Гребневики ( / t ə ˈ n ɒ f ər ə / tə- NOF -ər-ə ; ед. ч .: гребневик / ˈ t ɛ n ə f ɔːr , ˈ t iː n ə -/ TEN -ə-for, TEE -nə- ; от древнегреческого κτείς (kteis) «гребень» и φέρω (pherō) «нести») [6] составляют тип морских беспозвоночных , обычно известных как гребневиковые медузы , которые населяют морские воды по всему миру. Они примечательны группами ресничек , которые они используют для плавания (обычно называемых «гребнями»), и они являются самыми крупными животными, плавающими с помощью ресничек.
В зависимости от вида, взрослые гребневики имеют размеры от нескольких миллиметров до 1,5 м (5 футов). В настоящее время известно всего 186 живых видов. [7]
Их тела состоят из массы желе, со слоем толщиной в две клетки снаружи и еще одним, выстилающим внутреннюю полость. Тип имеет широкий спектр форм тела, включая яйцевидных цидиппидов с парой втягивающихся щупалец, которые захватывают добычу, плоских, как правило, без гребней платиктенид и большеротых бероидов , которые охотятся на других гребневиков.
Почти все гребневики ведут хищный образ жизни , питаясь различной добычей — от микроскопических личинок и коловраток до взрослых особей мелких ракообразных ; исключение составляют молодь двух видов, которые паразитируют на сальпах , которыми питаются взрослые особи их вида.
Несмотря на свои мягкие, студенистые тела, окаменелости, которые, как полагают, представляют собой гребневиков, появляются в лагерштеттенах, датируемых еще ранним кембрием , около 525 миллионов лет назад. Положение гребневиков на «древе жизни» долгое время обсуждалось в исследованиях молекулярной филогенетики. Биологи предположили, что гребневики составляют вторую по раннему разветвлению линию животных, а губки являются сестринской группой для всех других многоклеточных животных ( гипотеза сестринства Porifera ). [8] Другие биологи утверждают, что гребневики появились раньше губок ( гипотеза сестринства Ctenophora ), которые сами появились до разделения на книдарий и билатерий . [9] [10] Пизани и др . повторно проанализировали данные и предположили, что компьютерные алгоритмы, используемые для анализа, были введены в заблуждение наличием специфических генов гребневиков, которые заметно отличались от генов других видов. [11] [12] [ нужна страница ] Последующий анализ Уилана и др. (2017) [13] дал дополнительную поддержку гипотезе «сестры гребневика»; этот вопрос остается предметом таксономического спора. [14] [15] Шульц и др. (2023) обнаружили необратимые изменения в синтении у сестры гребневика, Myriazoa , состоящей из остальных животных. [16]

Среди типов животных гребневики сложнее губок , примерно такие же сложные, как книдарии ( медузы , актинии и т. д.), и менее сложные, чем билатерии (которые включают почти всех остальных животных). В отличие от губок, и гребневики, и книдарии имеют:
Гребневики отличаются от всех других животных наличием коллобластов , которые липкие и прилипают к добыче, хотя у некоторых видов гребневиков они отсутствуют. [18] [19]
Как и книдарии, гребневики имеют два основных слоя клеток, которые зажаты между средним слоем желеобразного материала, который называется мезоглеей у книдарий и гребневиков; более сложные животные имеют три основных слоя клеток и не имеют промежуточного желеобразного слоя. Поэтому гребневики и книдарии традиционно называются диплобластическими . [18] [20] И гребневики, и книдарии имеют тип мышц , который у более сложных животных возникает из среднего слоя клеток , [21] и в результате некоторые недавние учебники классифицируют гребневиков как триплобластических , [22] в то время как другие по-прежнему считают их диплобластическими. [18] Гребневики имеют более 80 различных типов клеток , что превышает количество у других групп, таких как плакозои, губки, книдарии и некоторые глубоко ветвящиеся билатерии. [23]
Размером от 1 миллиметра (0,04 дюйма) до 1,5 метра (5 футов) [22] [24] гребневики являются крупнейшими неколониальными животными, которые используют реснички («волоски») в качестве основного способа передвижения. [22] У большинства видов есть восемь полос, называемых рядами гребней, которые проходят по всей длине их тела и несут гребнеобразные полосы ресничек, называемые «ктенами», расположенные вдоль рядов гребней так, что когда реснички бьют, реснички каждого гребня касаются гребня ниже. [22] Название «ctenophora» означает «несущий гребень», от греческого κτείς (основная форма κτεν- ), что означает «гребень», и греческого суффикса -φορος, что означает «несущий». [25]
Для типа с относительно небольшим количеством видов гребневики имеют широкий спектр планов тела. [22] Прибрежные виды должны быть достаточно прочными, чтобы выдерживать волны и завихряющиеся частицы осадка, в то время как некоторые океанические виды настолько хрупкие, что их очень трудно поймать в целости и сохранности для изучения. [19] Кроме того, океанические виды плохо сохраняются, [19] и известны в основном по фотографиям и заметкам наблюдателей. [30] Поэтому до недавнего времени основное внимание было сосредоточено на трех прибрежных родах – Pleurobrachia , Beroe и Mnemiopsis . [19] [31] По крайней мере два учебника основывают свои описания гребневиков на цидиппиде Pleurobrachia . [18] [22]
Поскольку тело многих видов почти радиально симметрично , главная ось направлена от орального к аборальному (от рта к противоположному концу). Однако, поскольку только два канала около статоциста заканчиваются анальными порами, у гребневиков нет зеркальной симметрии, хотя у многих есть вращательная симметрия. Другими словами, если животное вращается по полукругу, оно выглядит так же, как и в начале. [32]
Тип гребневиков имеет широкий спектр форм тела, включая уплощенных глубоководных платиктенид , у взрослых особей большинства видов которых отсутствуют гребни, и прибрежных бероидов , у которых отсутствуют щупальца и которые охотятся на других гребневиков с помощью огромных ртов, вооруженных группами крупных жестких ресничек , которые выполняют функцию зубов.

Как и у книдарий ( медуз , актиний и т. д.), тела гребневиков состоят из относительно толстой, желеобразной мезоглеи, зажатой между двумя эпителиями , слоями клеток, связанных межклеточными соединениями и фиброзной базальной мембраной , которую они секретируют . [18] [22] Эпителий гребневиков состоит из двух слоев клеток, а не из одного, и некоторые клетки в верхнем слое имеют несколько ресничек на клетку. [22]
Внешний слой эпидермиса ( наружная кожа) состоит из: сенсорных клеток; клеток, которые выделяют слизь , которая защищает тело; и интерстициальных клеток, которые могут трансформироваться в другие типы клеток. В специализированных частях тела внешний слой также содержит коллобласты , которые находятся вдоль поверхности щупалец и используются для захвата добычи, или клетки, несущие несколько больших ресничек, для передвижения. Внутренний слой эпидермиса содержит нервную сеть и миоэпителиальные клетки, которые действуют как мышцы . [22]
Внутренняя полость образует: рот, который обычно может быть закрыт мышцами; глотку («горло»); более широкую область в центре, которая действует как желудок ; и систему внутренних каналов. Они разветвляются через мезоглею к наиболее активным частям животного: рот и глотка; корни щупалец, если они есть; по всей нижней стороне каждого ряда гребней; и четыре ветви вокруг сенсорного комплекса на дальнем конце от рта — две из этих четырех ветвей заканчиваются анальными порами. Внутренняя поверхность полости выстлана эпителием , гастродермисом . Рот и глотка имеют как реснички , так и хорошо развитые мышцы. В других частях системы каналов гастродермис отличается на сторонах, ближайших и самых дальних от органа, который он снабжает. Ближайшая сторона состоит из высоких питательных клеток, которые хранят питательные вещества в вакуолях (внутренних отсеках), зародышевых клеток , которые производят яйцеклетки или сперму, и фотоцитов , которые производят биолюминесценцию . Сторона, наиболее удаленная от органа, покрыта ресничными клетками, которые циркулируют воду через каналы, перемежающиеся ресничными розетками, порами, которые окружены двойными завитками ресничек и соединяются с мезоглеей. [22]
Когда добыча проглатывается, она разжижается в глотке ферментами и мышечными сокращениями глотки. Полученная кашица проносится через систему каналов биением ресничек и переваривается питательными клетками. Реснички в каналах могут помогать транспортировать питательные вещества к мышцам мезоглеи. Анальные поры могут выбрасывать нежелательные мелкие частицы, но большая часть нежелательных веществ отрыгивается через рот. [22]
Мало что известно о том, как гребневики избавляются от отходов, производимых клетками. Ресничные розетки в гастродермисе могут помогать удалять отходы из мезоглеи, а также могут помогать регулировать плавучесть животного , закачивая воду в мезоглею или из нее. [22]
На внешней поверхности обычно расположено восемь рядов гребней, называемых плавательными пластинами, которые используются для плавания. Ряды ориентированы так, чтобы идти от рта («оральный полюс») к противоположному концу («аборальный полюс»), и расположены более или менее равномерно по всему телу, [18] хотя схемы расположения различаются в зависимости от вида, и у большинства видов ряды гребней простираются только на часть расстояния от аборального полюса по направлению к рту. «Гребни» (также называемые «ктенами» или «гребневыми пластинами») проходят через каждый ряд, и каждый состоит из тысяч необычно длинных ресничек, до 2 миллиметров (0,08 дюйма). В отличие от обычных ресничек и жгутиков, которые имеют структуру нитей, расположенных по схеме 9 + 2, эти реснички расположены по схеме 9 + 3, где предполагается, что дополнительная компактная нить выполняет опорную функцию. [33] Они обычно бьют так, чтобы движущий удар был направлен в сторону от рта, хотя они также могут менять направление. Поэтому гребневики обычно плывут в том направлении, в котором рот совершает прием пищи, в отличие от медуз . [22] Пытаясь спастись от хищников, один вид может ускориться до шестикратной скорости; [34] некоторые другие виды меняют направление в рамках своего поведения побега, изменяя направление рабочего хода ресничек гребневой пластины.
Неясно, как гребневики контролируют свою плавучесть, но эксперименты показали, что некоторые виды полагаются на осмотическое давление, чтобы адаптироваться к воде разной плотности. [35] Их жидкости в организме обычно такие же концентрированные , как морская вода. Если они попадают в менее плотную солоноватую воду, ресничные розетки в полости тела могут перекачивать ее в мезоглею , чтобы увеличить ее объем и уменьшить ее плотность, чтобы избежать затопления. И наоборот, если они перемещаются из солоноватой в полную морскую воду, розетки могут выкачивать воду из мезоглеи, чтобы уменьшить ее объем и увеличить ее плотность. [22]
У гребневиков нет мозга или центральной нервной системы , но вместо этого есть субэпидермальная нервная сеть (скорее похожая на паутину), которая образует кольцо вокруг рта и является наиболее плотной вблизи таких структур, как гребневые ряды, глотка, щупальца (если они есть) и сенсорный комплекс, наиболее удаленный от рта. [22] Связь между нервными клетками осуществляется двумя различными способами; обнаружено, что некоторые нейроны имеют синаптические связи , но нейроны в нервной сети весьма различимы, поскольку они слиты в синцитий , а не связаны синапсами. Некоторые животные за пределами гребневиков также имеют слитые нервные клетки, но никогда не в такой степени, чтобы они образовывали целую нервную сеть. [36] [37] [38] Ископаемые остатки показывают, что у кембрийских видов была более сложная нервная система с длинными нервами, которые соединялись с кольцом вокруг рта. Единственный известный на сегодняшний день гребневик с длинными нервами — это Euplokamis из отряда Cydippida. [39] Их нервные клетки возникают из тех же клеток-предшественников, что и коллобласты. [40]
Кроме того, существует менее организованная мезоглеальная нервная сеть, состоящая из отдельных невритов. Самая большая отдельная сенсорная особенность - аборальный орган (на противоположном конце от рта), который подчеркивается собственной нервной сетью. [41] Основным компонентом этого органа является статоцист , датчик равновесия, состоящий из статолита, крошечного зерна карбоната кальция, поддерживаемого четырьмя пучками ресничек , называемых «балансирами», которые чувствуют его ориентацию. Статоцист защищен прозрачным куполом, сделанным из длинных неподвижных ресничек. Гребневик не пытается автоматически удерживать статолит одинаково лежащим на всех балансирах. Вместо этого его реакция определяется «настроением» животного, другими словами, общим состоянием нервной системы. Например, если гребневик с волочащимися щупальцами захватывает добычу, он часто разворачивает некоторые ряды гребней в обратном направлении, вращая рот в сторону добычи. [22]
Исследования подтверждают гипотезу о том, что ресничные личинки книдарий и билатерий имеют древнее и общее происхождение. [42] Апикальный орган личинок участвует в формировании нервной системы. [43] Аборальный орган гребневиков не гомологичен апикальному органу других животных, и поэтому формирование их нервной системы имеет иное эмбриональное происхождение. [44]
Нервные клетки и нервная система гребневиков имеют иную биохимию по сравнению с другими животными. Например, у них отсутствуют гены и ферменты, необходимые для производства нейротрансмиттеров, таких как серотонин , дофамин , оксид азота , октопамин , норадреналин и другие, которые в противном случае наблюдаются у всех других животных с нервной системой, при этом гены, кодирующие рецепторы для каждого из этих нейротрансмиттеров, отсутствуют. [45] Монофункциональная каталаза (CAT), одно из трех основных семейств антиоксидантных ферментов, которые нацелены на перекись водорода , важную сигнальную молекулу для синаптической и нейронной активности, также отсутствует, скорее всего, из-за потери гена. [46] Было обнаружено, что они используют L-глутамат в качестве нейротрансмиттера и имеют необычно большое разнообразие ионотропных рецепторов глутамата и генов для синтеза и транспорта глутамата по сравнению с другими метазоа. [47] Геномное содержание генов нервной системы является наименьшим из известных среди всех животных и может представлять собой минимальные генетические требования для функциональной нервной системы. [48] Тот факт, что части нервной системы имеют непосредственно слитые нейроны без синапсов, предполагает, что гребневики могут образовывать сестринскую группу для других метазоа, развив нервную систему независимо. [38] Если гребневики являются сестринской группой для всех других метазоа, нервные системы могли быть либо утрачены у губок и плакозоа, либо возникли более одного раза среди метазоа. [49]

Гребневики Cydippid имеют тела, которые более или менее округлые, иногда почти сферические, а иногда более цилиндрические или яйцевидные; обычный прибрежный «морской крыжовник», Pleurobrachia , иногда имеет яйцевидное тело со ртом на узком конце, [22] хотя некоторые особи более равномерно круглые. С противоположных сторон тела тянется пара длинных, тонких щупалец, каждое из которых заключено в оболочку, в которую оно может быть втянуто. [18] Некоторые виды cydippid имеют тела, которые в разной степени сплющены, так что они шире в плоскости щупалец. [22]
Щупальца гребневиков цидиппид обычно окаймлены тентиллами («маленькими щупальцами»), хотя у нескольких родов есть простые щупальца без этих боковых ответвлений. Щупальца и тентиллы густо покрыты микроскопическими коллобластами , которые захватывают добычу, прилипая к ней. Коллобласты — это специализированные грибовидные клетки во внешнем слое эпидермиса, имеющие три основных компонента: куполообразную головку с пузырьками (камерами), содержащими клей; стебель, который закрепляет клетку в нижнем слое эпидермиса или в мезоглее; и спиральную нить, которая обвивается вокруг стебля и прикрепляется к головке и к корню стебля. Функция спиральной нити не определена, но она может поглощать стресс, когда добыча пытается сбежать, и таким образом предотвращать разрыв коллобласта. [22] Один вид, Minictena luteola, диаметр которого составляет всего 1,5 мм, имеет пять различных типов клеток коллобласта. [50] [51]
В дополнение к коллобластам, представители рода Haeckelia , которые питаются в основном медузами , включают жалящие нематоциты своих жертв в свои собственные щупальца — некоторые питающиеся книдариями голожаберники аналогичным образом включают нематоциты в свои тела для защиты. [52] Тентилла Euplokamis значительно отличается от таковых у других цидиппидов: они содержат поперечнополосатую мускулатуру , тип клеток, неизвестный в типе Ctenophora; и они сворачиваются в расслабленном состоянии, в то время как тентилла всех других известных гребневиков удлиняется в расслабленном состоянии. Тентилла Euplokamis имеет три типа движения, которые используются для захвата добычи: они могут очень быстро выбрасываться (за 40–60 миллисекунд ); они могут извиваться, что может заманивать добычу, ведя себя как мелкие планктонные черви; и они обвиваются вокруг добычи. Уникальное дергание — это раскручивающееся движение, приводимое в действие сокращением поперечно -полосатой мышцы . Извивающееся движение производится гладкими мышцами , но весьма специализированного типа. Обвивание вокруг добычи осуществляется в основном за счет возвращения тентиллы в неактивное состояние, но спирали могут быть стянуты гладкими мышцами. [53]
Существует восемь рядов гребней, которые идут от рта до противоположного конца и равномерно распределены по всему телу. [18] «Гребни» бьют в метахрональном ритме, похожем на ритм мексиканской волны . [54] От каждого балансира в статоцисте ресничный желобок идет под куполом и затем разделяется, чтобы соединиться с двумя соседними рядами гребней, а у некоторых видов проходит вдоль рядов гребней. Это образует механическую систему для передачи ритма ударов от гребней к балансирам посредством водных возмущений, создаваемых ресничками. [55]
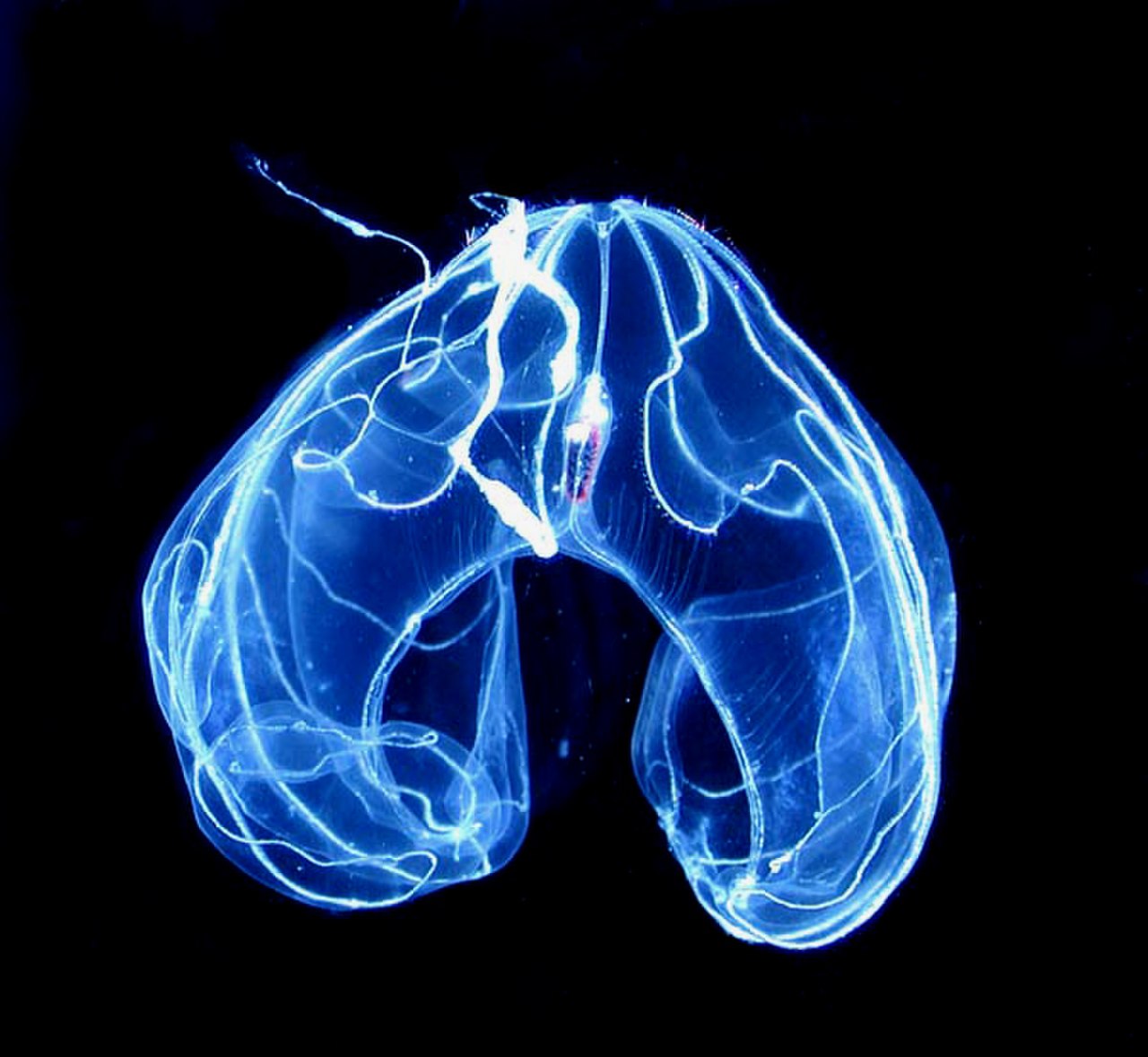
У Lobata есть пара лопастей, которые представляют собой мускулистые, чашеобразные расширения тела, выступающие за пределы рта. Их незаметные щупальца берут начало из углов рта, проходя в извилистых бороздках и распространяясь по внутренней поверхности лопастей (а не волочась далеко позади, как у Cydippida). Между лопастями по обе стороны рта у многих видов лопастных есть четыре ушка, студенистые выступы, окаймленные ресничками, которые создают водные потоки, помогающие направлять микроскопическую добычу ко рту. Такое сочетание структур позволяет лопастным непрерывно питаться взвешенной планктонной добычей. [22]
У лопастных есть восемь рядов гребней, начинающихся на аборальном полюсе и обычно не выходящих за пределы тела к лопастям; у видов с (четырьмя) ушными раковинами реснички, окаймляющие ушные раковины, являются продолжениями ресничек в четырех рядах гребней. Большинство лопастных довольно пассивны при движении в воде, используя реснички на своих рядах гребней для движения вперед, [22] хотя у Leucothea длинные и активные ушные раковины, движения которых также способствуют движению вперед. Представители родов лопастных Bathocyroe и Ocyropsis могут спасаться от опасности, хлопая лопастями, так что струя выбрасываемой воды очень быстро отбрасывает их назад. [56] В отличие от цидиппидов, движения гребней лопастных ракообразных координируются нервами, а не водными возмущениями, создаваемыми ресничками, однако гребни в одном ряду бьются в том же мексиканском волновом стиле, что и механически координируемые ряды гребней цидиппидов и бероидов. [55] Это могло позволить лопастным ракообразным вырасти больше, чем цидиппиды, и иметь менее яйцевидную форму. [54]
Необычный вид, впервые описанный в 2000 году, Lobatolampea tetragona , был классифицирован как лопастной, хотя лопасти являются «примитивными», а тело напоминает медузу , когда плавает, и похоже на диск, когда отдыхает на морском дне. [30]

Beroida , также известные как Nuda , не имеют придатков для кормления, но их большая глотка , расположенная внутри большого рта и заполняющая большую часть мешковидного тела, несет «макроцилии» на оральном конце. Эти сросшиеся пучки из нескольких тысяч крупных ресничек способны «откусывать» куски добычи, которые слишком велики, чтобы проглотить их целиком – почти всегда это другие гребневики. [ 57] Перед полем макроцилий, на «губах» рта у некоторых видов Beroe , находится пара узких полосок адгезивных эпителиальных клеток на стенке желудка, которые «застегивают» рот, когда животное не питается, образуя межклеточные соединения с противоположной адгезивной полосой. Это плотное закрытие обтекает переднюю часть животного, когда оно преследует добычу. [58]
У Ганешиды есть пара маленьких ротовых лопастей и пара щупалец. Тело имеет скорее круглую, чем овальную форму в поперечном сечении, а глотка простирается над внутренними поверхностями долей. [22]
Thalassocalycida , обнаруженные только в 1978 году и известные только по одному виду, [ 59] похожи на медуз, с телами, которые укорочены в орально-аборальном направлении, и короткими гребнями на поверхности, наиболее удаленной от рта, берущими начало вблизи аборального полюса. Они захватывают добычу движениями колокола и, возможно, с помощью двух коротких щупалец. [22]
Cestida («поясные животные») — лентовидные планктонные животные, у которых рот и аборальный орган выровнены посередине противоположных краев ленты. Вдоль каждого аборального края есть пара гребневых рядов, а также тентилла, выходящая из канавки вдоль всего орального края, которая струится назад по большей части крыловидной поверхности тела. Cestida могут плавать, совершая волнообразные движения своим телом, а также ударяя своими гребневыми рядами. Существует два известных вида, которые распространены по всему миру в теплых и умеренно-теплых водах: Cestum veneris (« пояс Венеры ») является одним из крупнейших гребневиков — до 1,5 метров (4,9 фута) в длину и может совершать волнообразные движения медленно или довольно быстро. Velamen parallelum , длина которого обычно составляет менее 20 сантиметров (0,66 фута), может двигаться гораздо быстрее, что было описано как «стремительное движение». [22] [60]
Большинство Platyctenida имеют овальные тела, сплющенные в орально-аборальном направлении, с парой щупалец с тентиллами на аборальной поверхности. Они цепляются за поверхности и ползают по ним, выворачивая глотку и используя ее как мускулистую «ногу». Все, кроме одного из известных видов Platyctenida, не имеют гребневых рядов. [22] Platyctenida обычно имеют криптическую окраску, живут на камнях, водорослях или поверхностях тел других беспозвоночных и часто обнаруживаются по своим длинным щупальцам со множеством боковых ответвлений, которые видны струящимися от спины гребневика в течение.

Взрослые особи большинства видов могут регенерировать ткани, которые были повреждены или удалены, [61] хотя только платиктениды размножаются путем клонирования , отщепляя от краев своих плоских тел фрагменты, которые развиваются в новых особей. [22] Лабораторные исследования Mnemiopsis leidyi также показывают, что когда двум особям удаляют части их тел, они способны сливаться вместе, включая их нервную и пищеварительную системы, даже если две особи генетически различны. Феномен, который до сих пор был обнаружен только у гребневиков. [62]
Последний общий предок (LCA) гребневиков был гермафродитом . [63] Некоторые из них являются одновременными гермафродитами, которые могут производить как яйца, так и сперму одновременно, в то время как другие являются последовательными гермафродитами, у которых яйца и сперма созревают в разное время. Метаморфоза нет . [64] Известно, что по крайней мере три вида развили раздельные полы ( двуполость); Ocyropsis crystallina и Ocyropsis maculata в роде Ocyropsis и Bathocyroe fosteri в роде Bathocyroe. [65] Гонады расположены в частях внутренней сети каналов под рядами гребней , а яйца и сперма высвобождаются через поры в эпидермисе. Оплодотворение, как правило, внешнее , но платиктениды используют внутреннее оплодотворение и держат яйца в выводковых камерах до тех пор, пока они не вылупятся. Самооплодотворение иногда наблюдалось у видов рода Mnemiopsis [22] , и считается, что большинство гермафродитных видов являются самооплодотворяющими. [19]
Развитие оплодотворенных яиц прямое; отличительной личиночной формы нет. Молодь всех групп, как правило, планктонная , и большинство видов напоминают миниатюрных взрослых цидиппид, постепенно развивая свои взрослые формы тела по мере роста. Однако в роде Beroe у молоди большие рты и, как и у взрослых, отсутствуют как щупальца, так и оболочки щупалец. В некоторых группах, таких как плоские, обитающие на дне платиктениды, молодь ведет себя больше как настоящие личинки. Они живут среди планктона и, таким образом, занимают иную экологическую нишу, чем их родители, достигая взрослой формы только путем более радикального онтогенеза [22] после падения на морское дно. [19]
По крайней мере, у некоторых видов молодые гребневики, по-видимому, способны производить небольшое количество яйцеклеток и спермы, пока они значительно ниже взрослых размеров, а взрослые производят яйцеклетки и сперму до тех пор, пока у них достаточно пищи. Если у них заканчивается еда, они сначала прекращают производить яйцеклетки и сперму, а затем уменьшаются в размерах. Когда поступление пищи улучшается, они вырастают до нормального размера, а затем возобновляют размножение. Эти особенности делают гребневиков способными очень быстро увеличивать свою популяцию. [19] Представители Lobata и Cydippida также имеют форму размножения, называемую диссогенией; две половозрелые стадии, сначала как личинка, а затем как молодь и взрослые особи. Во время своего пребывания в качестве личинки они способны периодически выпускать гаметы. После того, как их первый репродуктивный период заканчивается, они не будут производить больше гамет снова до более позднего времени. Популяция Mertensia ovum в центральной части Балтийского моря стала педогенетической и состоит исключительно из половозрелых личинок размером менее 1,6 мм. [66] [67]
У Mnemiopsis leidyi сигнализация оксида азота (NO) присутствует как во взрослых тканях, так и дифференциально выражена на более поздних эмбриональных стадиях, что предполагает участие NO в механизмах развития. [68] Зрелая форма того же вида также способна возвращаться к стадии цидиппида при воздействии стрессовых факторов окружающей среды. [69]

Большинство гребневиков, которые живут вблизи поверхности, в основном бесцветны и почти прозрачны. Однако некоторые виды, живущие на большей глубине, сильно пигментированы, например, вид, известный как «Tortugas red» [70] (см. иллюстрацию здесь), который еще не был официально описан. [19] Платиктениды обычно живут, прикрепленные к другим организмам морского дна, и часто имеют схожую окраску с этими организмами-хозяевами. [19] Кишечник глубоководного рода Bathocyroe красный, что скрывает биолюминесценцию веслоногих рачков, которых он проглотил. [56]
Ряды гребней большинства планктонных гребневиков создают радужный эффект, который вызван не биолюминесценцией , а рассеиванием света при движении гребней. [19] [71] Большинство видов также биолюминесцентны, но свет обычно синий или зеленый и виден только в темноте. [19] Однако некоторые значительные группы, включая всех известных платиктенид и род цидиппид Pleurobrachia , не способны к биолюминесценции. [72]
Когда некоторые виды, включая Bathyctena chuni , Euplokamis stationis и Eurhamphaea vexilligera , потревожены, они производят секрецию (чернила), которые люминесцируют на тех же длинах волн , что и их тела. Молодь будет люминесцировать ярче по отношению к размеру своего тела, чем взрослые, чье свечение рассеивается по всему телу. Подробное статистическое исследование не выявило функции биолюминесценции гребневиков и не выявило никакой корреляции между ее точным цветом и каким-либо аспектом среды обитания животных, таким как глубина или то, живут ли они в прибрежных или средних океанических водах. [73]
У гребневиков биолюминесценция вызвана активацией кальций-активируемых белков, называемых фотопротеинами, в клетках, называемых фотоцитами , которые часто ограничены меридиональными каналами, лежащими под восемью рядами гребней. В геноме Mnemiopsis leidyi десять генов кодируют фотопротеины. Эти гены коэкспрессируются с генами опсина в развивающихся фотоцитах Mnemiopsis leidyi , что повышает вероятность того, что производство света и обнаружение света могут работать вместе у этих животных. [74]

Гребневики встречаются в большинстве морских сред: от полярных вод при температуре −2 °C до тропиков при температуре 30 °C; вблизи побережий и в середине океана; от поверхностных вод до глубин океана на глубине более 7000 метров. [75] Наиболее изученными являются роды Pleurobrachia , Beroe и Mnemiopsis , поскольку эти планктонные прибрежные формы чаще всего встречаются вблизи берега. [31] [56] В пресной воде гребневики не обнаружены.
В 2013 году Mnemiopsis был зарегистрирован в озере Биркет-Карун, а в 2014 году — в озере Эль-Райян II, оба около Файюма в Египте, куда они были случайно завезены при транспортировке мальков рыбы (кефали). Хотя многие виды предпочитают солоноватые воды, такие как эстуарии и прибрежные лагуны, имеющие открытое сообщение с морем, это была первая запись из внутренней среды. Оба озера соленые, причем Биркет-Карун является гиперсоленым, и показывает, что некоторые гребневики могут обосноваться в соленых лимнических средах без связи с океаном. В долгосрочной перспективе не ожидается, что популяции выживут. Двумя ограничивающими факторами в соленых озерах являются доступность пищи и разнообразный рацион, а также высокие температуры в жаркое лето. Поскольку паразитический изопод Livoneca redmanii был завезен в то же время, трудно сказать, какая часть экологического воздействия инвазивных видов вызвана только гребневиком. [76] [77]
Гребневики могут быть многочисленны в летние месяцы в некоторых прибрежных районах, но в других местах они редки и их трудно найти.
В заливах, где они встречаются в очень больших количествах, хищничество гребневиков может контролировать популяции мелких зоопланктонных организмов, таких как веслоногие рачки , которые в противном случае могли бы уничтожить фитопланктон (планктонные растения), являющийся важной частью морских пищевых цепей .
Почти все гребневики являются хищниками — среди них нет вегетарианцев, и только один род является частично паразитическим . [56] Если пищи много, они могут съесть в 10 раз больше собственного веса в день. [78] В то время как Beroe охотится в основном на других гребневиков, другие виды, обитающие на поверхности воды, охотятся на зоопланктон (планктонных животных) размером от микроскопических, включая личинок моллюсков и рыб, до мелких взрослых ракообразных, таких как веслоногие рачки , амфиподы и даже криль . Представители рода Haeckelia охотятся на медуз и включают нематоцисты (стрекательные клетки) своей добычи в свои собственные щупальца вместо коллобластов . [19] Гребневиков сравнивали с пауками по их широкому спектру методов захвата добычи — некоторые неподвижно висят в воде, используя свои щупальца как «паутину», некоторые являются засадными хищниками, как пауки-скакуны Salticid , а некоторые подвешивают липкую каплю на конце тонкой нити, как пауки-болы . Это разнообразие объясняет широкий спектр форм тела в типе с довольно небольшим количеством видов. [56] Двухщупальцевый «цидиппид» Lampea питается исключительно сальпами , близкими родственниками асцидий, которые образуют большие цепочечные плавающие колонии, а молодые особи Lampea прикрепляются как паразиты к сальпам, которые слишком велики для того, чтобы они могли их проглотить. [56] Представители рода цидиппид Pleurobrachia и лопастного Bolinopsis часто достигают высокой плотности популяции в одном и том же месте и в одно и то же время, поскольку они специализируются на разных типах добычи: длинные щупальца Pleurobrachia в основном захватывают относительно сильных пловцов, таких как взрослые веслоногие рачки, в то время как Bolinopsis обычно питается более мелкими и слабыми пловцами, такими как коловратки , личинки моллюсков и ракообразных . [79]
Гребневики раньше считались «тупиками» в морских пищевых цепях, поскольку считалось, что их низкое соотношение органического вещества к соли и воде делало их плохой диетой для других животных. Также часто бывает трудно идентифицировать остатки гребневиков в кишечнике возможных хищников, хотя гребни иногда остаются нетронутыми достаточно долго, чтобы дать подсказку. Подробное исследование кеты , Oncorhynchus keta , показало, что эти рыбы переваривают гребневиков в 20 раз быстрее, чем такой же вес креветок , и что гребневики могут обеспечить хорошую диету, если их достаточно вокруг. Бероиды охотятся в основном на других гребневиков. Некоторые медузы и черепахи поедают большое количество гребневиков, и медузы могут временно уничтожить популяции гребневиков. Поскольку гребневики и медузы часто имеют большие сезонные колебания в популяции, большинство рыб, которые охотятся на них, являются универсалами и могут оказывать большее влияние на популяции, чем специализированные желееды. Это подчеркивается наблюдением за травоядными рыбами, намеренно питающимися студенистым зоопланктоном во время цветения в Красном море. [80] Личинки некоторых актиний являются паразитами на гребневиках, как и личинки некоторых плоских червей , которые паразитируют на рыбах, когда они достигают взрослой особи. [81]
Большинство видов являются гермафродитами , и детеныши по крайней мере некоторых видов способны к размножению до достижения размеров и формы взрослой особи. Такое сочетание гермафродитизма и раннего размножения позволяет небольшим популяциям расти с взрывной скоростью.

Гребневики могут балансировать морские экосистемы, не давая чрезмерному количеству веслоногих рачков поедать весь фитопланктон (планктонные растения) [82] , которые являются доминирующими морскими производителями органического вещества из неорганических ингредиентов. [83]
С другой стороны, в конце 1980-х годов западноатлантический гребневик Mnemiopsis leidyi был случайно занесен в Черное и Азовское моря через балластные цистерны судов и был обвинен в резком падении уловов рыбы, поедая как личинки рыб, так и мелких ракообразных, которые в противном случае служили бы пищей для взрослых рыб. [82] Мнемиопсис хорошо оснащен для вторжения на новые территории (хотя это не было предсказано до тех пор, пока он так успешно не колонизировал Черное море), поскольку он может очень быстро размножаться и выдерживать широкий диапазон температур и солености воды . [84] Воздействие было усилено хроническим переловом и эвтрофикацией , которая дала всей экосистеме кратковременный импульс, в результате чего популяция мнемиопсиса увеличилась даже быстрее, чем обычно [85] – и, прежде всего, отсутствием эффективных хищников для этих завезенных гребневиков. [84] Популяции мнемиопсиса в этих районах в конечном итоге были взяты под контроль случайным заносом североамериканского гребневика Beroe ovata , питающегося мнемиопсисом , [86] и похолоданием местного климата с 1991 по 1993 год, [85] что значительно замедлило метаболизм животного. [84] Однако обилие планктона в этом районе вряд ли восстановится до уровня, существовавшего до появления мнемиопсиса . [87]
В конце 1990-х годов Mnemiopsis появился в Каспийском море . Вскоре после этого появился Beroe ovata , который, как ожидается, сократит, но не устранит влияние Mnemiopsis в этом районе. Mnemiopsis также достиг восточного Средиземноморья в конце 1990-х годов и теперь, по-видимому, процветает в Северном и Балтийском морях . [19]
Число известных ныне живущих видов гребневиков неизвестно, поскольку многие из названных и формально описанных оказались идентичны видам, известным под другими научными названиями. Клаудия Миллс оценивает, что существует около 100–150 действительных видов, которые не являются дубликатами, и что по крайней мере еще 25, в основном глубоководные формы, были признаны отдельными, но еще не проанализированы достаточно подробно, чтобы поддержать формальное описание и наименование. [70]
Ранние авторы объединяли гребневиков с книдариями в один тип, называемый кишечнополостными, из-за морфологического сходства между двумя группами. Как и у книдарий, тела гребневиков состоят из массы желе, с одним слоем клеток снаружи и другим, выстилающим внутреннюю полость. Однако у гребневиков эти слои имеют глубину в две клетки, в то время как у книдарий они имеют глубину только в одну клетку. Гребневики также напоминают книдарий тем, что полагаются на поток воды через полость тела как для пищеварения, так и для дыхания, а также имеют децентрализованную нервную сеть , а не мозг. Геномные исследования показали, что нейроны гребневиков, которые во многом отличаются от нейронов других животных, эволюционировали независимо от нейронов других животных, [88] и растущее понимание различий между гребневиками и другими кишечнополостными убедило более поздних авторов классифицировать их как отдельные типы . Положение гребневиков в эволюционном генеалогическом древе животных уже давно является предметом споров, и в настоящее время большинство ученых придерживается точки зрения, основанной на данных молекулярной филогенетики , что книдарии и билатерии более тесно связаны друг с другом, чем каждый из них с гребневиками.

Традиционная классификация делит гребневиков на два класса : со щупальцами ( Tentaculata ) и без них ( Nuda ). Nuda содержит только один отряд ( Beroida ) и семейство ( Beroidae ), а также два рода : Beroe (несколько видов) и Neis (один вид). [70]
Tentaculata делятся на следующие восемь отрядов : [70 ]
Существуют ископаемые роды, считающиеся стволовой группой .
Несмотря на их хрупкие, студенистые тела, окаменелости , которые , как полагают, представляют собой гребневиков – по-видимому, без щупалец, но с гораздо большим количеством гребневых рядов, чем у современных форм – были найдены в лагерштеттенах еще в раннем кембрии , около 515 миллионов лет назад . Тем не менее, недавний анализ молекулярной филогенетики приходит к выводу, что общий предок возник приблизительно 350 миллионов лет назад ± 88 миллионов лет назад, что противоречит предыдущим оценкам, которые предполагают, что это произошло 66 миллионов лет назад после мел-палеогенового вымирания . [89]
Из-за своих мягких, студенистых тел гребневики чрезвычайно редки в качестве окаменелостей, а окаменелости, которые были интерпретированы как гребневики, были найдены только в лагерштеттенах , местах, где окружающая среда была исключительно подходящей для сохранения мягких тканей. До середины 1990-х годов было известно только два образца, достаточно хороших для анализа, оба из группы короны, из раннего девона (эмсского) периода . Затем три дополнительных предполагаемых вида были найдены в сланцах Берджесс и других канадских породах аналогичного возраста, около 505 миллионов лет назад в середине кембрийского периода. У всех трех не было щупалец, но было от 24 до 80 рядов гребней, что намного больше, чем восемь, типичных для современных видов. У них также, по-видимому, были внутренние органоподобные структуры, непохожие на что-либо, найденное у современных гребневиков. У одного из ископаемых видов, впервые зарегистрированного в 1996 году, был большой рот, по-видимому, окруженный складчатым краем, который мог быть мускулистым. [3] Данные из Китая, полученные годом позже, свидетельствуют о том, что такие гребневики были широко распространены в кембрии, но, возможно, сильно отличались от современных видов — например, гребневые ряды одной окаменелости были установлены на выступающих лопастях. [90] Самая молодая окаменелость вида за пределами группы короны — это вид Daihuoides из позднего девона, который принадлежит к базальной группе, которая, как предполагалось, вымерла более 140 миллионов лет назад. [91]
Эдиакарская эоандромеда предположительно может представлять собой гребневик. [4] У нее восьмикратная симметрия с восемью спиральными рукавами, напоминающими гребневидные ряды гребневика. Если это действительно гребневик, то это помещает группу близко к происхождению Bilateria. [92] Раннекембрийская сидячая вейцеобразная окаменелость Stromatoveris из китайского лагерштетта Чэнцзян , датируемая примерно 515 миллионами лет назад , очень похожа на Vendobionta предыдущего эдиакарского периода. Де-Ган Шу, Саймон Конвей Моррис и др. обнаружили на ее ветвях то, что они считали рядами ресничек, используемых для фильтрационного питания . Они предположили, что Stromatoveris был эволюционной «тетушкой» гребневиков, и что гребневики произошли от сидячих животных, чьи потомки стали пловцами и изменили реснички с механизма питания на двигательную систему. [93] Другие кембрийские окаменелости, которые подтверждают идею о том, что гребневики произошли от сидячих форм, — это Dinomischus , Daihua , Xianguangia и Siphusauctum, которые также жили на морском дне, имели органические скелеты и покрытые ресничками щупальца, окружавшие их рот, которые были обнаружены кладистическим анализом как члены стволовой группы гребневиков [94] [95]
Кембрийские окаменелости возрастом 520 миллионов лет, также из Чэнцзяна в Китае, показывают ныне полностью вымерший класс гребневиков, названный « Scleroctenophora », который имел сложный внутренний скелет с длинными шипами. [96] Скелет также поддерживал восемь мягкотелых лоскутов, которые могли использоваться для плавания и, возможно, питания. Одна из форм, Thaumactena , имела обтекаемое тело, напоминающее тело стреловидных червей , и могла быть ловким пловцом. [5]
Филогенетическая связь гребневиков с остальными Metazoa очень важна для нашего понимания ранней эволюции животных и происхождения многоклеточности. Она была предметом дебатов в течение многих лет. Гребневики были заявлены как сестринские линии для Bilateria , [ 97] [98] сестры для Cnidaria , [99] [100] [101] [102] сестры для Cnidaria , Placozoa и Bilateria , [103] [104] [105] и сестры для всех других животных. [9] [106]
Вальтер Гарстанг в своей книге «Личинки и другие зоологические стихи» ( Mülleria и гребневик ) даже высказал теорию о том, что гребневики произошли от неотенической личинки Mülleria поликлады .
Серия исследований, в которых рассматривалось наличие и отсутствие членов семейств генов и сигнальных путей (например, гомеобоксов , ядерных рецепторов , сигнального пути Wnt и натриевых каналов ), продемонстрировала доказательства, соответствующие последним двум сценариям, что гребневики являются либо сестринскими по отношению к Cnidaria , Placozoa и Bilateria , либо сестринскими по отношению ко всем другим типам животных. [107] [108] [109] [110] Несколько более поздних исследований, сравнивающих полностью секвенированные геномы гребневиков с другими секвенированными геномами животных, также подтвердили, что гребневики являются сестринской линией по отношению ко всем другим животным. [111] [28] [112] [113] Эта позиция предполагает, что типы нервных и мышечных клеток либо были утрачены в основных линиях животных (например, Porifera и Placozoa ), либо эволюционировали независимо в линии гребневиков. [111]
Другие исследователи утверждали, что размещение Ctenophora как сестры по отношению ко всем другим животным является статистической аномалией, вызванной высокой скоростью эволюции геномов гребневиков, и что Porifera (губки) являются самым ранним дивергентным таксоном животных. [105] [114] [115] [116] [117] У них также чрезвычайно высокие скорости митохондриальной эволюции, [118] и наименьшее известное содержание РНК/белка в геноме мтДНК у животных. [119] Таким образом, Ctenophora, по-видимому, являются базальной кладой диплобластов . В соответствии с последним пунктом, анализ очень большого выравнивания последовательностей в таксономической шкале метазоа (1719 белков, в общей сложности около 400 000 аминокислотных позиций) показал, что гребневики появляются как вторая по раннему разветвляющаяся линия животных, а губки являются сестринской группой для всех других многоклеточных животных. [8] Кроме того, исследование генов муцина , которые позволяют животным вырабатывать слизь, показывает, что у губок их никогда не было, в то время как все другие животные, включая гребневиков, по-видимому, имеют гены общего происхождения. [120] И было обнаружено, что, несмотря на все их различия, нейроны гребневиков имеют ту же основу, что и нейроны книдарий, после того, как открытия показали, что нейроны, экспрессирующие пептиды, вероятно, являются предками химических нейротрансмиттеров. [121]
Еще одно исследование решительно отвергает гипотезу о том, что губки являются сестринской группой для всех других существующих животных, и устанавливает размещение гребневиков как сестринской группы для всех других животных, а несогласие с последней упомянутой статьей объясняется методологическими проблемами в анализах в этой работе. [13] Ни гребневики, ни губки не обладают путями HIF , [122] их геном экспрессирует только один тип потенциалзависимого кальциевого канала в отличие от других животных, у которых есть три типа, [123] и они являются единственными известными типами животных, у которых отсутствуют какие-либо истинные hox-гены . [28] Несколько видов из других типов; личинка немертины пилидий, личинка вида форонид Phoronopsis harmeri и личинка желудевого червя Schizocardium californicum , также не зависят от hox-генов в своем личиночном развитии, но нуждаются в них во время метаморфоза, чтобы достичь своей взрослой формы. [124] [125] [126] Гены иннексина , которые кодируют белки, используемые для межклеточной коммуникации у животных, также, по-видимому, независимо эволюционировали у гребневиков. [127]
Поскольку все современные гребневики, за исключением бероидов, имеют личинки, похожие на цидиппидов, широко распространено мнение, что их последний общий предок также напоминал цидиппидов, имея яйцевидное тело и пару втягивающихся щупалец. Чисто морфологический анализ Ричарда Харбисона в 1985 году пришел к выводу, что цидиппиды не являются монофилетическими, другими словами, не содержат всех и только потомков одного общего предка, который сам был цидиппидом. Вместо этого он обнаружил, что различные семейства цидиппидов были больше похожи на членов других отрядов гребневиков , чем на других цидиппидов. Он также предположил, что последний общий предок современных гребневиков был либо цидиппидоподобным, либо бероидоподобным. [130] Молекулярный филогенный анализ 2001 года с использованием 26 видов, включая 4 недавно открытых, подтвердил, что цидиппиды не являются монофилетическими, и пришел к выводу, что последний общий предок современных гребневиков был цидиппидоподобным. Он также обнаружил, что генетические различия между этими видами были очень малы — настолько малы, что отношения между Lobata, Cestida и Thalassocalycida оставались неопределенными. Это говорит о том, что последний общий предок современных гребневиков был относительно недавним и, возможно, пережил мел-палеогеновое вымирание 65,5 миллионов лет назад, в то время как другие линии вымерли. Когда анализ был расширен, включив представителей других типов , он пришел к выводу, что книдарии, вероятно, более тесно связаны с билатериями, чем любая из групп с гребневиками, но этот диагноз неопределен. [128] Более позднее исследование 2017 года подтверждает парафилию Cydippida , но также обнаруживает, что Lobata парафилетична по отношению к Cestida . Однако, помимо этого, в дереве гребневиков все еще есть много пробелов, несмотря на их интенсивное изучение. Несколько семейств и отрядов не имеют видов с полными геномами, и, таким образом, их размещение остается неопределенным.
Исследование гребневиков выявило тип мессенджера, который, вероятно, функционировал в нервной системе предков.— цитирует журнальную статью:
Мозг гребневика предполагает, что если бы эволюция началась снова, интеллект бы снова появился, потому что природа повторяется.— частично обзор исследований до Мороза и др. (2014)
{{cite report}}: CS1 maint: неподходящий URL ( ссылка )— включает в себя фотографии