Most abundant type of granulocytes and the most abundant white blood cell
Нейтрофилы (также известные как нейтроциты , гетерофилы или полиморфно-ядерные лейкоциты ) представляют собой тип лейкоцитов . Точнее, они образуют наиболее распространенный тип гранулоцитов и составляют от 40% до 70% всех лейкоцитов у человека. [1] Они составляют важную часть врожденной иммунной системы , причем их функции различаются у разных животных. [2]
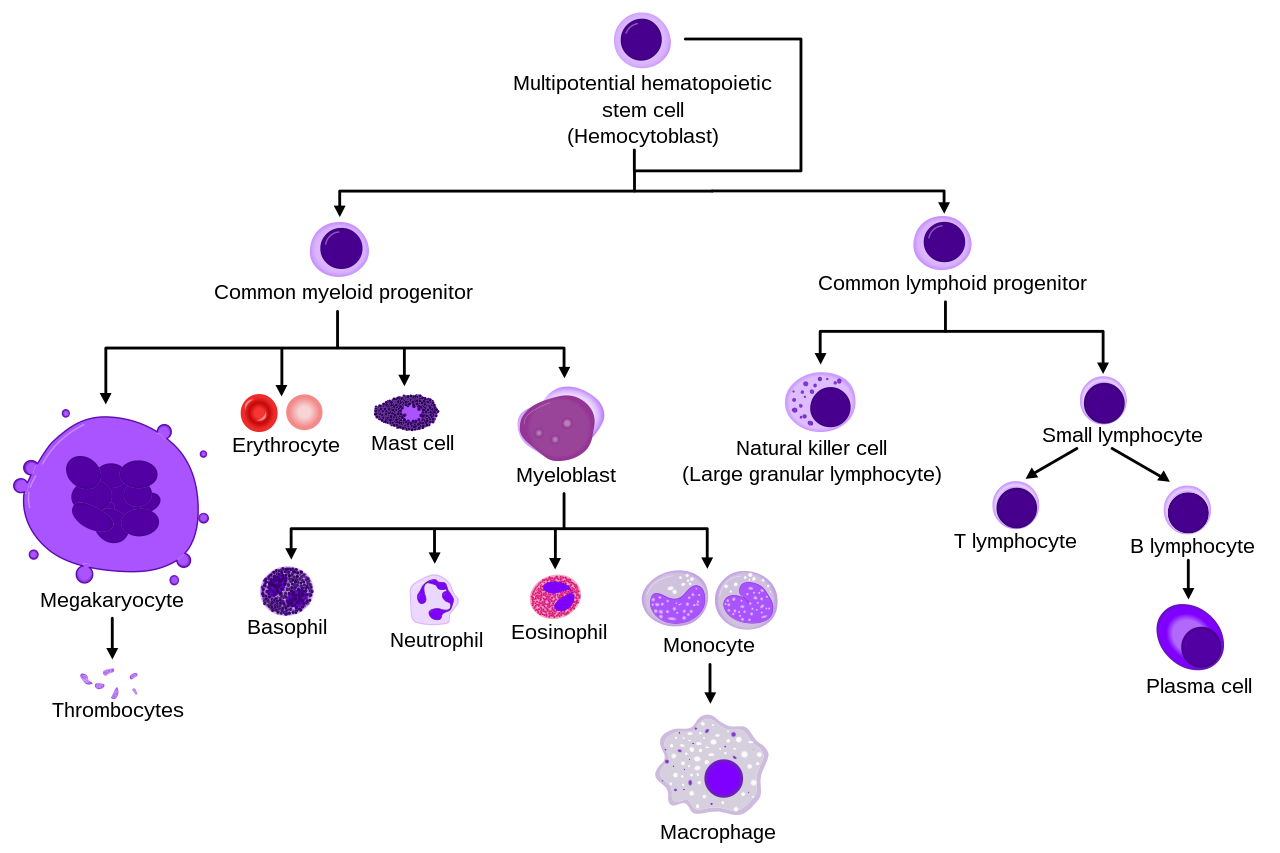
Они образуются из стволовых клеток костного мозга и дифференцируются в субпопуляции нейтрофилов-киллеров и нейтрофилов-клеток. Они недолговечны (от 5 до 135 часов, см. § Продолжительность жизни) и очень подвижны, поскольку могут проникать в те части тканей, куда другие клетки/молекулы не могут проникнуть. Нейтрофилы можно подразделить на сегментоядерные нейтрофилы и полосчатые нейтрофилы (или полосы ). Они составляют часть семейства полиморфно-ядерных клеток (ПМЯ) вместе с базофилами и эозинофилами . [3] [4] [5]
Название нейтрофил происходит от характеристик окрашивания гистологических или цитологических препаратов гематоксилина и эозина ( H&E ) . В то время как базофильные лейкоциты окрашиваются в темно-синий цвет, а эозинофильные лейкоциты окрашиваются в ярко-красный цвет, нейтрофилы окрашиваются в нейтральный розовый цвет. В норме нейтрофилы содержат ядро, разделенное на 2–5 долей. [6]
Нейтрофилы представляют собой разновидность фагоцитов и обычно обнаруживаются в кровотоке . Во время начальной ( острой ) фазы воспаления , особенно в результате бактериальной инфекции , воздействия окружающей среды [7] и некоторых видов рака, [8] [9] нейтрофилы являются одними из первых, кто реагирует на миграцию воспалительных клеток к месту воспаления. воспаление. Они мигрируют через кровеносные сосуды, а затем через интерстициальное пространство, следуя химическим сигналам, таким как интерлейкин-8 (IL-8), C5a , fMLP , лейкотриен B4 и перекись водорода (H 2 O 2 ) [10] в процессе, называемом хемотаксисом. . Они являются преобладающими клетками гноя , что объясняет его беловатый/желтоватый вид. [11]
Нейтрофилы рекрутируются в месте повреждения в течение нескольких минут после травмы и являются признаком острого воспаления; [12] однако из-за того, что некоторые патогены неперевариваются, они могут быть не в состоянии справиться с некоторыми инфекциями без помощи других типов иммунных клеток.
Состав
Нейтрофильный гранулоцит мигрирует из кровеносного сосуда в матрикс, секретируя протеолитические ферменты, растворяющие межклеточные связи (с улучшением его подвижности) и обволакивающие бактерии посредством фагоцитоза.Гиперсегментированный нейтрофил
Вместе с эозинофилами и базофилами они образуют класс полиморфно-ядерных клеток , названный в честь многодольчатой формы ядра (по сравнению с лимфоцитами и моноцитами , другими типами лейкоцитов). Ядро имеет характерный дольчатый вид: отдельные доли соединены хроматином . Ядрышко исчезает по мере созревания нейтрофила, что происходит лишь с некоторыми другими типами ядросодержащих клеток. [14] : 168 До 17% ядер женских нейтрофилов человека имеют придаток в форме голени, который содержит инактивированную Х-хромосому . [15] В цитоплазме аппарат Гольджи небольшой, митохондрии и рибосомы редки, шероховатая эндоплазматическая сеть отсутствует. [14] : 170 В цитоплазме также содержится около 200 гранул, из них треть азурофильных . [14] : 170
По мере взросления нейтрофилы будут демонстрировать увеличивающуюся сегментацию (множество сегментов ядра). Нормальный нейтрофил должен иметь 3–5 сегментов. Гиперсегментация не является нормой, но встречается при некоторых заболеваниях, особенно при дефиците витамина B12 . Это отмечается при ручном анализе мазка крови и является положительным, когда большинство или все нейтрофилы имеют 5 или более сегментов.
Нейтрофилы являются наиболее распространенными лейкоцитами у человека (приблизительно 10 11 производятся ежедневно); на их долю приходится примерно 50–70% всех лейкоцитов (лейкоцитов). Установленный нормальный диапазон показателей крови человека варьируется в зависимости от лаборатории, но число нейтрофилов 2,5–7,5 × 10 9 /л является стандартным нормальным диапазоном. Люди африканского и ближневосточного происхождения могут иметь более низкие показатели, что по-прежнему является нормальным. [16] В отчете нейтрофилы могут быть разделены на сегментоядерные нейтрофилы и полосы .
При циркуляции в кровотоке и инактивации нейтрофилы имеют сферическую форму. После активации они меняют форму, становятся более аморфными или похожими на амебы и могут расширять ложноножки , охотясь за антигенами . [17]
В 1973 году Санчес и др. обнаружили, что способность нейтрофилов поглощать бактерии снижается при приеме простых сахаров, таких как глюкоза, фруктоза, а также сахароза, мед и апельсиновый сок, тогда как прием крахмалов не оказывает никакого эффекта. С другой стороны, голодание усилило фагоцитарную способность нейтрофилов поглощать бактерии. Был сделан вывод, что при попадании в организм сахаров изменяется функция, а не количество фагоцитов в поглощающих бактериях. [18] В 2007 году исследователи из Института биомедицинских исследований Уайтхеда обнаружили, что при выборе сахаров на микробных поверхностях нейтрофилы реагировали преимущественно на некоторые типы сахаров. Нейтрофилы преимущественно поглощали и убивали мишени бета-1,6-глюкана по сравнению с мишенями бета-1,3-глюкана. [19] [20]
Согласно различным оценкам, средняя продолжительность жизни инактивированных нейтрофилов человека в кровообращении составляет от 5 до 135 часов (5 дней и 15 часов). [21] [22]
После активации они маргинализируются (прилегают к эндотелию кровеносных сосудов) и подвергаются селектин -зависимому захвату с последующей в большинстве случаев интегрин -зависимой адгезией, после чего мигрируют в ткани, где выживают в течение 1–2 дней. [23] Также было продемонстрировано, что нейтрофилы высвобождаются в кровь из резерва селезенки после инфаркта миокарда . [24]
Нейтрофилы гораздо более многочисленны, чем долгоживущие моноциты / макрофаги -фагоциты. Патоген (болезнетворный микроорганизм или вирус), скорее всего, сначала встретится с нейтрофилом . Некоторые эксперты предполагают, что короткое время жизни нейтрофилов является эволюционной адаптацией. Короткое время жизни нейтрофилов сводит к минимуму распространение тех патогенов, которые паразитируют на фагоцитах (например, Leishmania [25] ), поскольку чем больше времени такие паразиты проводят вне клетки -хозяина , тем больше вероятность того, что они будут уничтожены каким-либо компонентом защиты организма. Кроме того, поскольку противомикробные препараты нейтрофилов также могут повреждать ткани хозяина , их короткий срок действия ограничивает повреждение хозяина во время воспаления . [23]
Нейтрофилы подвергаются процессу, называемому хемотаксисом, посредством амебоидного движения , что позволяет им мигрировать к местам инфекции или воспаления. Рецепторы клеточной поверхности позволяют нейтрофилам обнаруживать химические градиенты молекул, таких как интерлейкин-8 (IL-8), интерферон гамма (IFN-γ), C3a, C5a и лейкотриен B4 , которые эти клетки используют для направления пути своей миграции.
Нейтрофилы имеют множество специфических рецепторов, в том числе рецепторы комплемента , цитокинов, таких как интерлейкины и IFN-γ, хемокинов , лектинов и других белков. Они также экспрессируют рецепторы для обнаружения и прикрепления к эндотелию и Fc - рецепторам опсонина . [26]
В лейкоцитах, реагирующих на хемоаттрактант , клеточная полярность регулируется активностью малых Rho- гуанозинтрифосфатаз ( Rho GTPases ) и фосфоинозитид-3-киназ ( PI3Ks ). В нейтрофилах липидные продукты PI3K регулируют активацию Rac1, гемопоэтических Rac2 и RhoG GTPases семейства Rho и необходимы для подвижности клеток . Rac-GTPases регулируют динамику цитоскелета и облегчают адгезию, миграцию и распространение нейтрофилов. [27] Они накапливаются асимметрично плазматической мембране на переднем крае поляризованных клеток. Пространственно регулируя Rho-GTPases и организуя передний край клетки, PI3K и их липидные продукты могут играть ключевую роль в установлении полярности лейкоцитов, выступая в качестве молекул-компасов, которые сообщают клетке, куда ползти.
На мышах было показано, что в определенных условиях нейтрофилы обладают особым типом миграционного поведения, называемым роением нейтрофилов , во время которого они мигрируют высокоскоординированным образом, накапливаются и группируются в участках воспаления. [28]
Антимикробная функция
Обладая высокой подвижностью , нейтрофилы быстро собираются в очаге инфекции , привлекаемые цитокинами , экспрессируемыми активированным эндотелием , тучными клетками и макрофагами . Нейтрофилы экспрессируют [29] и высвобождают цитокины, которые, в свою очередь, усиливают воспалительные реакции со стороны некоторых других типов клеток.
Помимо рекрутирования и активации других клеток иммунной системы, нейтрофилы играют ключевую роль в защите на передовой от вторжения патогенов. У нейтрофилов есть три метода непосредственной атаки на микроорганизмы: фагоцитоз (проглатывание), дегрануляция (высвобождение растворимых антимикробных препаратов) и образование внеклеточных ловушек нейтрофилов (NET). [30]
Нейтрофилы — это фагоциты , способные поглощать микроорганизмы или частицы. Чтобы мишени были распознаны, они должны быть покрыты опсонинами – процесс, известный как опсонизация антител . [17] Они могут усваивать и убивать многие микробы , каждое событие фагоцитирования приводит к образованию фагосомы, в которую секретируются активные формы кислорода и гидролитические ферменты. Потребление кислорода во время образования активных форм кислорода было названо « дыхательным взрывом », хотя оно не связано с дыханием или производством энергии.
Респираторный взрыв включает активацию фермента НАДФН -оксидазы , который производит большое количество супероксида , активной формы кислорода. Супероксид распадается самопроизвольно или расщепляется с помощью ферментов, известных как супероксиддисмутазы (Cu/ZnSOD и MnSOD), до перекиси водорода, которая затем превращается в хлорноватистую кислоту (HClO) под действием фермента зеленого гема миелопероксидазы . Считается, что бактерицидных свойств HClO достаточно, чтобы убить бактерии, фагоцитируемые нейтрофилами, но вместо этого это может быть шагом, необходимым для активации протеаз. [31]
Хотя нейтрофилы могут убивать многие микробы, взаимодействие нейтрофилов с микробами и молекулами, продуцируемыми микробами, часто изменяет оборот нейтрофилов. Способность микробов изменять судьбу нейтрофилов весьма разнообразна, может быть микроб-специфичной и варьируется от продления продолжительности жизни нейтрофилов до быстрого лизиса нейтрофилов после фагоцитоза. Сообщалось, что Chlamydia pneumoniae и Neisseria gonorrhoeae замедляют апоптоз нейтрофилов . [32] [33] [34] Таким образом, некоторые бактерии – и те, которые являются преимущественно внутриклеточными патогенами – могут продлить продолжительность жизни нейтрофилов, нарушая нормальный процесс спонтанного апоптоза и/или PICD (гибель клеток, вызванная фагоцитозом). На другом конце спектра находятся некоторые патогены, такие как Streptococcus pyogenes, которые способны изменять судьбу нейтрофилов после фагоцитоза, способствуя быстрому лизису клеток и/или ускоряя апоптоз до точки вторичного некроза. [35] [36]
Дегрануляция
Нейтрофилы также выделяют ряд белков в трех типах гранул в процессе, называемом дегрануляцией . Содержимое этих гранул обладает противомикробными свойствами и помогает бороться с инфекцией. Блестящие клетки представляют собой полиморфно-ядерные лейкоцитарные нейтрофилы с гранулами. [37]
Нейтрофильные внеклеточные ловушки
В 2004 году Бринкманн и его коллеги описали поразительное наблюдение: активация нейтрофилов вызывает высвобождение паутинных структур ДНК; это представляет собой третий механизм уничтожения бактерий. [39] Эти нейтрофильные внеклеточные ловушки (NET) представляют собой сеть волокон, состоящих из хроматина и сериновых протеаз [40] , которые улавливают и убивают внеклеточные микробы. Предполагается, что NET обеспечивают высокую локальную концентрацию антимикробных компонентов и связывают, обезвреживают и убивают микробы независимо от фагоцитарного поглощения. Помимо возможных антимикробных свойств, сети могут служить физическим барьером, предотвращающим дальнейшее распространение патогенов. Улавливание бактерий может играть особенно важную роль для НЭО при сепсисе , когда НЭО образуются внутри кровеносных сосудов. [41] Наконец, было продемонстрировано, что образование NET увеличивает бактерицидную активность макрофагов во время инфекции. [42] [43] Недавно было показано, что НЭО играют роль в воспалительных заболеваниях, поскольку НЭО могут быть обнаружены при преэклампсии , воспалительном заболевании, связанном с беременностью, при котором, как известно, активируются нейтрофилы. [44] Образование нейтрофильных сетей может также влиять на сердечно-сосудистые заболевания , поскольку сети могут влиять на образование тромбов в коронарных артериях . [45] [46]
Сейчас известно, что НЭО проявляют протромботические эффекты как in vitro [47], так и in vivo . [48] [49] Совсем недавно, в 2020 году, НЭО были вовлечены в образование тромбов в случаях тяжелого течения COVID-19 . [50]
Нейтрофилы, ассоциированные с опухолью
TAN могут проявлять повышенную скорость внеклеточного закисления при повышении уровня гликолиза. [51] Когда происходит метаболический сдвиг в TANS, это может привести к прогрессированию опухоли в определенных областях тела, таких как легкие. В 2021 году Анси и др. определили, как TAN поддерживают рост и прогрессирование опухолей в отличие от нормальных нейтрофилов, которые подавляют прогрессирование опухоли посредством фагоцитоза опухолевых клеток. Используя модель на мышах, они определили, что метаболизм Glut1 и глюкозы увеличивается в TAN, обнаруженных у мышей с аденокарциномой легких. [51]
Клиническое значение
Микрофотография, показывающая несколько нейтрофилов во время острого воспаления.
Низкое количество нейтрофилов называется нейтропенией . Это может быть врожденным (развилось при рождении или до него) или позднее, как в случае апластической анемии или некоторых видов лейкемии . Это также может быть побочным эффектом лекарств , особенно химиотерапии . Нейтропения делает человека очень восприимчивым к инфекциям. Это также может быть результатом колонизации внутриклеточными нейтрофильными паразитами.
При дефиците альфа-1-антитрипсина важная эластаза нейтрофилов не ингибируется должным образом альфа-1-антитрипсином , что приводит к чрезмерному повреждению тканей при наличии воспаления, наиболее выраженным из которых является эмфизема . Отрицательное действие эластазы было показано также в тех случаях, когда нейтрофилы чрезмерно активируются (у здоровых людей) и выделяют фермент во внеклеточное пространство. Нерегулируемая активность нейтрофильной эластазы может привести к нарушению легочного барьера, проявляясь симптомами, соответствующими острому повреждению легких . [52] Фермент также влияет на активность макрофагов, расщепляя их toll-подобные рецепторы (TLR) и подавляя экспрессию цитокинов путем ингибирования ядерной транслокации NF-κB . [53]
Гипергликемия может привести к дисфункции нейтрофилов. Дисфункция нейтрофильного биохимического пути миелопероксидазы , а также снижение дегрануляции связаны с гипергликемией. [55]
Абсолютное количество нейтрофилов (АНК) также используется в диагностике и прогнозе. АНК является золотым стандартом для определения тяжести нейтропении и, следовательно, нейтропенической лихорадки. Любой АНК < 1500 клеток/мм 3 считается нейтропенией, а <500 клеток/мм 3 - тяжелой формой. [56] Есть также новое исследование, связывающее АНК с инфарктом миокарда как средство ранней диагностики. [57] [58] Нейтрофилы способствуют желудочковой тахикардии при остром инфаркте миокарда. [59]
При аутопсии наличие нейтрофилов в сердце или головном мозге является одним из первых признаков инфаркта и полезно для определения времени и диагностики инфаркта миокарда и инсульта .
При инсульте они начинают проникать в пораженный мозг через 6–8 часов. [61]
Нейтрофильные антигены
Распознается пять (HNA 1–5) наборов антигенов нейтрофилов. [62] Три антигена HNA-1 (ac) расположены на низкоаффинном рецепторе Fc-γ IIIb (FCGR3B: CD16b ). Единственный известный антиген HNA-2a расположен на CD177 . Антигенная система HNA-3 включает два антигена (3a и 3b), которые расположены на седьмом экзоне гена CLT2 ( SLC44A2 ). Каждая антигенная система HNA-4 и HNA-5 имеет по два известных антигена (a и b) и расположена в интегрине β2 . HNA-4 расположен на цепи αM ( CD11b ), а HNA-5 расположен на единице интегрина αL ( CD11a ).
Субпопуляции
Активность нейтрофилов-киллеров и нейтрофилов-клеток в тесте НБТ [63]
Выявлены две функционально неравные субпопуляции нейтрофилов на основании разного уровня образования ими активных метаболитов кислорода, проницаемости мембран, активности ферментной системы и способности к инактивации. Клетки одной субпопуляции с высокой проницаемостью мембран (нейтрофилы-киллеры) интенсивно генерируют активные метаболиты кислорода и инактивируются вследствие взаимодействия с субстратом, тогда как клетки другой субпопуляции (нейтрофилы-клетки) менее интенсивно продуцируют активные формы кислорода, не прикрепляются к субстрату и сохраняют свою активность. [63] [64] [65] [66] [67] Дополнительные исследования показали, что опухоли легких могут быть инфильтрированы различными популяциями нейтрофилов. [68]
видео
Можно увидеть быстро движущийся нейтрофил, занимающий несколько конидий в течение времени визуализации 2 часа с одним кадром каждые 30 секунд.
Здесь можно увидеть нейтрофил, избирательно поглощающий несколько дрожжей Candida ( флуоресцентно помеченных зеленым), несмотря на несколько контактов с конидиями Aspergillus fumigatus (немечеными, белыми/прозрачными) в трехмерной коллагеновой матрице. Время съемки составляло 2 часа по одному кадру каждые 30 секунд.
Нейтрофилы проявляют направленную амебоидную подвижность в инфицированных подушечках лап и фалангах. Прижизненную визуализацию проводили на пути подушечек лап мышей LysM-eGFP через 20 минут после заражения Listeria monocytogenes . [69]
^ Эрмерт Д., Ниемец М.Дж., Рем М., Глентхой А., Боррегор Н., Урбан CF (август 2013 г.). «Candida albicans ускользает от нейтрофилов мыши». Журнал биологии лейкоцитов . 94 (2): 223–236. дои : 10.1189/jlb.0213063. PMID 23650619. S2CID 25619835.
^ Витко-Сарсат В., Рье П., Декамп-Лача Б., Лесавр П., Хальбвакс-Мекарелли Л. (май 2000 г.). «Нейтрофилы: молекулы, функции и патофизиологические аспекты». Лабораторные исследования; Журнал технических методов и патологии . 80 (5): 617–653. дои : 10.1038/labinvest.3780067 . PMID 10830774. S2CID 22536645.
^ Клебанофф С.Дж., Кларк Р.А. (1978). Нейтрофил: функции и клинические расстройства . Эльзевир/Северная Голландия, Амстердам. ISBN978-0-444-80020-6.
^ Натан С. (март 2006 г.). «Нейтрофилы и иммунитет: проблемы и возможности». Обзоры природы. Иммунология . 6 (3): 173–182. дои : 10.1038/nri1785. PMID 16498448. S2CID 1590558.
^ Уэлш, Чарльз Дж. (2021). Основы анатомии и физиологии человека Хоула (14-е изд.). Нью-Йорк, США: МакГроу Хилл. п. 336. ИСБН978-1-260-57521-7. Проверено 28 февраля 2023 г.
^ Джейкобс Л., Наврот Т.С., де Геус Б., Меусен Р., Дегреуле Б., Бернар А. и др. (октябрь 2010 г.). «Субклинические реакции у здоровых велосипедистов, кратковременно подвергавшихся воздействию загрязнения воздуха, связанного с дорожным движением: интервенционное исследование». Состояние окружающей среды . 9 (64): 64. дои : 10.1186/1476-069X-9-64 . ПМЦ 2984475 . ПМИД 20973949.
^ Во DJ, Уилсон С. (ноябрь 2008 г.). «Путь интерлейкина-8 при раке». Клинические исследования рака . 14 (21): 6735–6741. дои : 10.1158/1078-0432.CCR-07-4843. PMID 18980965. S2CID 9415085.
^ Де Ларко Дж. Э., Вюрц БР, Фурхт Л.Т. (август 2004 г.). «Потенциальная роль нейтрофилов в продвижении метастатического фенотипа опухолей, высвобождающих интерлейкин-8». Клинические исследования рака . 10 (15): 4895–4900. дои : 10.1158/1078-0432.CCR-03-0760. PMID 15297389. S2CID 9782495.
^ Ю СК, Старнс Т.В., Дэн К., Хуттенлохер А. (ноябрь 2011 г.). «Lyn — это окислительно-восстановительный сенсор, который обеспечивает притяжение лейкоцитов к ране in vivo». Природа . 480 (7375): 109–112. Бибкод : 2011Natur.480..109Y. дои : 10.1038/nature10632. ПМЦ 3228893 . ПМИД 22101434.
^ Коэн С., Бернс RC (2002). Пути пульпы (8-е изд.). Сент-Луис: Мосби. п. 465.
^ Ниемец М.Дж., Де Самбер Б., Гарревет Дж., Вергухт Э., Векеманс Б., Де Райке Р. и др. (июнь 2015 г.). «Микроэлементный ландшафт покоящихся и активированных нейтрофилов человека на субмикрометровом уровне». Металломика . 7 (6): 996–1010. дои : 10.1039/c4mt00346b . ПМИД 25832493.
^ abc Цукер-Франклин Д., Гривз М.Ф., Гросси CE, Мармонт AM (1988). «Нейтрофилы». Атлас клеток крови: функции и патология . Том. 1 (2-е изд.). Филадельфия: Леа и Фербигер. ISBN978-0-8121-1094-4.
^ Карни Р.Дж., Ван Л.Дж., Санчес Дж.А. (август 2001 г.). «Неслучайное расположение и ориентация неактивной Х-хромосомы в ядрах нейтрофилов человека». Хромосома . 110 (4): 267–274. дои : 10.1007/s004120100145. PMID 11534818. S2CID 24750407.
^ Райх Д., Ноллс М.А., Као WH, Акылбекова Е.Л., Тандон А., Паттерсон Н. и др. (январь 2009 г.). «Снижение количества нейтрофилов у людей африканского происхождения связано с регуляторным вариантом рецептора антигена Даффи для гена хемокинов». ПЛОС Генетика . 5 (1): e1000360. дои : 10.1371/journal.pgen.1000360 . ПМЦ 2628742 . ПМИД 19180233.
^ аб Эдвардс SW (1994). Биохимия и физиология нейтрофилов . Издательство Кембриджского университета. п. 6. ISBN978-0-521-41698-6.
^ Санчес А., Ризер Дж.Л., Лау Х.С., Яхику П.Ю., Уиллард Р.Э., Макмиллан П.Дж. и др. (ноябрь 1973 г.). «Роль сахаров в нейтрофильном фагоцитозе человека». Американский журнал клинического питания . 26 (11): 1180–1184. дои : 10.1093/ajcn/26.11.1180 . PMID 4748178. Эти данные позволяют предположить, что при приеме сахаров изменялась функция, а не количество фагоцитов. Это указывает на участие глюкозы и других простых углеводов в контроле фагоцитоза и показывает, что эффект длится не менее 5 часов. С другой стороны, голодание в течение 36 или 60 часов значительно повышало (P <0,001) фагоцитарный индекс.
^ Рубин-Бейерано I, Абейон С, Магнелли П, Грисафи П, Финк GR (июль 2007 г.). «Фагоцитоз нейтрофилов человека стимулируется уникальным компонентом клеточной стенки грибов». Клетка-хозяин и микроб . 2 (1): 55–67. doi :10.1016/j.chom.2007.06.002. ПМК 2083279 . ПМИД 18005717.
^ Кнеллер А (2007). «Белые кровяные тельца требовательны к сахару». Институт Уайтхеда . Проверено 9 августа 2013 г.
^ Так Т, Тесселаар К., Пиллэй Дж., Борганс Дж.А., Кундерман Л. (октябрь 2013 г.). «Сколько вам еще лет? Еще раз об определении периода полураспада нейтрофилов человека». Журнал биологии лейкоцитов . 94 (4): 595–601. дои : 10.1189/jlb.1112571. PMID 23625199. S2CID 40113921.
^ Пиллэй Дж., ден Брабер И., Врисекуп Н., Кваст Л.М., де Бур Р.Дж., Борганс Дж.А. и др. (июль 2010 г.). «Метки in vivo с помощью 2H2O показывают, что продолжительность жизни нейтрофилов человека составляет 5,4 дня». Кровь . 116 (4): 625–627. doi : 10.1182/blood-2010-01-259028 . PMID 20410504. S2CID 909519.
^ ab Wheater PR, Стивенс А. (2002). Основная гистопатология Уитера: цветной атлас и текст . Эдинбург: Черчилль Ливингстон. ISBN978-0-443-07001-3.
^ Акбар, Навид; Брейтуэйт, Адам Т; Корр, Эмма М; Коэлвин, Грэм Дж; ван Золинген, Коэн; Кошен, Клеман; Салиба, Антуан-Эммануэль; Корбин, Аластер; Пеццолла, Даниэла; Мёллер Йоргенсен, Мален; Бэк, Рикке; Эдгар, Лориен; Де Вильерс, Карла; Гунадаса-Ролинг, Мала; Банерджи, Абхируп; Пэджет, Даан; Ли, Шарлотта; Хогг, Элеонора; Костин, Адам; Даливал, Раман; Джонсон, Эррин; Краусгрубер, Томас; Рипсааме, Джоуи; Меллинг, Женевьева Э; Шанмуганатан, Майоран; Баннинг, Адриан; Харбанда, Радж; Рупарелия, Нил; Алхалил, Мохаммед; Де Мария, ДжанЛюджи; Гогран, Лиза; Далл'Армеллина, Эрика; Феррейра, Ванесса; Борлотти, Алессандра; Нг, Юджун; Бок, Кристоф; Картер, Дэвид РФ; Ченнон, Кейт М; Райли, Пол Р.; Удалова Ирина А; Мур, Кэтрин Дж; Энтони, Дэниел С; Чоудри, Робин П. (4 февраля 2022 г.). «Быстрая мобилизация нейтрофилов с помощью внеклеточных везикул, происходящих из эндотелиальных клеток VCAM-1+». Сердечно-сосудистые исследования . 119 : 236–251. doi : 10.1093/cvr/cvac012. ПМЦ 10022859 . ПМИД 35134856.
^ Риттер, Уве; Фришкнехт, Фредди; ван Зандберген, Гер (2009). «Являются ли нейтрофилы важными клетками-хозяевами для паразитов Leishmania?». Тенденции в паразитологии . 25 (11): 505–510. дои : 10.1016/j.pt.2009.08.003.
^ Серхан CN, Уорд П.А., Гилрой Д.В. (2010). Основы воспаления. Издательство Кембриджского университета. стр. 53–54. ISBN978-0-521-88729-8.
^ Пантарелли С., Уэлч ХК (ноябрь 2018 г.). «Rac-GTPases и Rac-GEF в адгезии, миграции и рекрутировании нейтрофилов». Европейский журнал клинических исследований . 48 (Приложение 2): e12939. дои : 10.1111/eci.12939. ПМК 6321979 . ПМИД 29682742.
^ Леммерманн Т., Афонсо П.В., Ангерманн Б.Р., Ван Дж.М., Кастенмюллер В., Родитель Калифорния, Жермен Р.Н. (июнь 2013 г.). «Рои нейтрофилов требуют LTB4 и интегринов в местах гибели клеток in vivo». Природа . 498 (7454): 371–375. Бибкод : 2013Natur.498..371L. дои : 10.1038/nature12175. ПМЦ 3879961 . ПМИД 23708969.
^ Ear T, McDonald PP (апрель 2008 г.). «Генерация цитокинов, активация промотора и независимая от оксиданта активация NF-kappaB в трансфицируемой модели нейтрофильных клеток человека». БМК Иммунология . 9:14 . дои : 10.1186/1471-2172-9-14 . ПМК 2322942 . ПМИД 18405381.
^ Hickey MJ, Kubes P (май 2009 г.). «Внутрисосудистый иммунитет: встреча хозяина и возбудителя в кровеносных сосудах». Обзоры природы. Иммунология . 9 (5): 364–375. дои : 10.1038/nri2532. PMID 19390567. S2CID 8068543.
^ Саймонс, член парламента, Наусиф В.М., Гриффит Т.С., Апичелла М.А. (ноябрь 2006 г.). «Neisseria gonorrhoeae задерживает начало апоптоза в полиморфно-ядерных лейкоцитах». Клеточная микробиология . 8 (11): 1780–1790. дои : 10.1111/j.1462-5822.2006.00748.x. PMID 16803582. S2CID 25253422.
^ Чен А, Зейферт HS (ноябрь 2011 г.). «Опосредованное Neisseria gonorrhoeae ингибирование апоптотической передачи сигналов в полиморфно-ядерных лейкоцитах». Инфекция и иммунитет . 79 (11): 4447–4458. дои : 10.1128/IAI.01267-10. ПМК 3257915 . ПМИД 21844239.
^ ван Зандберген Г., Гифферс Дж., Коте Х., Рупп Дж., Боллинджер А., Ага Е. и др. (февраль 2004 г.). «Chlamydia pneumoniae размножается в нейтрофильных гранулоцитах и задерживает их спонтанный апоптоз». Журнал иммунологии . 172 (3): 1768–1776. doi : 10.4049/jimmunol.172.3.1768 . PMID 14734760. S2CID 27422510.
^ Кобаяши С.Д., Бротон К.Р., Уитни А.Р., Войич Дж.М., Шван Т.Г., Массер Дж.М., ДеЛео Ф.Р. (сентябрь 2003 г.). «Бактериальные патогены модулируют программу дифференцировки апоптоза в нейтрофилах человека». Труды Национальной академии наук Соединенных Штатов Америки . 100 (19): 10948–10953. дои : 10.1073/pnas.1833375100 . ЧВК 196908 . ПМИД 12960399.
^ Кобаяши С.Д., Малахова Н., ДеЛео ФР (2017). «Влияние микробов на жизнь и смерть нейтрофилов». Границы клеточной и инфекционной микробиологии . 7 (159): 159. дои : 10.3389/fcimb.2017.00159 . ПМК 5410578 . ПМИД 28507953.Материал был скопирован из этого источника, который доступен по международной лицензии Creative Commons Attribution 4.0.
^ Берман, LB; Шрайнер, GE; Фейс, Дж.О. (22 ноября 1956 г.). «Наблюдения за феноменом блестящих клеток». Медицинский журнал Новой Англии . 255 (21): 989–991. дои : 10.1056/NEJM195611222552104. ISSN 0028-4793. ПМИД 13378597.
^ Бринкманн В., Райхард У., Гусманн С., Фаулер Б., Улеманн Ю., Вайс Д.С. и др. (март 2004 г.). «Нейтрофильные внеклеточные ловушки убивают бактерии». Наука . 303 (5663): 1532–1535. Бибкод : 2004Sci...303.1532B. дои : 10.1126/science.1092385. PMID 15001782. S2CID 21628300.
^ Урбан К.Ф., Эрмерт Д., Шмид М., Абу-Абед У., Гусманн С., Накен В. и др. (октябрь 2009 г.). «Внеклеточные ловушки нейтрофилов содержат кальпротектин, цитозольный белковый комплекс, участвующий в защите хозяина от Candida albicans». ПЛОС Патогены . 5 (10): e1000639. дои : 10.1371/journal.ppat.1000639 . ПМЦ 2763347 . ПМИД 19876394.
^ Кларк С.Р., Ма А.К., Тавенер С.А., Макдональд Б., Гударзи З., Келли М.М. и др. (апрель 2007 г.). «Тромбоциты TLR4 активируют внеклеточные ловушки нейтрофилов, чтобы поймать бактерии в гнойную кровь». Природная медицина . 13 (4): 463–469. дои : 10.1038/нм1565. PMID 17384648. S2CID 22372863.
^ Монтейт, Эндрю Дж.; Миллер, Жанетт М.; Максвелл, К. Ноэль; Чазин, Уолтер Дж.; Скаар, Эрик П. (10 сентября 2021 г.). «Внеклеточные ловушки нейтрофилов усиливают уничтожение бактериальных патогенов макрофагами». Достижения науки . 7 (37): eabj2101. Бибкод : 2021SciA....7.2101M. doi : 10.1126/sciadv.abj2101. ISSN 2375-2548. ПМЦ 8442908 . ПМИД 34516771.
^ Монтейт, Эндрю Дж.; Миллер, Жанетт М.; Биверс, Уильям Н.; Мэлони, К. Николь; Зайферт, Эрин Л.; Хайноцкий, Дьердь; Скаар, Эрик П. (06 декабря 2021 г.). «Унипортер митохондриального кальция влияет на бактерицидную активность нейтрофилов при инфекции Staphylococcus aureus». Инфекция и иммунитет . 90 (2): IAI.00551–21. дои : 10.1128/IAI.00551-21. ISSN 0019-9567. ПМЦ 8853686 . ПМИД 34871043.
^ Гупта А.К., Хаслер П., Хольцгрев В., Хан С. (июнь 2007 г.). «Нейтрофильные сети: новый вклад в плацентарную гипоксию, связанную с преэклампсией?» (PDF) . Семинары по иммунопатологии . 29 (2): 163–167. дои : 10.1007/s00281-007-0073-4. PMID 17621701. S2CID 12887059.
^ Хойер Ф.Ф., Нарендорф М. (февраль 2017 г.). «Вклад нейтрофилов в развитие ишемической болезни сердца». Европейский кардиологический журнал . 38 (7): 465–472. doi : 10.1093/eurheartj/ehx017 . ПМИД 28363210.
^ Мангольд А., Алиас С., Шерц Т., Хофбауэр М., Якович Дж., Панценбёк А. и др. (март 2015 г.). «Нагрузка внеклеточных ловушек коронарных нейтрофилов и активность дезоксирибонуклеазы при остром коронарном синдроме с подъемом сегмента ST являются предикторами разрешения сегмента ST и размера инфаркта». Исследование кровообращения . 116 (7): 1182–1192. дои : 10.1161/CIRCRESAHA.116.304944 . PMID 25547404. S2CID 2532741.
^ Фукс Т.А., Брилл А., Дюршмид Д., Шацберг Д., Монестье М., Майерс Д.Д. и др. (сентябрь 2010 г.). «Ловушки внеклеточной ДНК способствуют тромбозу». Труды Национальной академии наук Соединенных Штатов Америки . 107 (36): 15880–15885. Бибкод : 2010PNAS..10715880F. дои : 10.1073/pnas.1005743107 . ПМЦ 2936604 . ПМИД 20798043.
^ Брилл А., Фукс Т.А., Савченко А.С., Томас Г.М., Мартинод К., Де Мейер С.Ф. и др. (январь 2012 г.). «Внеклеточные ловушки нейтрофилов способствуют тромбозу глубоких вен у мышей». Журнал тромбозов и гемостаза . 10 (1): 136–144. дои : 10.1111/j.1538-7836.2011.04544.x. ПМК 3319651 . ПМИД 22044575.
^ Борисов JI, тен Кейт H (сентябрь 2011 г.). «От высвобождения внеклеточных ловушек нейтрофилов к тромбозу: избыточный механизм защиты хозяина?». Журнал тромбозов и гемостаза . 9 (9): 1791–1794. дои : 10.1111/j.1538-7836.2011.04425.x . PMID 21718435. S2CID 5368241.
^ Цзо, Ю и др. (2020). «Нейтрофильные внеклеточные ловушки при COVID-19». Журнал клинических исследований . 5 (11): e138999. doi : 10.1172/jci.insight.138999. ПМК 7308057 . PMID 32329756. S2CID 216109364 . Проверено 19 февраля 2022 г.
^ аб Анси, Пьер-Бенуа; Контат, Кэролайн; Бойвен, Гаэль; Сабатино, Сильвия; Паскуаль, Жюстин; Зангер, Надин; Перентес, Жан Яннис; Питерс, Соланж; Абель, Э.; Кирш, Дэвид; Ратмелл, Джеффри; Возенин, Мария-Катрин; Мейлан, Этьен (22 марта 2021 г.). «Экспрессия Glut1 в опухолеассоциированных нейтрофилах способствует росту рака легких и устойчивости к лучевой терапии». Исследования рака . 81 (9): 2345–2357. дои : 10.1158/0008-5472.CAN-20-2870. ПМЦ 8137580 . ПМИД 33753374.
^ Кавабата К., Хагио Т., Мацуока С. (сентябрь 2002 г.). «Роль нейтрофильной эластазы при остром повреждении легких». Европейский журнал фармакологии . 451 (1): 1–10. дои : 10.1016/S0014-2999(02)02182-9. ПМИД 12223222.
^ Домон Х., Нагай К., Маэкава Т., Ода М., Ёнезава Д., Такеда В. и др. (2018). «Нейтрофильная эластаза подавляет иммунный ответ путем расщепления толл-подобных рецепторов и цитокинов при пневмококковой пневмонии». Границы в иммунологии . 9 : 732. дои : 10.3389/fimmu.2018.00732 . ПМЦ 5996908 . ПМИД 29922273.
^ Озен С (июль 2003 г.). «Семейная средиземноморская лихорадка: возвращение к древней болезни». Европейский журнал педиатрии . 162 (7–8): 449–454. дои : 10.1007/s00431-003-1223-x. PMID 12751000. S2CID 3464945.
^ Сю Ф, Станойчич М, Диао Л, Ешке МГ (8 мая 2014 г.). «Стрессовая гипергликемия, лечение инсулином и врожденные иммунные клетки». Международный журнал эндокринологии . 2014 : 486403. doi : 10.1155/2014/486403 . ПМК 4034653 . ПМИД 24899891.
^ Аль-Гвайз Л.А., Бабай Х.Х. (2007). «Диагностическая ценность абсолютного количества нейтрофилов, количества полос и морфологических изменений нейтрофилов в прогнозировании бактериальных инфекций». Медицинские принципы и практика . 16 (5): 344–347. дои : 10.1159/000104806 . PMID 17709921. S2CID 5499290.
^ Хан Х.А., Алхомида А.С., Собки Ш.Х., Могайри А.А., Коронки Х.Э. (2012). «Количество клеток крови и их корреляция с креатинкиназой и С-реактивным белком у больных острым инфарктом миокарда». Международный журнал клинической и экспериментальной медицины . 5 (1): 50–55. ПМЦ 3272686 . ПМИД 22328948.
^ Базили С., Ди Франсуа М., Роза А., Феррони П., Дюрни В., Скарпеллини М.Г., Бертаццони Г. (апрель 2004 г.). «Абсолютное количество нейтрофилов и уровень фибриногена как помощь в ранней диагностике острого инфаркта миокарда». Акта Кардиологика . 59 (2): 135–140. дои : 10.2143/ac.59.2.2005167. PMID 15139653. S2CID 37382677.
^ Грюне, Яна; Льюис, Эндрю Дж. М.; Ямазоэ, Масахиро; Хульсманс, Мартен; Роде, Дэвид; Сяо, Лин; Чжан, Шуан; Отт, Кристиана; Кальканьо, Дэвид М.; Чжоу, Ижун; Тимм, Керстин (11 июля 2022 г.). «Нейтрофилы провоцируют, а макрофаги предотвращают грозу после инфаркта миокарда». Природные сердечно-сосудистые исследования . 1 (7): 649–664. doi : 10.1038/s44161-022-00094-w. ISSN 2731-0590. ПМЦ 9410341 . ПМИД 36034743.
^ Мишо К., Бассо С., д'Амати Дж., Джордано С., Холова I, Престон С.Д. и др. (февраль 2020 г.). «Диагностика инфаркта миокарда на аутопсии: переоценка AECVP в свете современной клинической классификации». Архив Вирхова . 476 (2): 179–194. дои : 10.1007/s00428-019-02662-1 . ПМК 7028821 . ПМИД 31522288. «Эта статья распространяется на условиях международной лицензии Creative Commons Attribution 4.0 (http://creativecommons.org/licenses/by/4.0/. Архивировано 21 ноября 2015 г. на Wayback Machine )»
^ Jickling GC, Лю Д., Андер Б.П., Стамова Б., Жан X, Sharp FR (июнь 2015 г.). «Нацеливание на нейтрофилы при ишемическом инсульте: выводы из экспериментальных исследований». Журнал церебрального кровотока и метаболизма . 35 (6): 888–901. дои : 10.1038/jcbfm.2015.45. ПМК 4640255 . ПМИД 25806703.
^ Чу Х.Т., Линь Х., Цао Т.Т., Чанг К.Ф., Сяо В.В., Йе Т.Дж. и др. (Сентябрь 2013). «Генотипирование нейтрофильных антигенов человека (HNA) на основе данных полногеномного секвенирования». BMC Медицинская Геномика . 6 (1): 31. дои : 10.1186/1755-8794-6-31 . ПМЦ 3849977 . ПМИД 24028078.
^ аб Игнатов Д.Ю. (2012). Функциональная гетерогенность нейтрофилов человека и их роль в регуляции количества лейкоцитов периферической крови (доктор философии). Донецкий национальный медицинский университет. дои : 10.13140/RG.2.2.35542.34884.
^ Герасимов ИГ, Игнатов ДИ (2001). «[Функциональная гетерогенность нейтрофилов крови человека: генерация активных форм кислорода]». Цитология . 43 (5): 432–436. ПМИД 11517658.
^ Герасимов ИГ, Игнатов ДИ (2004). «[Активация нейтрофилов in vitro]». Цитология . 46 (2): 155–158. ПМИД 15174354.
^ Герасимов И.Г., Игнатов Д.И., Котельницкий М.А. (2005). «[Восстановление нитросинего тетразолия нейтрофилами крови человека. I. Влияние pH]». Цитология . 47 (6): 549–553. ПМИД 16708848.
^ Герасимов ИГ, Игнатов ДИ (2005). «[Восстановление нитросинего тетразолия нейтрофилами крови человека. II. Влияние ионов натрия и калия]». Цитология . 47 (6): 554–558. ПМИД 16708849.
^ Зилионис Р., Энгблом С., Пфиршке С., Савова В., Земмур Д., Саатчиоглу HD и др. (май 2019 г.). «Одноклеточная транскриптомика рака легких человека и мыши показывает консервативные миелоидные популяции у отдельных людей и видов». Иммунитет . 50 (5): 1317–1334.e10. doi :10.1016/j.immuni.2019.03.009. ПМК 6620049 . ПМИД 30979687.
^ Грэм Д.Б., Зинсельмейер Б.Х., Маскаренхас Ф., Дельгадо Р., Миллер М.Дж., Сват В. (2009). Унутмаз Д (ред.). «Передача сигналов ITAM с помощью факторов обмена гуаниновых нуклеотидов Rho семейства Vav регулирует скорость интерстициального транзита нейтрофилов in vivo». ПЛОС ОДИН . 4 (2): е4652. Бибкод : 2009PLoSO...4.4652G. дои : 10.1371/journal.pone.0004652 . ПМЦ 2645696 . ПМИД 19247495.
Внешние ссылки
Информация о нейтропении
Калькулятор абсолютного количества нейтрофилов
Содержание и распределение микроэлементов нейтрофилов






_diagram_en.svg/1280px-Hematopoiesis_(human)_diagram_en.svg.png)
![Нейтрофилы наблюдаются при инфаркте миокарда примерно через 12–24 часа [60], как видно на этой микрофотографии.](http://upload.wikimedia.org/wikipedia/commons/thumb/6/6c/Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg/1280px-Histopathology_of_neutrophil_infiltration_in_myocardial_infarction.jpg)
![При инсульте они начинают проникать в пораженный мозг через 6–8 часов.[61]](http://upload.wikimedia.org/wikipedia/commons/thumb/1/1e/Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg/1280px-Histopathology_of_thalamus_infarction_at_approximately_24_hours,_high_magnification,_annotated.jpg)
