
Растительные клетки — это клетки зеленых растений , фотосинтезирующих эукариот царства Plantae . Их отличительные особенности включают первичные клеточные стенки, содержащие целлюлозу, гемицеллюлозу и пектин, наличие пластид, способных осуществлять фотосинтез и запасать крахмал, крупную вакуоль , регулирующую тургорное давление, отсутствие жгутиков или центриолей , за исключением гамет, и уникальный метод клеточного деления, включающий образование клеточной пластинки или фрагмопласта , разделяющего новые дочерние клетки.
Растительные клетки дифференцируются из недифференцированных меристематических клеток (аналог стволовых клеток животных) с образованием основных классов клеток и тканей корней , стеблей , листьев , цветков и репродуктивных структур, каждый из которых может состоять из нескольких типов клеток.
Клетки паренхимы — это живые клетки, функции которых варьируются от хранения и поддержки до фотосинтеза ( клетки мезофилла ) и загрузки флоэмы ( переносные клетки ). Помимо ксилемы и флоэмы в проводящих пучках листья состоят преимущественно из клеток паренхимы. Некоторые клетки паренхимы, например эпидермиса, специализируются на проникновении и фокусировке света или регуляции газообмена , но другие относятся к наименее специализированным клеткам растительной ткани и могут оставаться тотипотентными , способными делиться с образованием новых популяций недифференцированных клеток. на протяжении всей их жизни. [17] Клетки паренхимы имеют тонкие, проницаемые первичные стенки, позволяющие транспортировать небольшие молекулы между ними, а их цитоплазма отвечает за широкий спектр биохимических функций, таких как секреция нектара или производство вторичных продуктов , препятствующих травоядности . Клетки паренхимы, содержащие множество хлоропластов и участвующие в основном в фотосинтезе, называются клетками хлоренхимы . Клетки хлоренхимы – это клетки паренхимы, участвующие в фотосинтезе. [18] Другие, такие как большинство клеток паренхимы клубней картофеля и семядоли семян бобовых , выполняют функцию хранения.
Клетки колленхимы живы в зрелом возрасте и имеют утолщенные клеточные стенки из целлюлозы. [19] Эти клетки созревают из производных меристемы, которые изначально напоминают паренхиму, но различия быстро становятся очевидными. Пластиды не развиваются, а секреторный аппарат (ЭР и Гольджи) пролиферирует, секретируя дополнительную первичную стенку. Стенка чаще всего бывает самой толстой в углах, где соприкасаются три или более ячеек, и самой тонкой, когда соприкасаются только две ячейки, хотя возможны и другие варианты утолщения стенки. [19] Пектин и гемицеллюлоза являются доминирующими компонентами клеточных стенок колленхимы двудольных покрытосеменных растений , которые могут содержать всего лишь 20% целлюлозы в Petasites . [20] Клетки колленхимы обычно довольно удлинены и могут делиться поперечно, образуя перегородки. Роль этого типа клеток заключается в поддержке растущих в длину осей растения, а также в придании тканям гибкости и прочности на растяжение. В первичной стенке отсутствует лигнин, который сделал бы ее жесткой и жесткой, поэтому этот тип клеток обеспечивает то, что можно было бы назвать пластической опорой – опорой, которая может удерживать молодой стебель или черешок на воздухе, но в клетках, которые могут растягиваться, как и клетки вокруг них. удлиненный. Растягивающаяся опора (без эластичной обратной фиксации) — хороший способ описать, что делает колленхима. Часть нитей сельдерея представляет собой колленхиму.
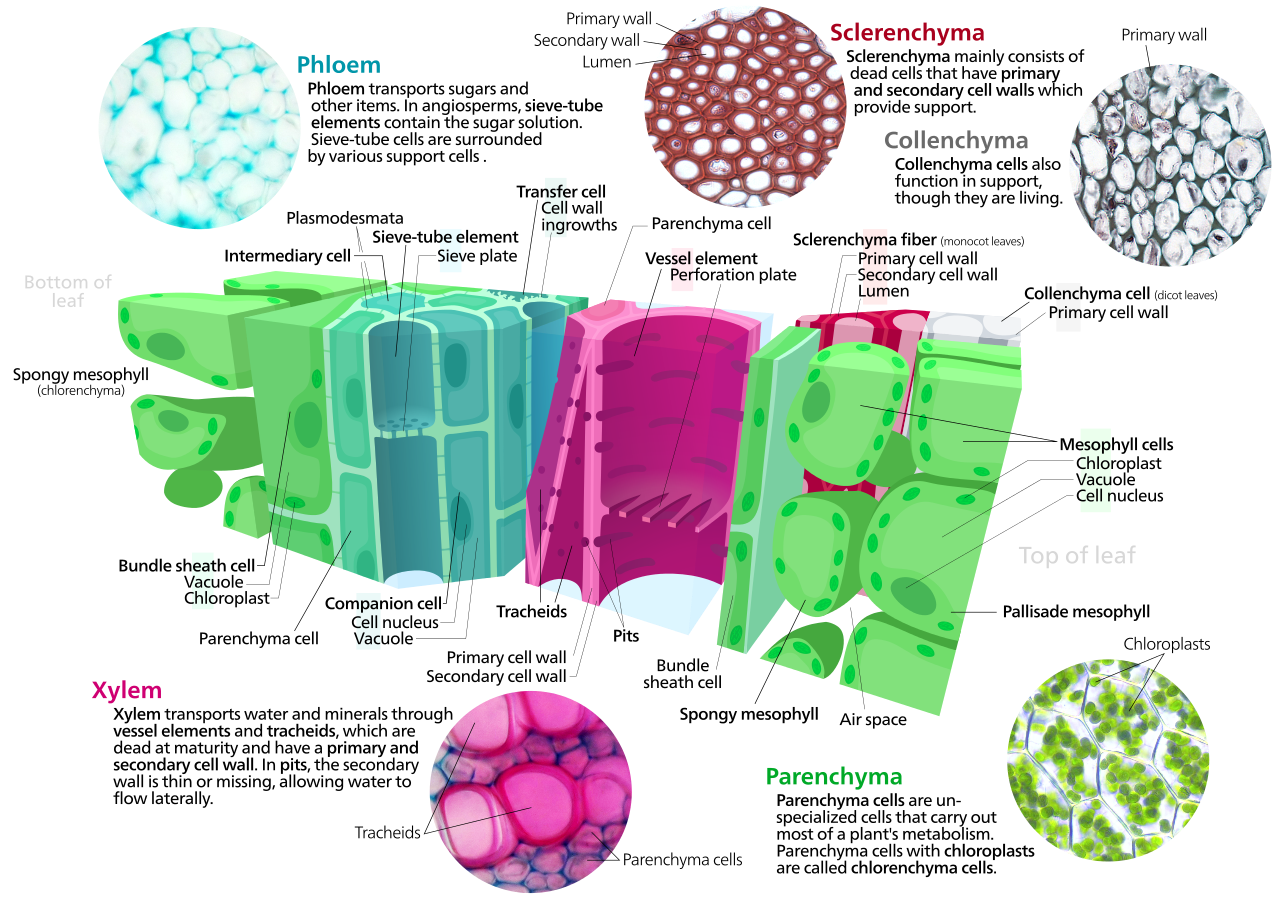
Склеренхима представляет собой ткань, состоящую из двух типов клеток: склереидов и волокон , имеющих утолщенные, лигнифицированные вторичные стенки [19] : 78, отложенные внутри первичной клеточной стенки . Вторичные стенки укрепляют клетки и делают их непроницаемыми для воды. Следовательно, склереиды и волокна при функциональной зрелости обычно погибают, а цитоплазма отсутствует, оставляя пустую центральную полость. Склереиды или каменные клетки (от греческого skleros — твердый ) — это твердые, прочные клетки, которые придают листьям или плодам шероховатую текстуру. Они могут препятствовать травоядности, повреждая пищеварительные пути на личиночных стадиях мелких насекомых. Склереиды образуют твердую стенку косточки персиков и многих других фруктов, обеспечивая физическую защиту развивающегося ядра. Волокна представляют собой удлиненные клетки с одревесневшими вторичными стенками, которые обеспечивают несущую поддержку и прочность на растяжение листьям и стеблям травянистых растений. Волокна склеренхимы не участвуют в проводимости ни воды и питательных веществ (как в ксилеме ), ни углеродных соединений (как во флоэме ), но вполне вероятно, что они развились как модификации инициалей ксилемы и флоэмы у ранних наземных растений.

Ксилема представляет собой сложную сосудистую ткань, состоящую из проводящих воду трахеид или сосудистых элементов вместе с волокнами и клетками паренхимы. Трахеиды [21] — удлиненные клетки с одревесневшими вторичными утолщениями клеточных стенок, специализированные для проведения воды, впервые появившиеся у растений при их переходе на сушу в силурийский период более 425 млн лет назад (см. Куксония ). Наличие ксилемных трахеид определяет сосудистые растения или трахеофиты. Трахеиды — заостренные, удлиненные клетки ксилемы, простейшие из которых имеют сплошные первичные клеточные стенки и одревесневшие утолщения вторичных стенок в виде колец, обручей или сетчатых сетей. Голосеменные характеризуются более сложными трахеидами с клапанообразными перфорациями, называемыми окаймленными ямками . У папоротников , других птеридофитов и голосеменных имеются только ксилемные трахеиды , а у цветковых растений — еще и ксилемные сосуды . Элементы сосудов представляют собой полые клетки ксилемы без торцевых стенок, выровненные встык так, что образуют длинные непрерывные трубки. У мохообразных отсутствует настоящая ксилемная ткань, но у их спорофитов имеется водопроводящая ткань, известная как гидром, состоящая из удлиненных клеток более простой конструкции.
Флоэма — это специализированная ткань для транспорта пищи у высших растений, которая в основном транспортирует сахарозу по градиентам давления, создаваемым осмосом (процесс, называемый транслокацией) . Флоэма представляет собой сложную ткань, состоящую из двух основных типов клеток: ситовидных трубок и тесно связанных с ними клеток-спутников , а также клеток паренхимы, флоэмных волокон и склереидов. [19] : 171 Ситовые трубки соединены встык с помощью перфорированных концевых пластин между так называемыми ситовыми пластинами , которые позволяют транспортировать фотосинтат между ситовыми элементами. Элементы ситовидной трубки лишены ядер и рибосом , а их метаболизм и функции регулируются соседними ядросодержащими клетками-компаньонами. Клетки-спутники, соединенные с ситовидными трубками через плазмодесмы , отвечают за загрузку флоэмы сахарами . У мохообразных отсутствует флоэма, но спорофиты мхов имеют более простую ткань с аналогичной функцией, известную как лептом.

Эпидермис растения представляет собой специализированную ткань, состоящую из клеток паренхимы, покрывающую внешнюю поверхность листьев, стеблей и корней. В эпидермисе могут присутствовать несколько типов клеток. Среди них следует выделить замыкающие клетки устьиц, которые контролируют скорость газообмена между растением и атмосферой, железистые и одежные волоски или трихомы , а также корневые волоски первичных корней. В эпидермисе побегов большинства растений хлоропласты имеются только в замыкающих клетках . Хлоропласты содержат зеленый пигмент хлорофилл, необходимый для фотосинтеза. Эпидермальные клетки надземных органов возникают из поверхностного слоя клеток, известного как оболочка (слои L1 и L2), который покрывает верхушку побега растения [19] , тогда как кора и сосудистые ткани возникают из самого внутреннего слоя верхушки побега, известного как корпус (слой L3). Эпидермис корней происходит из слоя клеток непосредственно под корневым чехликом. Эпидермис всех надземных органов, кроме корней, покрыт кутикулой из полиэфирного кутина или полимера кутана (или того и другого) с поверхностным слоем эпикутикулярных восков . Считается, что эпидермальные клетки первичного побега — единственные растительные клетки, обладающие биохимической способностью синтезировать кутин. [22]
{{cite book}}: CS1 maint: отсутствует местоположение издателя ( ссылка )