эволюция фототаксиса». Philosophical Transactions of the Royal Society B: Biological Sciences . 364 (1531): 2795–2808. doi :10.1098/rstb.2009.0072. PMC 2781859. PMID 19720645 .Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ Кларк, MA, Чой, J. и Дуглас, M. (2018) Характеристики биологии протистов 2e. OpenStax . ISBN 9781947172951 . Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ Хадер, Д.-П; Леберт, М. (19 июня 2001 г.). Фотодвижение. Эльзевир. ISBN9780080538860.
^ Грунг, Мерете; Креймер, Георг; Каленберг, Майкл; Мелконян, Майкл; Лиаен-Йенсен, Синнёве (1994). «Каротиноиды в аппарате глазного пятна жгутиконосной зеленой водоросли Spermatozopsis similis: адаптация к фоторецептору на основе сетчатки». Planta . 193 . doi :10.1007/BF00191604. S2CID 29443649.
^ Реннингер, С.; Бакендорф, Э.; Краймер, Г. (2001). «Субфракционирование аппаратов глазного пятна из зеленой водоросли Spermatozopsis similis: выделение и характеристика глобул глазного пятна». Planta . 213 (1): 51–63. Bibcode : 2001Plant.213...51R. doi : 10.1007/s004250000473. PMID 11523656. S2CID 24880210.
^ Арнотт, Ховард Дж.; Браун, Р. Малкольм (1967). «Ультраструктура глазного пятна и ее возможное значение в фототаксисе Tetracystis excentrica*†». Журнал протозоологии . 14 (4): 529–539. doi :10.1111/j.1550-7408.1967.tb02038.x.
^ Мелконян, М.; Робенек, Х. (1979). «Глазное пятно жгутиконосца Tetraselmis cordiformis stein (Chlorophyceae): структурная специализация наружной хлоропластной мембраны и ее возможное значение в фототаксисе зеленых водорослей». Protoplasma . 100 (2): 183–197. doi :10.1007/BF01283929. S2CID 24606055.
^ Мелконян, Майкл (1978). «Структура и значение крестообразных жгутиковых корневых систем у зеленых водорослей: сравнительные исследования видов Chlorosarcinopsis (Chlorosarcinales)». Систематика и эволюция растений . 130 (3–4): 265–292. Bibcode : 1978PSyEv.130..265M. doi : 10.1007/BF00982810. S2CID 22938771.
^ Нагель, Г.; Оллиг, Д.; Фурманн, М.; Катерия, С.; Мусти, AM; Бамберг, Э.; Хегеманн, П. (2002). «Канал родопсин-1: светозависимый протонный канал в зеленых водорослях». Наука . 296 (5577): 2395–2398. Бибкод : 2002Sci...296.2395N. дои : 10.1126/science.1072068. PMID 12089443. S2CID 206506942.
^ Nagel, G.; Szellas, T.; Huhn, W.; Kateriya, S.; Adeishvili, N.; Berthold, P.; Ollig, D.; Hegemann, P.; Bamberg, E. (2003). "Channelrhodopsin-2, a direct light-gated cation-selective membrane channel". Труды Национальной академии наук . 100 (24): 13940–13945. Bibcode : 2003PNAS..10013940N. doi : 10.1073/pnas.1936192100 . PMC 283525. PMID 14615590 .
^ ab Синещеков, О.А.; Юнг, К.-Х.; Спудич, Дж.Л. (2002). «Два родопсина опосредуют фототаксис к свету низкой и высокой интенсивности у Chlamydomonas reinhardtii». Труды Национальной академии наук . 99 (13): 8689–8694. doi : 10.1073/pnas.122243399 . PMC 124360. PMID 12060707 .
^ ab Бертольд, Питер; Цунода, Сатоши П.; Эрнст, Оливер П.; Магес, Вольфганг; Градманн, Дитрих; Хегеманн, Питер (2008). «Каналородопсин-1 инициирует фототаксис и фотофобные реакции у хламидомонады путем немедленной деполяризации, вызванной светом». The Plant Cell . 20 (6): 1665–1677. doi :10.1105/tpc.108.057919. PMC 2483371 . PMID 18552201.
^ Говорунова, Елена Г.; Юнг, Кванг-Хван; Синещеков, Олег А.; Спудич, Джон Л. (2004). «Сенсорные родопсины a и B хламидомонады: клеточное содержание и роль в фотофобных реакциях». Biophysical Journal . 86 (4): 2342–2349. Bibcode :2004BpJ....86.2342G. doi :10.1016/S0006-3495(04)74291-5. PMC 1304083 . PMID 15041672.
^ Кавальер-Смит, Т. (2002). «Фаготрофное происхождение эукариот и филогенетическая классификация простейших». Международный журнал систематической и эволюционной микробиологии . 52 (2): 297–354. doi :10.1099/00207713-52-2-297. PMID 11931142.
^ Кавальер-Смит, Томас (2009). «Мегафилогения, планы строения клеток, адаптивные зоны: причины и сроки базальной радиации эукариот». Журнал эукариотической микробиологии . 56 (1): 26–33. doi : 10.1111/j.1550-7408.2008.00373.x . PMID 19340985. S2CID 10205240.
^ Харрис, Элизабет Х. (2009) «Род Chlamydomonas» в The Chlamydomonas Sourcebook (Второе издание), глава 1, том 1, страницы 1-24. ISBN 9780080919553 doi :10.1016/B978-0-12-370873-1.00001-0
^ abc Дженнингс HS (1907) «Поведение низших организмов» The American Naturalist , 41 (481): 42-44.
^ abc Бахат, Анат; Тур-Каспа, Илан; Гакамски, Анна; Джохалас, Лаура К.; Брейтбарт, Хаим; Эйзенбах, Майкл (2003). «Термотаксис сперматозоидов млекопитающих: потенциальный механизм навигации в женских половых путях». Nature Medicine . 9 (2): 149–150. doi :10.1038/nm0203-149. hdl : 11336/66658 . PMID 12563318. S2CID 36538049.
^ abcde Секигути, Масая; Камеда, Сигетоши; Куросава, Сатоши; Ёсида, Мегуми; Ёсимура, Кенджиро (2018). «Термотаксис у Chlamydomonas вызывается возбуждением мембраны и контролируется окислительно-восстановительными условиями». Научные отчеты . 8 (1): 16114. Бибкод : 2018NatSR...816114S. дои : 10.1038/s41598-018-34487-4. ПМК 6208428 . ПМИД 30382191.Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ Тавада, К.; Осава, Ф. (1972). «Реакции парамеций на изменение температуры». Журнал протозоологии . 19 (1): 53–57. doi :10.1111/j.1550-7408.1972.tb03412.x. PMID 5008849.
^ Накаока, Ясуо; Осава, Фумио (1977). «Температурно-чувствительное поведение Paramecium caudatum». Журнал протозоологии . 24 (4): 575–580. doi :10.1111/j.1550-7408.1977.tb01018.x.
^ Хеннесси, Тодд М.; Саими, Йоширо; Кунг, Чинг (1983). «Деполяризация Paramecium , вызванная теплом , и ее связь с поведением избегания тепла». Журнал сравнительной физиологии A. 153 : 39–46. doi :10.1007/BF00610340. S2CID 7152549.
^ Клегг, Марк Р.; Маберли, Стивен К.; Джонс, Роджер И. (2003). «Поведенческие реакции пресноводных фитопланктонных жгутиконосцев на градиент температуры». Европейский журнал физиологии . 38 (3): 195–203. Bibcode : 2003EJPhy..38..195C. doi : 10.1080/0967026031000121697 . S2CID 85353895.
^ Исогай, Нахоко; Камия, Ритсу; Ёсимура, Кэндзиро (2000). «Доминирование между двумя жгутиками во время фототаксического поворота у хламидомонады». Зоологическая наука . 17 (9): 1261–1266. doi : 10.2108/zsj.17.1261 . S2CID 84890095.
^ Хорст, CJ; Витман, GB (1993). «Ptx1, нефототактический мутант Chlamydomonas, не контролирует доминирование жгутиков». Журнал клеточной биологии . 120 (3): 733–741. doi :10.1083/jcb.120.3.733. PMC 2119553. PMID 8425899 .
^ Окита, Норико; Исогай, Нахоко; Хироно, Масафуми; Камия, Рицу; Ёсимура, Кенджиро (2005). «Фототаксическая активность у нефототаксических мутантов Chlamydomonas с дефицитом Са2+-зависимого контроля над доминированием жгутиков или динеина во внутренней руке». Журнал клеточной науки . 118 (3): 529–537. дои : 10.1242/jcs.01633 . PMID 15657081. S2CID 2379702.
^ Вакабаяси, К.-и.; Мисава, И.; Мочиджи, С.; Камия, Р. (2011). «Окислительно-восстановительное равновесие регулирует знак фототаксиса у Chlamydomonas reinhardtii». Труды Национальной академии наук . 108 (27): 11280–11284. Bibcode : 2011PNAS..10811280W. doi : 10.1073/pnas.1100592108 . PMC 3131381. PMID 21690384 .
^ abcdef Ван, Кирсти Й .; Йекели, Гаспар (2021). «Истоки возбудимости эукариот». Philosophical Transactions of the Royal Society B: Biological Sciences . 376 (1820). arXiv : 2007.13388 . doi : 10.1098/rstb.2019.0758. PMC 7935092. PMID 33487111.Материал скопирован из этого источника, который доступен по лицензии Creative Commons Attribution 4.0 International.
^ Фостер, К. У.; Смит, Р. Д. (1980). «Световые антенны в фототактических водорослях». Microbiological Reviews . 44 (4): 572–630. doi :10.1128/MR.44.4.572-630.1980. PMC 373196. PMID 7010112 .
^ Холланд, Э.М.; Харц, Х.; Уль, Р.; Хегеманн, П. (1997). «Контроль фобических поведенческих реакций с помощью фототоков, вызванных родопсином, у хламидомонады». Biophysical Journal . 73 (3): 1395–1401. Bibcode :1997BpJ....73.1395H. doi :10.1016/S0006-3495(97)78171-2. PMC 1181038 . PMID 9284306.
^ Хаяси, Масахито; Яги, Тошики; Йошимура, Кэндзиро; Камия, Ритсу (1998). «Наблюдение в реальном времени за переориентацией базального тельца, вызванной Ca2+, у хламидомонады ». Подвижность клеток и цитоскелет . 41 (1): 49–56. doi :10.1002/(SICI)1097-0169(1998)41:1<49::AID-CM4>3.0.CO;2-A. PMID 9744298.
^ Андо, Мотонори; Сигенака, Ёсинобу (1989). «Структура и функция цитоскелета у солнечников: I. Механизм быстрого аксоподиального сокращения у эхинофагов ». Подвижность клеток и цитоскелет . 14 (2): 288–301. doi :10.1002/cm.970140214.
^ Брюне, Тибо; Арендт, Детлев (2016). «От ответа на повреждение к потенциалам действия: ранняя эволюция нейронных и сократительных модулей у стволовых эукариот». Philosophical Transactions of the Royal Society B: Biological Sciences . 371 (1685). doi :10.1098/rstb.2015.0043. PMC 4685582 . PMID 26598726.
^ Экерт, Р.; Брем, П. (1979). «Ионные механизмы возбуждения парамеций». Annual Review of Biophysics and Bioengineering . 8 : 353–383. doi :10.1146/annurev.bb.08.060179.002033. PMID 383005.
^ Экерт, Роджер; Найто, Ютака (1972). «Биоэлектрический контроль локомоции у инфузорий*†». Журнал протозоологии . 19 (2): 237–243. doi :10.1111/j.1550-7408.1972.tb03444.x. PMID 4624297.
^ Вуд, Дэвид К. (1982). «Проницаемость мембран, определяющая покой, действие и потенциалы механорецепторов у Stentor coeruleus». Журнал сравнительной физиологии A. 146 ( 4): 537–550. doi :10.1007/bf00609450. S2CID 21083419.
^ Тейлор, Элисон Р. (2009). "Быстрый потенциал действия на основе Na+/Ca2+ в морской диатомовой водоросли". PLOS ONE . 4 (3): e4966. Bibcode : 2009PLoSO...4.4966T. doi : 10.1371/journal.pone.0004966 . PMC 2654917. PMID 19305505 .
^ Хелливелл, Кэтрин Э.; Крахри, Абдул; Кестер, Джули А.; Варам, Сьюзан; Верре, Фредерик; Тейлор, Элисон Р.; Уилер, Глен Л.; Браунли, Колин (2019). «Альтернативные механизмы быстрой передачи сигналов Na+/Ca2+ у эукариот через новый класс однодоменных потенциал-зависимых каналов». Current Biology . 29 (9): 1503–1511.e6. doi :10.1016/j.cub.2019.03.041. PMC 6509283 . PMID 31006567.
^ Бингли, М.С.; Томпсон, К.М. (1962). «Биоэлектрические потенциалы в связи с движением амеб». Журнал теоретической биологии . 2 (1): 16–32. Bibcode : 1962JThBi...2...16B. doi : 10.1016/s0022-5193(62)80024-1.
^ Кимбалл РФ (1942) «Природа и наследование типов спаривания у Euplotes Patella». Генетика , 27 (3): 269–285. PMID 17247040, PMC PMC1209158.
^ Wood, DC (1988). «Привыкание при стенторе: вызванное модификацией механорецепторного канала». Журнал нейронауки . 8 (7): 2254–2258. doi : 10.1523 /JNEUROSCI.08-07-02254.1988. PMC 6569508. PMID 3249223.
^ Дженнингс, Х. С. (1899). «Исследования реакций на стимулы у одноклеточных организмов. III Реакции на локализованные стимулы у спиростом и стенторов». The American Naturalist . 33 (389): 373–389. doi :10.1086/277256. S2CID 85272784.
^ Карлсен, Рика Райт; Ситти, Метин (2014). «Биогибридные клеточные актуаторы для микросистем». Small . 10 (19): 3831–3851. doi :10.1002/smll.201400384. PMID 24895215.
^ Нгуен, Ван Ду; Хан, Джи-Вон; Чой, Ён Джин; Чо, Сунгхун; Чжэн, Шаохуэй; Ко, Сонг Ён; Пак, Чон-О; Пак, Сухо (2016). «Активный липосомальный бактериобот для лечения опухолей, объединяющий инкапсулированную в лекарство (паклитаксел) липосому с таргетингом бактерий (Salmonella Typhimurium)». Датчики и приводы B: Химические . 224 : 217–224. doi :10.1016/j.snb.2015.09.034.
^ ab Harris, Elizabeth H. (2001). "Chlamydomonasas Amodelorganism". Annual Review of Plant Physiology and Plant Molecular Biology . 52 : 363–406. doi :10.1146/annurev.arplant.52.1.363. PMID 11337403.
^ ab Weibel, DB; Garstecki, P.; Ryan, D.; Diluzio, WR; Mayer, M.; Seto, JE; Whitesides, GM (2005). «Микровоки: микроорганизмы для перемещения микромасштабных грузов». Труды Национальной академии наук . 102 (34): 11963–11967. Bibcode : 2005PNAS..10211963W. doi : 10.1073/pnas.0505481102 . PMC 1189341. PMID 16103369 .
Кон, Стэнли; Манойлов, Калина; Гордон, Ричард (2021). Скользящая подвижность диатомовых водорослей: биология и применение . Беверли, Массачусетс: Scrivener Publishing. ISBN 978-1-119-52648-3. OCLC 1262966612.







.jpg/1280px-Chlamydomonas_(10000x).jpg)





