Где 3-оксо-5α-стероид и акцептор являются субстратами , а соответствующий 3-оксо-Δ4-стероид и восстановленный акцептор являются продуктами . Примером этой обобщенной реакции, которую катализирует 5α-редуктаза типа 2, является:
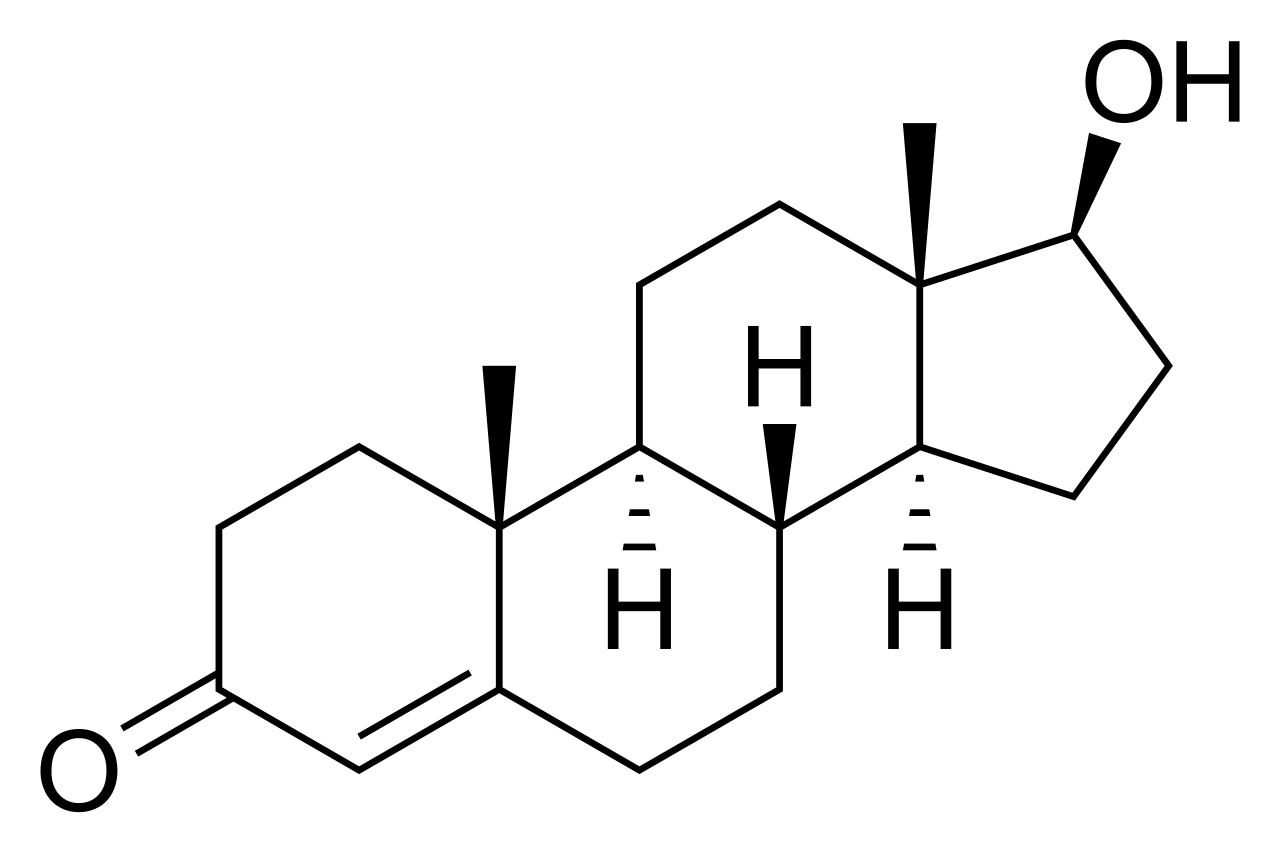
где дигидротестостерон представляет собой 3-оксо-5α-стероид, НАДФ + является акцептором, а тестостерон представляет собой 3-оксо-Δ4-стероид, а НАДФН – восстановленный акцептор.
5α-редуктазы действуют на стероиды 3-оксо (3-кето), Δ 4,5 C19/C21 в качестве своих субстратов; «3-кето» относится к двойной связи третьего углерода с кислородом. Углероды 4 и 5 также имеют двойную связь, обозначенную «Δ 4,5 ». Реакция включает стереоспецифический и постоянный разрыв Δ 4,5 с помощью НАДФН в качестве кофактора. Гидрид-анион (H-) также расположен на α-грани пятого углерода, а протон — на β-грани у углерода 4. [8]
Распределение по возрасту
5α-R1 экспрессируется в коже головы плода и внегенитальной коже спины в 5–50 раз меньше, чем у взрослых. 5α-R2 экспрессируется в предстательной железе плода так же, как и у взрослых. 5α-R1 экспрессируется главным образом в эпителии , а 5α-R2 — в строме предстательной железы плода. Ученые искали экспрессию 5α-R2 в печени, надпочечниках, яичках, яичниках, головном мозге, волосистой части головы, груди и коже половых органов плода с помощью иммуноблоттинга и смогли обнаружить его только в коже половых органов. [8]
После рождения 5α-R1 экспрессируется в большем количестве мест, включая печень, кожу, кожу головы и простату. 5α-R2 экспрессируется в предстательной железе, семенных пузырьках, придатках яичка, печени и, в меньшей степени, в волосистой части головы и коже. Экспрессия в печени как 5α-R1, так и 2 происходит немедленно, но исчезает в коже и волосистой части головы на 18-м месяце. Затем, в период полового созревания, только 5α-R2 реэкспрессируется в коже и волосистой части головы.
5α-R1 и 5α-R2, по-видимому, экспрессируются в предстательной железе у плодов мужского пола и на протяжении всей постнатальной жизни. 5α-R1 и 5α-R2 также экспрессируются, хотя и в разной степени, в печени, генитальной и внегенитальной коже, предстательной железе, придатках яичек, семенных пузырьках, семенниках, яичниках, матке, почках, экзокринной части поджелудочной железы и головном мозге. [3] [8]
Во взрослом возрасте 5α-R1-3 экспрессируется повсеместно .
Субстраты
Конкретные субстраты включают тестостерон , прогестерон , андростендион , [9] эпитестостерон , кортизол , альдостерон и дезоксикортикостерон . За исключением дигидротестостерона, большая часть физиологической роли 5α-восстановленных стероидов неизвестна. [8] Помимо восстановления тестостерона до дигидротестостерона, изоформы фермента 5-альфа-редуктазы I и II восстанавливают прогестерон до дигидропрогестерона (DHP), а дезоксикортикостерон до дигидродезоксикортикостерона (DHDOC). Модели in vitro и на животных предполагают, что последующее 3-альфа-восстановление DHT, DHP и DHDOC приводит к образованию стероидных метаболитов, влияние на функцию мозга достигается за счет усиления эргического ингибирования ГАМК . Эти производные нейроактивных стероидов усиливают действие ГАМК посредством аллостерической модуляции рецепторов ГАМК(А) и оказывают противосудорожное, антидепрессивное и анксиолитическое действие, а также изменяют сексуальное и связанное с алкоголем поведение. [10] 5α-дигидрокортизол присутствует во внутриглазной жидкости глаза, синтезируется в хрусталике и может способствовать образованию самой водянистой влаги. [11] Аллопрегнанолон и THDOC являются нейростероидами , причем последний оказывает влияние на восприимчивость животных к судорогам. У социально изолированных мышей уровень 5α-R1 специфически снижается в глутаматергических пирамидных нейронах , которые сходятся к миндалевидному телу из областей коры и гиппокампа. Это подавление может быть причиной появления поведенческих расстройств, таких как тревога, агрессия и когнитивная дисфункция. [3] [4] 5α-дигидроальдостерон является мощным антинатриуретиком , хотя и отличается от альдостерона . Его образование в почках усиливается при ограничении потребления соли, что позволяет предположить, что он может способствовать удержанию натрия следующим образом: [12]
Это механизм 5α-редуктазы тестостерона, превращающий его в ДГТ (дигидротестостерон). 5α-редуктаза работает, используя восстанавливающую способность НАДФН для осуществления гидридного сдвига двойной углеродной связи в кольце, вызывая образование енолята и последующую таутамеризацию с образованием ДГТ. [13]
5α-DHP является основным гормоном в кровообращении у нормальных велосипедистов и беременных женщин. [14]
Вид сбоку 5AR с моноолеином и НАДФН в гидрофобном кармане; вид на 7 трансмембранных доменов сверху
5α-редуктаза представляет собой мембраносвязанный фермент, который катализирует НАДФН- зависимое восстановление двойных связей в стероидных субстратах для повышения эффективности. [15] Кристаллическая структура гомолога изоферментов 5α-редуктазы 1 и 2 обнаружена у протеобактерий (протеобактерии 5α-редуктаза). Он существует в виде мономера с трансмембранной структурой из семи альфа-спиралей, содержащей гидрофобный карман, который удерживает кофактор НАДФН и моноолеин, который занимает карман связывания стероидного субстрата. [16] В клетках насекомых моноолеин не обнаружен, но заменяется другими андрогенами и ингибиторами. [17] Интегральная трансмембранная топология семи, вероятно, консервативна у разных видов: N-конец находится в просвете эндоплазматического ретикулума , а C-конец обращен к цитозолю . Высокая конформационная динамика цитозольной области, вероятно, регулирует обмен НАДФН/НАДФ+. [17] Сохранение последовательностей в известных кристаллических структурах подтверждает высокую консервативность структуры фермента. [16]
Ингибирование фермента можно разделить на две категории: стероидное, которое является необратимым, и нестероидное. Есть и другие стероидные ингибиторы, например, финастерид (МК-906), дутастерид (GG745), 4-МА, туростерид, МК-386, МК-434 и МК-963. Исследователи занялись синтезом нестероидных препаратов для ингибирования 5α-редуктазы из-за нежелательных побочных эффектов стероидов. Наиболее мощные и селективные ингибиторы 5α-R1 обнаружены в этом классе и включают бензохинолоны, нестероидные арильные кислоты, производные бутановой кислоты и, что более известно, полиненасыщенные жирные кислоты (особенно линоленовая кислота ), цинк и зеленый чай . [8] Рибофлавин также был идентифицирован как ингибитор 5α-редуктазы. [18]
Гинекомастия , эректильная дисфункция , нарушение когнитивной функции, утомляемость , гипогликемия , нарушение функции печени, запор и депрессия — это лишь некоторые из возможных побочных эффектов ингибирования 5α-редуктазы. Сообщалось о долгосрочных побочных эффектах, которые продолжались даже после отмены препарата. [20]
Финастерид
Финастерид ингибирует два изофермента 5α-редуктазы (II и III), тогда как дутастерид ингибирует все три. [2] Финастерид мощно ингибирует 5α-R2 при средней ингибирующей концентрации IC 50 69 нМ, но менее эффективен с 5α-R1 до IC 50 360 нМ. [21] Финастерид снижает средний уровень ДГТ в сыворотке на 71% через 6 месяцев, [22] и было показано, что in vitro ингибирует 5α-R3 с такой же эффективностью, как и 5α-R2 в трансфицированных клеточных линиях. [2]
Дутастерид
Дутастерид ингибирует изоферменты 5α-редуктазы 1 и 2 типа лучше, чем финастерид, что приводит к более полному снижению уровня ДГТ через 24 недели (94,7% против 70,8%). [23] Он также снижает внутрипростатический ДГТ на 97% у мужчин с раком простаты при приеме 5 мг/день в течение трех месяцев. [24] Второе исследование с применением 3,5 мг/день в течение 4 месяцев снизило внутрипростатический ДГТ еще на 99%. [25] Подавление ДГТ in vivo и сообщение о том, что дутастерид ингибирует 5α-R3 in vitro [26] позволяют предположить, что дутастерид может быть тройным ингибитором 5α-редуктазы. [8]
Это механизм ингибирования финастерида на 5α-редуктазе [27]. Финастерид ингибирует 5α-редуктазу посредством конкурентного ингибирования [28], поэтому этот механизм показывает, что НАДФ+ подвергается атаке со стороны образующегося енолята, а не происходит мгновенная таутамеризация, как при случай действия 5α-редуктазы на тестостерон. Финастерид в конечном итоге высвобождается из фермент-субстратного комплекса 5α-редуктазы, поэтому его не считают ингибитором самоубийства, поскольку в конечном итоге образующимся продуктом является дигидрофинастерид.
Врожденные недостатки
5α-редуктаза 1
У самцов мышей, инактивированных 5α-редуктазой 1 типа, снижается костная масса и сила хвата мышц передних конечностей, что, как предполагается, связано с отсутствием экспрессии 5α-редуктазы 1 типа в костях и мышцах. [29] Считается, что у мужчин с дефицитом 5-альфа-редуктазы 2-го типа изофермент 1-го типа ответственен за вирилизацию в период полового созревания. [6]
5α-редуктаза 2
Нарушение активности 5α-редуктазы 2 может быть результатом мутаций в основном гене SRD5A2. Состояние, известное как дефицит 5α-редуктазы 2 , имеет ряд проявлений в виде атипичного внешнего вида наружных половых органов у мужчин. Это связано с тем, что 5α-редуктаза 2 катализирует трансформацию тестостерона в мощный андроген дигидротестостерон, который необходим для правильной маскулинизации мужских гениталий. [30]
5α-редуктаза 3
Когда малая интерферирующая РНК используется для подавления экспрессии изозима 5α-R3 в клеточных линиях , наблюдается снижение роста клеток, их жизнеспособности и снижение соотношения DHT/T. [31] Также была продемонстрирована способность аденовирусных векторов снижать уровень тестостерона, андростендиона и прогестерона в андроген-стимулированных клеточных линиях простаты. [8]
Врожденный дефицит 5α-R3 в гене SRD53A связан с редким аутосомно-рецессивным заболеванием, при котором пациенты рождаются с тяжелой интеллектуальной дисфункцией и дефектами мозжечка и глаз. Предполагаемый дефицит заключается в уменьшении концевой связи полипренола с долихолом, что является важным этапом N-гликозилирования белков, что, в свою очередь, важно для правильного сворачивания остатков аспарагина на образующемся белке в эндоплазматическом ретикулуме . [32]
Нервная система
Аффективные расстройства
Было показано, что изолированное выращивание снижает экспрессию белка изоферментов 5α-редуктазы 1 и 2 в корковых и подкорковых областях мозга на моделях крыс. Однако количество 5α-восстановленного метаболита осталось неизменным. Это означает, что изолированное выращивание, вероятно, приводит к изменениям в экспрессии и активности 5α-редуктазы в головном мозге, что приводит к нарушению регуляции нейротрансмиссии дофамина , что приводит к раннему хроническому стрессу [33]. Было показано, что лечение финастеридом , ингибитором 5α-редуктазы, оказывает имитируют эффекты СИОЗС, вызывающие сексуальную дисфункцию . [34] Исследования показали, что 5α-редуктаза является ферментом, ограничивающим скорость синтеза нейростероидов , в частности, превращения прогестерона в аллопрегнанолон , [35] низкие уровни аллопрегнанолона связаны с депрессией , тревогой и шизофренией . Лишение сна может усилить экспрессию и активность 5α-редуктазы в префронтальной коре головного мозга , что приводит к симптомам, связанным с манией, у крыс. [35] Также оспаривается, связано ли использование ингибиторов 5α-редуктазы с суицидальными мыслями и депрессией у пациентов, которые используют их при доброкачественной гиперплазии предстательной железы . [36] [37] Эти симптомы были обнаружены во время активного применения ингибиторов и при непосредственном наблюдении. [36] Однако неизвестно, возникают ли эти симптомы естественным путем из-за доброкачественной гиперплазии предстательной железы. [37]
Дисфункция гипоталамо-гипофизарно-надпочечниковой системы.
Альтернативный механизм регуляции кортизола регулируется с помощью 5α-редуктазы, которая катализирует восстановление А-кольца кортизола, метаболизируя это соединение. [38] Типы 1 и 2 5α-редуктазы являются основными ферментами, участвующими в клиренсе кортизола через печень . [39] Избыток кортизола связан с неалкогольной жировой болезнью печени (НАЖБП), но исследования in vitro показали, что чрезмерная экспрессия 5α-редуктазы типа 2 может подавлять липогенез . [40] Ключевая роль 5α-редуктазы в расщеплении кортизола и накоплении жира прояснила некоторые побочные эффекты ингибиторов 5α-редуктазы. В рандомизированных исследованиях на людях-добровольцах было обнаружено, что ингибирование 5α-редуктазы с помощью дутастерида и финастерида может привести к накоплению липидов в печени у мужчин. [41] При критических заболеваниях чрезмерная стимуляция кортизола как часть реакции на стресс может привести к снижению клиренса кортизола через печень через 5α-редуктазу и почки через 11β-гидроксистероиддегидрогеназу типа 2. [ 39] длительное повышение уровня кортизола может привести к Синдром Кушинга .
Номенклатура
Этот фермент принадлежит к семейству оксидоредуктаз , а именно, действующих на группу донора CH-CH с другими акцепторами. Систематическое название этого класса ферментов — 3-оксо-5α-стероид:акцептор Δ4 - оксидоредуктаза . Другие широко используемые имена включают:
^ Пинна Дж., Агис-Бальбоа Р.К., Пибири Ф., Нельсон М., Гуидотти А., Коста Э. (октябрь 2008 г.). «Биосинтез нейростероидов регулирует половой диморфный страх и агрессивное поведение у мышей». Нейрохимические исследования . 33 (10): 1990–2007. дои : 10.1007/s11064-008-9718-5. PMID 18473173. S2CID 19338424.
^ abcd Ямана К., Лабри Ф., Луу-В. (август 2010 г.). «Человеческая 5α-редуктаза 3-го типа экспрессируется в периферических тканях на более высоких уровнях, чем 1-й и 2-й типы, и ее активность сильно ингибируется финастеридом и дутастеридом». Гормональная молекулярная биология и клинические исследования . 2 (3): 293–9. дои : 10.1515/hmbci.2010.035. PMID 25961201. S2CID 28841145.
^ abc Агис-Бальбоа RC, Пинна Г, Жуби А, Малоку Э, Вельдик М, Коста Э и др. (сентябрь 2006 г.). «Характеристика нейронов головного мозга, которые экспрессируют ферменты, опосредующие биосинтез нейростероидов». Труды Национальной академии наук Соединенных Штатов Америки . 103 (39): 14602–7. Бибкод : 2006PNAS..10314602A. дои : 10.1073/pnas.0606544103 . ПМК 1600006 . ПМИД 16984997.
^ ab Агис-Бальбоа RC, Пинна Г, Пибири Ф, Кадриу Б, Коста Э, Гуидотти А (ноябрь 2007 г.). «Понижающая регуляция биосинтеза нейростероидов в кортиколимбических цепях опосредует поведение у мышей, вызванное социальной изоляцией». Труды Национальной академии наук Соединенных Штатов Америки . 104 (47): 18736–41. Бибкод : 2007PNAS..10418736A. дои : 10.1073/pnas.0709419104 . ПМК 2141846 . ПМИД 18003893.
^ Киллиан Дж., Пратис К., Клифтон Р.Дж., Стэнтон П.Г., Робертсон Д.М., О'Доннелл Л. (май 2003 г.). «Изоферменты 5-альфа-редуктазы 1 и 2 в семенниках крыс во время постнатального развития». Биология размножения . 68 (5): 1711–8. doi : 10.1095/biolreprod.102.009142 . ПМИД 12606426.
^ аб Тиле С., Хоппе У., Холтерхус П.М., Хиорт О. (июнь 2005 г.). «Изофермент 5-альфа-редуктазы 1-го типа в большом количестве транскрибируется в нормальных фибробластах кожи гениталий человека и может играть важную роль в маскулинизации мужчин с дефицитом 5-альфа-редуктазы 2-го типа». Европейский журнал эндокринологии . 152 (6): 875–80. дои : 10.1530/eje.1.01927 . ПМИД 15941927.
^ Годой А, Кавински Е, Ли Ю, Ока Д, Алексиев Б, Аззуни Ф и др. (июль 2011 г.). «Экспрессия 5α-редуктазы типа 3 в доброкачественных и злокачественных тканях человека: сравнительный анализ при прогрессировании рака простаты». Простата . 71 (10): 1033–46. дои :10.1002/pros.21318. ПМЦ 4295561 . ПМИД 21557268.
^ abcdefg Аззуни Ф, Годой А, Ли Ю, Молер Дж (2012). «Семейство изоферментов 5-альфа-редуктазы: обзор фундаментальной биологии и их роли в заболеваниях человека». Достижения в урологии . 2012 : 530121. doi : 10.1155/2012/530121 . ПМЦ 3253436 . ПМИД 22235201.
^ аб Паба С., Фрау Р., Годар С.К., Девото П., Марросу Ф., Бортолато М. (2011). «Стероидная 5α-редуктаза как новая терапевтическая мишень при шизофрении и других нервно-психических расстройствах». Текущий фармацевтический дизайн . 17 (2): 151–67. doi : 10.2174/138161211795049589 (неактивен 11 апреля 2024 г.). ПМИД 21361868.{{cite journal}}: CS1 maint: DOI неактивен по состоянию на апрель 2024 г. ( ссылка )
^ Финн Д.А., Бидлс-Болинг А.С., Бекли Э.Х., Форд М.М., Гилилланд К.Р., Горин-Мейер Р.Э. и др. (2006). «Новый взгляд на ингибитор 5-альфа-редуктазы финастерид». Обзоры препаратов для ЦНС . 12 (1): 53–76. дои : 10.1111/j.1527-3458.2006.00053.x. ПМК 6741762 . ПМИД 16834758.
^ Вайнштейн Б.И., Кандалафт Н., Ритч Р., Камрас CB, Моррис DJ, Латиф С.А. и др. (июнь 1991 г.). «5-альфа-дигидрокортизол в водянистой жидкости человека и метаболизм кортизола человеческими линзами in vitro». Исследовательская офтальмология и визуальные науки . 32 (7): 2130–5. ПМИД 2055703.
^ Ахмед С., Денисон С. (сентябрь 1998 г.). «Механизм представления активного сайта 5-альфа-редуктазы (5AR)». Письма по биоорганической и медицинской химии . 8 (18): 2615–70. дои : 10.1016/S0960-894X(98)00463-6. ПМИД 9873591.
^ Милевич Л., Гомес-Санчес С., Кроули Г., Портер Дж.К., Мэдден Дж.Д., Макдональд ПК (октябрь 1977 г.). «Прогестерон и 5альфа-прегнан-3,20-дион в периферической крови здоровых молодых женщин: ежедневные измерения на протяжении менструального цикла». Журнал клинической эндокринологии и метаболизма . 45 (4): 617–22. doi : 10.1210/jcem-45-4-617 . ПМИД 914969.
^ Уилсон JD (8 февраля 2002 г.). «Роль 5-альфа-редукции в физиологии стероидных гормонов». Воспроизводство, рождаемость и развитие . 13 (7–8): 673–8. дои : 10.1071/rd01074. PMID 11999320. Архивировано из оригинала 26 мая 2021 года . Проверено 6 марта 2021 г.
^ Аб Хань Ю, Чжуан Q, Сунь Б, Лев В, Ван С, Сяо Q и др. (январь 2021 г.). «Кристаллическая структура стероидредуктазы SRD5A обнаруживает консервативный механизм восстановления стероидов». Природные коммуникации . 12 (1): 449. Бибкод : 2021NatCo..12..449H. дои : 10.1038/s41467-020-20675-2. ПМЦ 7815742 . ПМИД 33469028.
^ Аб Сяо Q, Ван Л, Супекар С, Шен Т, Лю Х, Йе Ф и др. (октябрь 2020 г.). «Структура человеческой стероидной 5α-редуктазы 2 с антиандрогенным препаратом финастеридом». Природные коммуникации . 11 (1): 5430. Бибкод : 2020NatCo..11.5430X. doi : 10.1038/s41467-020-19249-z. ПМЦ 7591894 . ПМИД 33110062.
^ Накаяма О, Яги М, Киёто С, Окухара М, Косака М (декабрь 1990 г.). «Рибофлавин, ингибитор тестостерон-5-альфа-редуктазы». Журнал антибиотиков . 43 (12): 1615–6. дои : 10.7164/антибиотики.43.1615 . ПМИД 2276981.
^ Андерссон С (2001). «Стероидогенные ферменты кожи». Европейский журнал дерматологии . 11 (4): 293–5. ПМИД 11399532.
^ Ирвиг М.С., Колукула С. (июнь 2011 г.). «Постоянные сексуальные побочные эффекты финастерида при выпадении волос по мужскому типу». Журнал сексуальной медицины . 8 (6): 1747–53. дои : 10.1111/j.1743-6109.2011.02255.x. ПМИД 21418145.
^ Тиан Г., Стюарт Дж.Д., Мосс М.Л., Доманико П.Л., Брэмсон Х.Н., Патель И.Р. и др. (март 1994 г.). «17-бета-(N-трет-бутилкарбамоил)-4-аза-5 альфа-андростан-1-ен-3-он представляет собой активный сайт-направленный медленный, зависящий от времени ингибитор человеческой стероидной 5-альфа-редуктазы 1». Биохимия . 33 (8): 2291–6. дои : 10.1021/bi00174a041. ПМИД 8117686.
^ МакКоннелл Дж.Д., Уилсон Дж.Д., Джордж Ф.В., Геллер Дж., Паппас Ф., Стоунер Э. (март 1992 г.). «Финастерид, ингибитор 5-альфа-редуктазы, подавляет дигидротестостерон простаты у мужчин с доброкачественной гиперплазией предстательной железы». Журнал клинической эндокринологии и метаболизма . 74 (3): 505–8. doi : 10.1210/jcem.74.3.1371291. ПМИД 1371291.
^ Кларк Р.В., Герман DJ, Каннингем Г.Р., Уилсон Т.Х., Моррилл Б.Б., Хоббс С. (май 2004 г.). «Заметное подавление дигидротестостерона у мужчин с доброкачественной гиперплазией предстательной железы дутастеридом, двойным ингибитором 5-альфа-редуктазы». Журнал клинической эндокринологии и метаболизма . 89 (5): 2179–84. дои : 10.1210/jc.2003-030330 . ПМИД 15126539.
^ Андриол Г.Л., Хамфри П., Рэй П., Глив М.Э., Трахтенберг Дж., Томас Л.Н. и др. (сентябрь 2004 г.). «Влияние двойного ингибитора 5-альфа-редуктазы дутастерида на маркеры регрессии опухоли при раке простаты». Журнал урологии . 172 (3): 915–9. дои : 10.1097/01.ju.0000136430.37245.b9. ПМИД 15310997.
^ Глив М., Цянь Дж., Андреу С., Поммервилль П., Чин Дж., Кейси Р. и др. (ноябрь 2006 г.). «Влияние двойного ингибитора 5-альфа-редуктазы дутастерида на локализованный рак простаты - результаты 4-месячного исследования перед радикальной простатэктомией». Простата . 66 (15): 1674–85. дои : 10.1002/pros.20499 . PMID 16927304. S2CID 40446842.
^ Мосс GP (декабрь 1989 г.). «Объединенная комиссия IUPAC-IUB по биохимической номенклатуре (JCBN). Номенклатура стероидов. Рекомендации 1989 г.». Европейский журнал биохимии . 186 (3): 429–58. doi :10.1111/j.1432-1033.1989.tb15228.x. ПМИД 2606099.
^ Друри Дж. Э., Ди Костанцо Л., Пеннинг Т. М., Кристиансон Д. В. (июль 2009 г.). «Ингибирование стероидной 5-бета-редуктазы человека (AKR1D1) финастеридом и структура комплекса фермент-ингибитор». Журнал биологической химии . 284 (30): 19786–90. дои : 10.1074/jbc.c109.016931 . ПМК 2740403 . ПМИД 19515843.
^ Хулин-Кёртис С.Л., Пети Д., Фигг В.Д., Хсинг А.В., Райхардт Дж.К. (декабрь 2010 г.). «Метаболизм финастерида и фармакогенетика: новые подходы к персонализированной профилактике рака простаты». Будущая онкология . 6 (12): 1897–913. дои : 10.2217/фон.10.149. ПМК 6300128 . ПМИД 21142863.
^ Виндал С.Х., Андерссон Н., Бёрьессон А.Е., Суонсон С., Свенссон Дж., Мовераре-Скртик С. и др. (2011). Ванакер Дж. М. (ред.). «Снижение костной массы и мышечной силы у самцов мышей, инактивированных 5α-редуктазой 1 типа». ПЛОС ОДИН . 6 (6): e21402. Бибкод : 2011PLoSO...621402W. дои : 10.1371/journal.pone.0021402 . ПМК 3120862 . ПМИД 21731732.
^ Зиннекер Г.Х., Хиорт О., Диббелт Л., Альберс Н., Дорр Х.Г., Хаус Х. и др. (3 мая 1996 г.). «Фенотипическая классификация мужского псевдогермафродитизма вследствие дефицита стероидной 5α-редуктазы 2». Американский журнал медицинской генетики . 63 (1): 223–230. doi :10.1002/(SICI)1096-8628(19960503)63:1<223::AID-AJMG39>3.0.CO;2-O. ПМИД 8723114.
^ Уэмура М., Тамура К., Чунг С., Хонма С., Окуяма А., Накамура Ю. и др. (январь 2008 г.). «Новая 5-альфа-стероидредуктаза (SRD5A3, тип-3) сверхэкспрессируется при гормонорезистентном раке простаты». Раковая наука . 99 (1): 81–6. дои : 10.1111/j.1349-7006.2007.00656.x . PMID 17986282. S2CID 51733620.
^ Кантагрел В., Лефебер Д.Д., Нг Б.Г., Гуан З., Силхави Дж.Л., Биелас С.Л. и др. (июль 2010 г.). «SRD5A3 необходим для превращения полипренола в долихол и мутирует при врожденном нарушении гликозилирования». Клетка . 142 (2): 203–17. doi :10.1016/j.cell.2010.06.001. ПМК 2940322 . ПМИД 20637498.
^ Бортолато М., Девото П., Ронкада П., Фрау Р., Флор Г., Саба П. и др. (июнь 2011 г.). «Вызванное изоляцией снижение экспрессии 5α-редуктазы в мозге: значение для дофаминергических нарушений». Нейрофармакология . 60 (7–8): 1301–8. doi :10.1016/j.neuropharm.2011.01.013. PMID 21256141. S2CID 20164197.
^ Джатти С., Дивиккаро С., Панзика Дж., Мелканги RC (август 2018 г.). «Постфинастеридный синдром и сексуальная дисфункция после приема СИОЗС: две стороны одной медали?». Эндокринный . 61 (2): 180–193. дои : 10.1007/s12020-018-1593-5. PMID 29675596. S2CID 4974636.
^ ab Фрау Р., Бини В., Соджиу А., Шегги С., Парду А., Фанни С. и др. (октябрь 2017 г.). «Нейростероидогенный фермент 5α-редуктаза опосредует психотические осложнения лишения сна». Нейропсихофармакология . 42 (11): 2196–2205. дои :10.1038/нпп.2017.13. ПМК 5603808 . ПМИД 28102229.
^ аб Велк Б., Макартур Э., Ордон М., Андерсон К.К., Хейворд Дж., Диксон С. (май 2017 г.). «Связь суицидальности и депрессии с ингибиторами 5α-редуктазы». JAMA Внутренняя медицина . 177 (5): 683–691. doi : 10.1001/jamainternmed.2017.0089. ПМК 5818776 . ПМИД 28319231.
^ аб Дайсон Т.Е., Кантрелл Массачусетс, Лунд, Британская Колумбия (октябрь 2020 г.). «Отсутствие связи между ингибиторами 5α-редуктазы и депрессией». Журнал урологии . 204 (4): 793–798. дои : 10.1097/JU.0000000000001079. PMID 32294395. S2CID 215794669.
^ Петреску А.Д., Кейн Дж., Лиере В., Хевенер Т., ДеМорроу С. (2018). «Дисфункция гипоталамуса-гипофиза-надпочечников при холестатическом заболевании печени». Границы эндокринологии . 9 : 660. дои : 10.3389/fendo.2018.00660 . ПМК 6240761 . ПМИД 30483216.
^ аб Боонен Э., Вервенн Х., Меерсеман П., Эндрю Р., Мортье Л., Деклерк П.Е. и др. (Апрель 2013). «Снижение метаболизма кортизола во время критического заболевания». Медицинский журнал Новой Англии . 368 (16): 1477–88. doi : 10.1056/NEJMoa1214969. ПМЦ 4413428 . ПМИД 23506003.
^ Насири М., Николау Н., Параджес С., Кроне Н.П., Валсамакис Г., Масторакос Г. и др. (август 2015 г.). «5α-редуктаза типа 2 регулирует действие глюкокортикоидов и метаболический фенотип в гепатоцитах человека». Эндокринология . 156 (8): 2863–71. дои : 10.1210/en.2015-1149. ПМК 4511138 . ПМИД 25974403.
^ Хэзлхерст Дж.М., Опреску А.И., Николау Н., Ди Гуида Р., Гринбергс А.Е., Дэвис Н.П. и др. (январь 2016 г.). «Ингибирование двойного 5α-редуктазы способствует накоплению липидов в печени у человека». Журнал клинической эндокринологии и метаболизма . 101 (1): 103–13. дои : 10.1210/jc.2015-2928. ПМК 4701851 . ПМИД 26574953.
дальнейшее чтение
Леви Х.Р., Талалай П. (август 1959 г.). «Бактериальное окисление стероидов. II. Исследования ферментативного механизма дегидрирования кольца А». Журнал биологической химии . 234 (8): 2014–21. дои : 10.1016/S0021-9258(18)69859-X . ПМИД 13673006.







{kind=link}