
Большинство млекопитающих являются живородящими , рождая живых детенышей. [1] Однако пять видов однопроходных, утконосы и ехидны , откладывают яйца. У однопроходных система определения пола отличается от большинства других млекопитающих. [2] В частности, половые хромосомы утконоса больше похожи на хромосомы курицы, чем на хромосомы териевых млекопитающих. [3]
Молочные железы млекопитающих специализированы для выработки молока, жидкости, используемой новорожденными в качестве основного источника питания. Однопроходные рано отделились от других млекопитающих и не имеют сосков, которые можно увидеть у большинства млекопитающих, но у них есть молочные железы. Детеныши слизывают молоко с молочного пятна на животе матери.
Живородящие млекопитающие относятся к подклассу Theria ; ныне живущие относятся к инфраклассам Marsupialia и Placentalia . У сумчатых короткий период беременности , как правило, короче эстрального цикла , и они рождают недоразвитых ( альтрициальных ) новорожденных , которые затем проходят дальнейшее развитие; у многих видов это происходит в мешковидном мешке, марсупиуме , расположенном в передней части живота матери . Некоторые плацентарные , например морская свинка , рождают полностью развитых ( выводковых ) детенышей , как правило, после длительных периодов беременности , в то время как некоторые другие , например мышь , рождают недоразвитых детенышей.
Половая зрелость и, следовательно, самый ранний возраст, в котором млекопитающие могут размножаться, значительно различаются у разных видов. Представители семейства грызунов Cricetidae могут достигать половой зрелости за 1–2 месяца, например, норвежский лемминг ( Lemmus lemmus ) за 39 дней. Многим собакам ( семейство Canidae ) и полорогим ( Bovidae ) требуется около года, чтобы достичь зрелости, в то время как приматам (включая людей ) и дельфинам ( Delphinidae ) требуется более 10 лет. Некоторым китам требуется еще больше времени, причем самая длительная продолжительность зарегистрирована для гренландского кита ( Balaena mysticetus ), который достигает зрелости в возрасте всего около 23 лет. [4]
Мужская репродуктивная система млекопитающих состоит из двух основных отделов: пениса и яичек , последний из которых является местом, где вырабатывается сперма, и обычно удерживается в мошонке . У людей оба этих органа находятся за пределами брюшной полости , но у других животных они могут в основном размещаться внутри брюшной полости. Например, пенис собаки покрыт пенильной оболочкой, за исключением случаев спаривания. Расположение яичек за пределами брюшной полости наилучшим образом способствует температурной регуляции спермы, которой для выживания требуются определенные температуры. Внешнее расположение может также привести к снижению вклада, вызванного теплом, в спонтанную скорость мутаций в мужской зародышевой ткани. [5] Сперма является меньшей из двух гамет и, как правило, очень недолговечна, что требует от самцов непрерывного производства ее с момента половой зрелости до смерти. Выработанная сперма хранится в придатках яичек до эякуляции через семявыносящий проток . Сперматозоиды подвижны и плавают, используя хвостовидные жгутики , чтобы продвигаться к яйцеклетке . Сперматозоид следует температурным градиентам ( термотаксис ) [6] и химическим градиентам ( хемотаксис ), чтобы найти яйцеклетку.
Женская репродуктивная система млекопитающих состоит из трех основных отделов: влагалище и матка , которые действуют как вместилище для спермы, яичники , которые производят яйцеклетки самки , и вульва , которая состоит из половых губ , преддверия и клитора . Влагалище, матка и яичники всегда внутренние, а вульва — наружная. Влагалище прикреплено к матке через шейку матки , в то время как матка прикреплена к яичникам через яйцеводы . Через определенные промежутки времени яичники выпускают яйцеклетку, которая проходит через яйцевод в матку.
Если при этом переходе она встречается со сперматозоидом , яйцеклетка выбирает сперматозоид, с которым она сливается; это называется оплодотворением . Оплодотворение обычно происходит в яйцеводах, но может произойти и в самой матке. Затем зигота имплантируется в стенку матки, где она начинает процессы эмбриогенеза и морфогенеза . Когда она достаточно развита, чтобы выжить вне матки, шейка матки расширяется, и сокращения матки продвигают плод через родовой канал, которым является влагалище.
Яйцеклетки, которые являются женскими половыми клетками, намного больше сперматозоидов и обычно образуются в яичниках плода до его рождения. Они в основном фиксируются в яичнике до своего перехода в матку и содержат питательные вещества для более поздней зиготы и эмбриона . В течение регулярного интервала времени в ответ на гормональные сигналы процесс оогенеза созревает одна яйцеклетка, которая высвобождается и отправляется вниз по яйцеводу. Если эта яйцеклетка не оплодотворена, она высвобождается через менструацию у людей и других человекообразных обезьян и реабсорбируется у других млекопитающих в цикле течки .
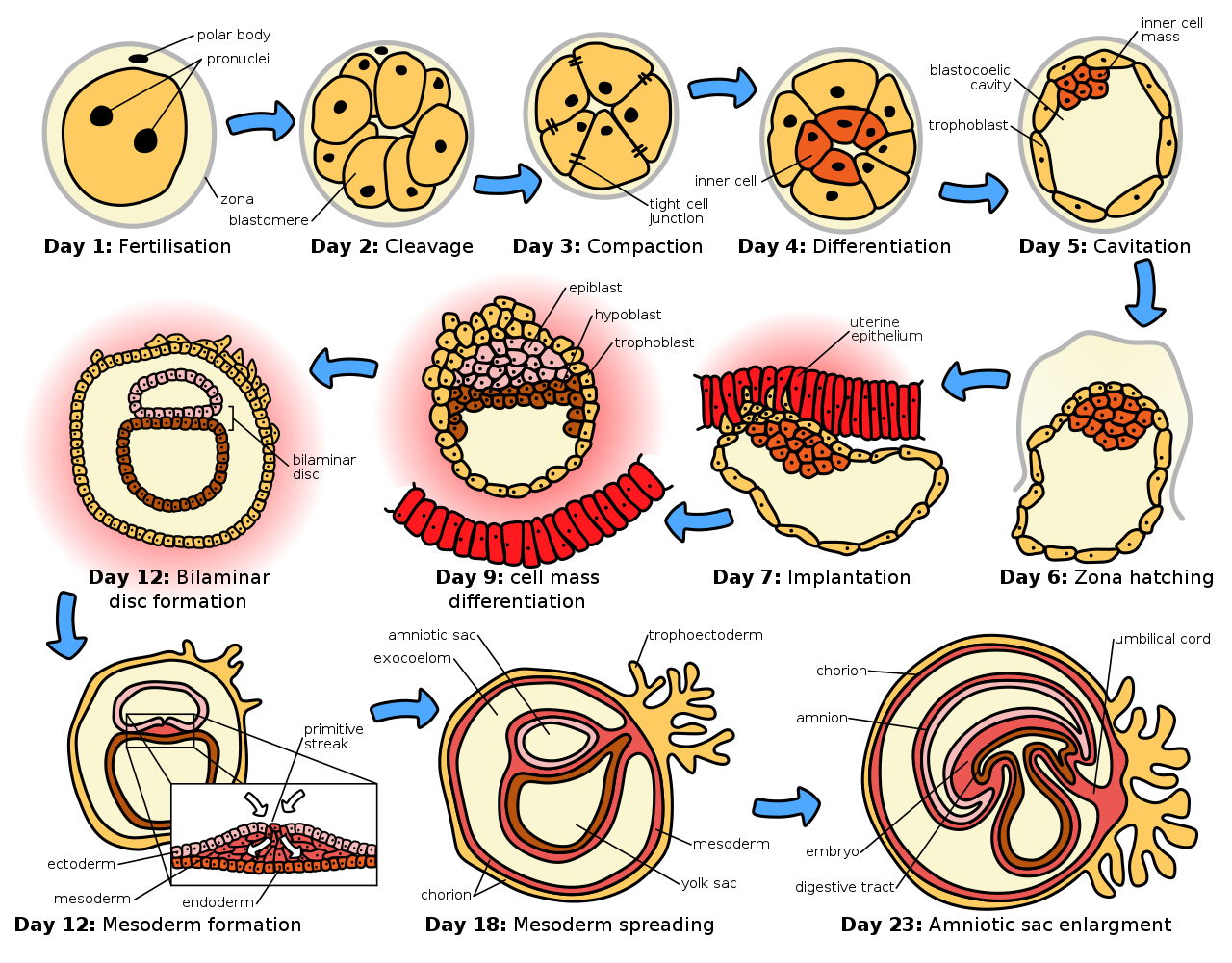
Гестация , называемая беременностью у людей, — это период времени, в течение которого плод развивается, делясь посредством митоза внутри самки. В это время плод получает все свое питание и насыщенную кислородом кровь от самки, отфильтрованную через плаценту , которая прикреплена к животу плода с помощью пуповины . Этот отток питательных веществ может быть довольно обременительным для самки, которой требуется потреблять немного больше калорий . Кроме того, некоторые витамины и другие питательные вещества требуются в больших количествах, чем обычно, что часто создает ненормальные привычки в еде. Продолжительность беременности, называемая периодом беременности , сильно различается у разных видов; у людей она составляет 40 недель, у жирафов — 56–60, а у хомяков — 16 дней .
Как только плод достаточно разовьется, химические сигналы запускают процесс рождения. Это начинается с сокращений матки и расширения шейки матки. Затем плод опускается к шейке матки, где он выталкивается во влагалище и, в конечном итоге, из женщины. Новорожденный, которого люди называют младенцем , обычно должен начать дышать самостоятельно вскоре после рождения. Вскоре после этого также выходит плацента.
Человеческие младенцы уникальны в животном мире из-за большого размера головы по сравнению с телом. Это влияет на процесс родов у людей, поскольку двуногая походка человека делает родовой канал относительно узким и скрученным посередине. В результате подавляющее большинство человеческих младенцев должны вращаться внутри родового канала, чтобы протиснуться через родовой канал и пройти через тазовые плоскости. Этот процесс известен как ротационные роды, и хотя это не уникальный процесс для людей, люди уникальны тем, что почти все человеческие младенцы проходят этот процесс по необходимости. Основная гипотеза о том, почему происходит этот процесс и другие, в результате чего роды у людей значительно сложнее, чем у других млекопитающих, известна как акушерская дилемма . [7]
Однопроходные , которых существует всего пять видов, все из Австралии и Новой Гвинеи , являются млекопитающими, которые откладывают яйца . У них есть одно отверстие для выделения и размножения, называемое клоакой . Они держат яйца внутри в течение нескольких недель, обеспечивая их питательными веществами, а затем откладывают их и покрывают, как птицы . Как и сумчатые « джоуи », однопроходные « паггли » являются личинками и плодоподобными, [8] поскольку, как и они, они не могут расширять свое туловище из-за наличия надлобковых костей, что заставляет их производить неразвитых детенышей.
Репродуктивная система сумчатых заметно отличается от таковой плацентарных млекопитающих, [9] [10] хотя это, вероятно, плезиоморфное состояние, обнаруженное у живородящих млекопитающих, включая неплацентарных плацентарных . [11] Во время эмбрионального развития у всех сумчатых формируется хориовителлиновая плацента . У бандикутов формируется дополнительная хориоаллантоисная плацента , хотя у нее отсутствуют хорионические ворсинки , обнаруженные в плацентах плацентарных .


Животные, включая млекопитающих, производят гаметы (сперму и яйцеклетку) посредством мейоза в гонадах (яички у самцов и яичники у самок). Сперматозоиды производятся в процессе сперматогенеза , а яйцеклетки производятся в процессе оогенеза . Эти процессы описаны в статье гаметогенез . Во время гаметогенеза у млекопитающих многие гены, кодирующие белки, которые принимают участие в механизмах репарации ДНК, демонстрируют усиленную или специализированную экспрессию [12]. Эти механизмы включают мейотическую гомологичную рекомбинационную репарацию и репарацию несоответствий .
Сексуальное поведение можно классифицировать на поведенческие состояния, связанные с мотивацией вознаграждения (« желание »), удовлетворением вознаграждения, также известным как удовольствие («симпатия»), и насыщением («торможение»); [13] эти поведенческие состояния регулируются у млекопитающих сексуальным обучением на основе вознаграждения, колебаниями различных нейрохимических веществ (например, дофамина − сексуальное желание , также известное как «желание»; норадреналина − сексуальное возбуждение ; окситоцина и меланокортинов − сексуальное влечение ), и циклами половых гормонов, а также находятся под влиянием половых феромонов и двигательных рефлексов (например, лордозное поведение ) у некоторых млекопитающих. [13] [14]
Эти поведенческие состояния коррелируют с фазами цикла сексуальной реакции человека : мотивация − возбуждение; завершение − плато и оргазм; насыщение − рефракция. [13] Сексуальное обучение (форма ассоциативного обучения ) происходит, когда животное начинает ассоциировать телесные особенности, личность, контекстуальные сигналы и другие стимулы с сексуальным удовольствием, вызванным генитально. [13] [14] После формирования эти ассоциации, в свою очередь, влияют как на сексуальное желание, так и на сексуальное влечение.
У большинства самок млекопитающих акт совокупления контролируется несколькими врожденными нейробиологическими процессами, включая двигательный половой рефлекс лордоза. [15] У самцов акт совокупления более сложен, поскольку необходимо некоторое обучение, но врожденные процессы (ретроконтроль введения пениса во влагалище , ритмическое движение таза, обнаружение женских феромонов) специфичны для совокупления. Эти врожденные процессы направляют гетеросексуальное совокупление. [ 16] Женское лордозное поведение стало вторичным у гоминидов и нефункционально у людей. [17] Млекопитающие обычно совокупляются в дорсовентральной позе, хотя некоторые виды приматов совокупляются в вентро-вентральной позе. [18] [19]
Большинство млекопитающих обладают вомероназальным органом , который участвует в обнаружении феромонов, включая половые феромоны . [20] Несмотря на то, что у людей нет этого органа, взрослые люди, по-видимому, чувствительны к определенным феромонам млекопитающих, которые способны обнаруживать предполагаемые белки рецепторов феромонов в обонятельном эпителии . [примечание 1] [20] Хотя половые феромоны явно играют роль в изменении сексуального поведения у некоторых млекопитающих, способность к общему обнаружению феромонов и участие феромонов в сексуальном поведении человека до сих пор не определены. [13]
Продолжительность копуляции значительно варьируется между видами млекопитающих [24] и может коррелировать с массой тела, продолжаясь дольше у крупных млекопитающих, чем у мелких млекопитающих. [25] Продолжительность копуляции также может коррелировать с длиной бакулума у млекопитающих [26]
Самцы млекопитающих извергают семя через пенис в половые пути самки во время совокупления. [27] [28] Эякуляция обычно происходит после одной интромиссии у людей, псовых и копытных , но происходит после нескольких интромиссий у большинства видов млекопитающих. [29] [30]
Копуляция может вызвать овуляцию у видов млекопитающих, которые не овулируют спонтанно. [31]Цикл сексуального удовольствия соответствует базовой структуре циклов удовольствия, связанных с другими вознаграждениями (такими как еда), и поэтому может быть также выражен в терминах мотивация–удовлетворение–насыщение или желание–симпатия–торможение (рисунок 1; вставка 2).6,11,1 ... Подобно другим формам обучения, сексуальное поведение развивается с течением времени, поскольку люди учатся связывать стимулы, такие как телесные особенности, личность и контекстуальные сигналы, с сексуальным удовольствием, вызванным гениталиями.7 Подростковый возраст, возможно, является наиболее критической фазой в сексуальном развитии ... Распространенное мнение гласит, что люди также реагируют на некоторые дистальные сексуальные стимулы (грудь, феромоны) безусловным образом, но это было трудно оценить эмпирически (вставка 1) ... Сексуальное желание как у крыс, так и у людей включает взаимодействие между половыми гормонами и внешними стимулами, которые становятся сексуальными стимулами через ассоциацию с Генитально-индуцированное сексуальное вознаграждение; приятная генитальная стимуляция, таким образом, является основным фактором сексуального обучения...
• Безусловные сексуальные стимулы (то есть те, для которых приятный эффект не требует обучения) включают проксимальную генитальную тактильную стимуляцию у людей и дистальные стимулы, такие как феромоны, запахи и определенные слуховые вокализации у крыс.7,16
• Сексуальное торможение включает в себя схожие механизмы мозга у крыс и людей
• Крысы демонстрируют схожую с людьми картину активации мозга в ответ на сигналы, связанные с сексуальным вознаграждением
• Кортикальные, лимбические, гипоталамические и мозжечковые области активируются стимулами, связанными с сексом, как у людей, так и у крыс
Сексуальное поведение следует за гормональным дисбалансом, по крайней мере у мужчин, но также в значительной степени основано на удовольствии. Получение и следование этим первичным пищевым и брачным вознаграждениям является основной причиной, по которой система вознаграждения мозга развилась в первую очередь. Обратите внимание, что "первичное" вознаграждение не относится к различию между безусловным и условным вознаграждением; на самом деле, большинство первичных вознаграждений усваиваются и, таким образом, обуславливаются (еда является первичным вознаграждением, которое обычно усваивается). ... Удовольствие - это не только одна из трех основных функций вознаграждения, но и дает определение вознаграждения. Поскольку гомеостаз объясняет функции лишь ограниченного числа вознаграждений, преобладающей причиной, по которой определенные стимулы, объекты, события, ситуации и действия являются вознаграждениями, может быть удовольствие. Это относится прежде всего к сексу (кто бы занимался нелепой гимнастикой репродуктивной активности, если бы не удовольствие) и к первичным гомеостатическим вознаграждениям в виде еды и жидкости, и распространяется на деньги, вкус, красоту, социальные встречи и нематериальные, внутренне заданные и внутренние вознаграждения. ... Желание делает поведение целенаправленным и направляет его к идентифицируемым целям. Таким образом, желание — это эмоция, которая помогает активно направлять поведение к известным вознаграждениям, тогда как удовольствие — это пассивный опыт, который вытекает из полученного или ожидаемого вознаграждения. Желание имеет множественные отношения с удовольствием; оно может быть приятным само по себе (я чувствую приятное желание), и оно может приводить к удовольствию (я желаю получить приятный объект). Таким образом, удовольствие и желание имеют отличительные характеристики, но тесно переплетены. Они представляют собой самые важные положительные эмоции, вызываемые вознаграждениями. Они отдают приоритет нашей сознательной обработке и, таким образом, составляют важные компоненты поведенческого контроля. Эти эмоции также называются симпатией (для удовольствия) и желанием (для желания) в исследованиях зависимости (471) и решительно поддерживают функции вознаграждения, связанные с обучением и подходом. ... Некоторые из стимулов и событий, которые доставляют удовольствие людям, могут даже не вызывать удовольствия у животных, а действовать через врожденные механизмы. Мы просто не знаем. Тем не менее, изобретение удовольствия и желания эволюцией имело огромное преимущество, позволяя большому количеству стимулов, объектов, событий, ситуаций и видов деятельности быть привлекательными. Этот механизм в значительной степени поддерживает основные функции вознаграждения при получении необходимых веществ и партнеров для спаривания.
Гены OR преимущественно экспрессируются в сенсорных нейронах основного обонятельного эпителия (MOE) в носовой полости. Млекопитающие обнаруживают многие типы химических веществ в воздухе и некоторые в воде как одоранты, тогда как рыбы распознают водорастворимые молекулы, такие как аминокислоты, желчные кислоты, половые стероиды и простагландины. Известно, что некоторые гены OR млекопитающих экспрессируются в других тканях, включая яички, язык, мозг и плаценту17. Однако функциональное значение такой «эктопической экспрессии» генов OR окончательно не известно. TAAR также экспрессируются в MOE. Эти рецепторы были впервые идентифицированы как мозговые рецепторы для следовых аминов, набора аминов, которые присутствуют в низких концентрациях в центральной нервной системе18. Первоначально предполагалось, что TAAR участвуют в психиатрических расстройствах19, но теперь известно, что они функционируют как второй класс обонятельных рецепторов10. Некоторые мышиные TAAR распознают летучие амины, которые присутствуют в моче, и, по-видимому, функция TAAR заключается в обнаружении лигандов, связанных с социальными сигналами10. ... Большинство млекопитающих обладают дополнительным обонятельным органом, называемым вомероназальным органом (VnO). ... Ранее считалось, что VnO является специализированным органом для обнаружения феромонов, но теперь известно, что VnO и MOE имеют некоторые перекрывающиеся функции22. ... Однако, по крайней мере, один из пяти генов V1R экспрессируется в обонятельной слизистой оболочке человека72. Кроме того, недавнее исследование предполагает, что эти пять генов могут активировать OR-подобный путь передачи сигнала в гетерологичной системе экспрессии73. Поэтому возможно, что продукты этих генов функционируют как феромонные или обонятельные рецепторы. У взрослых людей нет VnO, но они, по-видимому, чувствительны к феромонам74. Еще одно интересное наблюдение заключается в том, что у кур нет функциональных или нефункциональных генов V1R и V2R или VnO75, хотя птицы используют феромоны для выбора партнера и других форм поведения76. Возможно, что некоторые гены OR в MOE способны обнаруживать феромоны, как у людей74,77.
другого рецептора подтверждаются TAAR5-независимой триметиламиновой аносмией у людей [32]. ... Несколько TAAR обнаруживают летучие и аверсивные амины, но обонятельная система способна отбрасывать ограничения на основе лигандов или функций в эволюции TAAR. Определенные TAAR мутировали, чтобы распознавать новые лиганды, при этом почти вся клада костистых рыб потеряла канонический мотив распознавания аминов. Более того, хотя некоторые TAAR обнаруживают неприятные запахи, поведение, опосредованное TAAR, может различаться у разных видов. ... Способность отдельных TAAR опосредовать поведение отвращения и влечения предоставляет захватывающую возможность для механистического раскрытия кодирования валентности запаха.
Важно отметить, что три лиганда, идентифицированных для активации мышиных Taar, являются естественными компонентами мышиной мочи, основного источника социальных сигналов у грызунов. Мышиный Taar4 распознает β-фенилэтиламин, соединение, повышение уровня которого в моче коррелирует с увеличением стресса и стрессовых реакций как у грызунов, так и у людей. И мышиные Taar3, и Taar5 обнаруживают соединения (изоамиламин и триметиламин соответственно), которые обогащены в моче самцов по сравнению с самками мышей. Сообщается, что изоамиламин в мужской моче действует как феромон, ускоряя начало полового созревания у самок мышей [34]. Авторы предполагают, что семейство Taar имеет хемосенсорную функцию, которая отличается от обонятельных рецепторов с ролью, связанной с обнаружением социальных сигналов. ... Эволюционная модель семейства генов TAAR характеризуется специфической для линий филогенетической кластеризацией [26,30,35]. Эти характеристики очень похожи на те, которые наблюдаются в обонятельных GPCR и вомероназальных (V1R, V2R) семействах генов GPCR.
В то время как мыши производят специфичные для пола количества мочевого ТМА и привлекаются ТМА, этот запах отталкивает крыс и вызывает отвращение у людей [19], что указывает на то, что должны быть видоспецифичные функции. ... Более того, гомозиготный нокаут мышиного TAAR5 отменил поведение влечения к TMA [19]. Таким образом, делается вывод, что сам по себе TAAR5 достаточен для опосредования поведенческого ответа, по крайней мере, у мышей. ... Вызывает ли активация TAAR5 TMA специфический поведенческий выход, такой как поведение избегания у людей, все еще необходимо изучить.
_(18195445165).jpg/1280px-The_anatomy_of_the_domestic_animals_(1914)_(18195445165).jpg)
_medicine_(1890)_(20111730104).jpg/1280px-A_text-book_of_comparative_physiology_for_students_and_practitioners_of_comparative_(veterinary)_medicine_(1890)_(20111730104).jpg)
_(17571533394).jpg/1280px-Anatomy_of_the_cat_(1991)_(17571533394).jpg)
_(17571535364).jpg/1280px-Anatomy_of_the_cat_(1991)_(17571535364).jpg)
_(19805173074).jpg/1280px-Bulletin_(1973)_(19805173074).jpg)
_(20241179329).jpg/1280px-Image_from_page_91_of_%22Bulletin%22_(1918)_(20241179329).jpg)
_(18006271698).jpg/1280px-Anatomischer_Anzeiger_(1922)_(18006271698).jpg)
_(16042152204).jpg/1280px-Image_from_page_101_of_%22Practical_anatomy_of_the_rabbit_-_an_elementary_laboratory_textbook_in_mammalian_anatomy%22_(1921)_(16042152204).jpg)